
Phage Encounters Recorded in CRISPR Arrays in the Genus Oenococcus
 , and
, and
Abstract
:1. Introduction
2. Material and Methods
2.1. Bacteria and Culture Conditions
2.2. Phage Lysates Used in the Study
2.3. PCR Identification of of Lysogenic Strains
2.4. Mitomycin C Induction of Lysogenic Strains
2.5. Host Spectrum Determination
2.6. Genome Sequencing
2.7. Bioinformatic Analysis
2.8. Typing System for Rapid Assesment of Diversity amongst Uncharactized Oenophages
2.9. CRISPR-Cas Systems and Anti-CRISPR Identification
3. Results and Discussion
3.1. Distribution and Architecture of CRISPR Loci in the Genus
3.2. Selection of Representative MGE of O. oeni for Interspecies CRISPR Targeting
3.3. Intra- and Interspecies CRISPR Targeting of Known Prophages and Plasmids of Oenococcus oeni
3.4. Autoimmunity in O. sicerae
3.5. Protospacer-Containing Prophages and Genes in O. oeni
3.6. Search for Cross-Infections in the Oenococcus Genus
4. Conclusive Remarks
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- International Organisation of Vine and Wine [OIV]. Annual Report; International Organisation of Vine and Wine: Paris, France, 2021. [Google Scholar]
- Vitulo, N.; Lemos, W.J.F., Jr.; Calgaro, M.; Confalone, M.; Felis, G.E.; Zapparoli, G.; Nardi, T. Bark and grape microbiome of Vitis vinifera: Influence of geographic patterns and agronomic management on bacterial diversity. Front. Microbiol. 2019, 9, 3203. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fournier, P.; Pellan, L.; Barroso-Bergad, D.; Bohanc, D.A.; Candresse, T.; Delmotte, F.; Dufour, M.-C.; Lauvergeat, V.; Le Marrec, C.; Marais, A.; et al. The functional microbiome of grapevine throughout plant evolutionary history and lifetime. Adv. Ecol. Res. 2022; in press. [Google Scholar]
- Lorentzen, M.P.G.; Lucas, P.M. Distribution of Oenococcus oeni populations in natural habitats. Appl. Microbiol. Biotechnol. 2019, 103, 2937–2945. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sánchez, A.; Coton, M.; Coton, E.; Herrero, M.; García, L.A.; Díaz, M. Prevalent lactic acid bacteria in cider cellars and efficiency of Oenococcus oeni strains. Food Microbiol. 2012, 32, 32–37. [Google Scholar] [CrossRef] [PubMed]
- Lorentzen, M.P.; Campbell-Sills, H.; Jorgensen, T.S.; Nielsen, T.; Coton, M.; Coton, E.; Hansen, L.; Lucas, P.M. Expanding the biodiversity of Oenococcus oeni through comparative genomics of apple cider and kombucha strains. BMC Genom. 2019, 20, 330. [Google Scholar] [CrossRef] [PubMed]
- Suhre, T.; Mann, M.B.; Kothe, C.I.; Rocha, A.L.G.; Celso, P.G.; Varela, A.P.M.; Frazzon, A.P.G.; Frazzon, J. Microbial community and physicochemical characterization of kombuchas produced and marketed in Brazil. J. Food Sci. Nutr. Res. 2021, 4, 302–316. [Google Scholar]
- Leech, J.; Cabrera-Rubio, R.; Walsh, A.M.; Macori, G.; Walsh, C.J.; Barton, W.; Finnegan, L.; Crispie, F.; O’Sullivan, O.; Claesson, M.J.; et al. Fermented-Food Metagenomics Reveals Substrate-Associated Differences in Taxonomy and Health-Associated and Antibiotic Resistance Determinants. mSystems 2020, 5, e00522-20. [Google Scholar] [CrossRef]
- Varakumar, S.; Naresh, K.; Variyar, P.S.; Sharma, A.; Reddy, O.V.S. Role of Malolactic Fermentation on the Quality of Mango (Mangifera indica L.) Wine. Food Biotechnol. 2013, 27, 119–136. [Google Scholar] [CrossRef]
- Pothakos, V.; De Vuyst, L.; Zhang, S.J.; De Bruyn, F.; Verce, M.; Torres, J.; Callanan, M.; Moccand, C.; Weckx, S. Temporal shotgun metagenomics of an Ecuadorian coffee fermentation process highlights the predominance of lactic acid bacteria. Curr. Res. Biotechnol. 2020, 2, 1–15. [Google Scholar] [CrossRef]
- Mattila, H.R.; Rios, D.; Walker-Sperling, V.E.; Roeselers, G.; Newton, I.L.G. Characterization of the Active Microbiotas Associated with Honey Bees Reveals Healthier and Broader Communities when Colonies are Genetically Diverse. PLoS ONE 2012, 7, e32962. [Google Scholar] [CrossRef]
- Dicks, L.; Endo, A. Are fructophilic lactic acid bacteria (FLAB) beneficial to humans? Benef. Microbes 2022, 13, 3–11. [Google Scholar] [CrossRef] [PubMed]
- Badotti, F.; Moreira, A.P.; Tonon, L.A.; de Lucena, B.T.; Gomes Fde, C.; Kruger, R.; Thompson, C.C.; de Morais, M.A., Jr.; Rosa, C.A.; Thompson, F.L. Oenococcus alcoholitolerans sp. nov., a lactic acid bacteria isolated from cachaça and ethanol fermentation processes. Antonie Van Leeuwenhoek 2004, 106, 1259–1267. [Google Scholar] [CrossRef] [PubMed]
- Endo, A.; Okada, S. Oenococcus kitaharae sp. nov., a non-acidophilic and non-malolactic-fermenting oenococcus isolated from a composting distilled shochu residue. Int. J. Syst. Evol. Microbiol. 2006, 56, 2345–2348. [Google Scholar] [CrossRef] [Green Version]
- Cousin, F.J.; Le Guellec, R.; Chagnot, C.; Goux, D.; Dalmasso, M.; Laplace, J.-M.; Cretenet, M. Oenococcus sicerae sp. nov., isolated from French cider. Syst. Appl. Microbiol. 2019, 42, 302–308. [Google Scholar] [CrossRef] [PubMed]
- Verce, M.; De Vuyst, L.; Weckx, S. The metagenome-assembled genome of Candidatus Oenococcus aquikefiri from water kefir represents the species Oenococcus sicerae. Food Microbiol. 2020, 88, 103402. [Google Scholar] [CrossRef] [PubMed]
- Koskella, B.; Brockhurst, M.A. Bacteria–phage coevolution as a driver of ecological and evolutionary processes in microbial communities. FEMS Microbiol. Rev. 2014, 38, 916–931. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jaomanjaka, F.; Ballestra, P.; Dols-Lafargue, M.; Le Marrec, C. Expanding the diversity of oenococcal bacteriophages: Insights into a novel group based on the integrase sequence. Int. J. Food Microbiol. 2013, 166, 331–340. [Google Scholar] [CrossRef]
- Claisse, O.; Chaïb, A.; Jaomanjaka, F.; Philippe, C.; Barchi, Y.; Lucas, P.M.; Le Marrec, C. Distribution of Prophages in the Oenococcus oeni Species. Microorganisms 2021, 9, 856. [Google Scholar] [CrossRef]
- Jaomanjaka, F.; Philippe, C.; Claisse, O.; Le Marrec, C. Complete Genome Sequence of Lytic Oenococcus oeni Bacteriophage OE33PA. Microbiol. Resour. Announc. 2018, 7, e00818-18. [Google Scholar] [CrossRef] [Green Version]
- Philippe, C.; Jaomanjaka, F.; Claisse, O.; Laforgue, R.; Maupeu, J.; Petrel, M.; Le Marrec, C. A survey of oenophages during wine making reveals a novel group with unusual genomic characteristics. Int. J. Food Microbiol. 2017, 257, 138–147. [Google Scholar] [CrossRef]
- Philippe, C.; Chaïb, A.; Jaomanjaka, F.; Claisse, O.; Lucas, P.M.; Samot, O.J.; Cambillau, C.; Le Marrec, C. Characterization of the First Virulent Phage Infecting Oenococcus oeni, the Queen of the Cellars. Front. Microbiol. 2021, 11, 103389. [Google Scholar] [CrossRef] [PubMed]
- Walker, P.J.; Siddell, S.G.; Lefkowitz, E.J.; Mushegian, A.R.; Adriaenssens, E.M.; Alfenas-Zerbini, P.; Davison, A.J.; Dempsey, D.M.; Dutilh, B.E.; García, M.L.; et al. Changes to virus taxonomy and to the International Code of Virus Classification and Nomenclature ratified by the International Committee on Taxonomy of Viruses. Arch. Virol. 2021, 166, 2633–2648. [Google Scholar] [CrossRef] [PubMed]
- Chaïb, A.; Philippe, C.; Jaomanjaka, F.; Barchi, Y.; Oviedo-Hernandez, F.; Claisse, O.; Le Marrec, C. Phage-host interactions as a driver of population dynamics during wine fermentation: Betting on underdogs. Int. J. Food Microbiol. 2022, 383, 109936. [Google Scholar] [CrossRef] [PubMed]
- Barrangou, R.; Fremaux, C.; Deveau, H.; Richards, M.; Boyaval, P.; Moineau, S.; Romero, D.A.; Horvath, P. CRISPR provides acquired resistance against viruses in prokaryotes. Science 2007, 315, 1709–1712. [Google Scholar] [CrossRef]
- Virdis, C.; Sumby, K.; Bartowsky, E.; Jiranek, V. Lactic Acid Bacteria in Wine: Technological Advances and Evaluation of Their Functional Role. Front. Microbiol. 2021, 11, 612118. [Google Scholar] [CrossRef]
- Philippe, C.; Moineau, S. The endless battle between phages and CRISPR–Cas systems in Streptococcus thermophilus. Biochem. Cell Biol. 2021, 99, 397–402. [Google Scholar] [CrossRef]
- Westra, E.R.; Van Houte, S.; Gandon, S.; Whitaker, R. The ecology and evolution of microbial CRISPR-Cas adaptive immune systems. Philos. Trans. R. Soc. B Biol. Sci. 2019, 374, 20190101. [Google Scholar] [CrossRef] [Green Version]
- Marcobal, A.M.; Sela, D.A.; Wolf, Y.I.; Makarova, K.S.; Mills, D.A. Role of Hypermutability in the Evolution of the Genus Oenococcus. J. Bacteriol. 2008, 190, 564–570. [Google Scholar] [CrossRef] [Green Version]
- Sorek, R.; Kunin, V.; Hugenholtz, P. CRISPR—A widespread system that provides acquired resistance against phages in bacteria and archaea. Nat. Rev. Microbiol. 2008, 6, 181–186. [Google Scholar] [CrossRef]
- Chevallereau, A.; Meaden, S.; van Houte, S.; Westra, E.R.; Rollie, C. The effect of bacterial mutation rate on the evolution of CRISPR-Cas adaptive immunity. Philos. Trans. R. Soc. B 2019, 374, 20180094. [Google Scholar] [CrossRef] [Green Version]
- Borneman, A.R.; McCarthy, J.M.; Chambers, P.J.; Bartowsky, E.J. Functional Divergence in the Genus Oenococcus as Predicted by Genome Sequencing of the Newly-Described Species, Oenococcus kitaharae. PLoS ONE 2012, 7, e29626. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dion, M.B.; Plante, P.-L.; Zufferey, E.; Shah, S.A.; Corbeil, J.; Moineau, S. Streamlining CRISPR spacer-based bacterial host predictions to decipher the viral dark matter. Nucleic Acids Res. 2021, 49, 3127–3138. [Google Scholar] [CrossRef] [PubMed]
- Crawley, A.B.; Henriksen, E.D.; Stout, E.; Brandt, K.; Barrangou, R. Characterizing the activity of abundant, diverse and active CRISPR-Cas systems in lactobacilli. Sci. Rep. 2018, 8, 11544. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barchi, Y.; Chaïb, A.; Philippe, C.; Claisse, O.; Le Marrec, C. Draft genome sequence of Oenococcus kitaharae CRBO2176, isolated from a home-made water kefir. Genome Announc. 2022; submitted. [Google Scholar]
- Chaïb, A.; Claisse, O.; Delbarre, E.; Bosviel, J.; Le Marrec, C. Assessment of the lysogenic status in the lactic acid bacterium Oenococcus oeni during the spontaneous malolactic fermentation of red wines. Food Microbiol. 2022, 103, 103947. [Google Scholar] [CrossRef]
- Glonti, T.; Pirnay, J.-P. In Vitro Techniques and Measurements of Phage Characteristics That Are Important for Phage Therapy Success. Viruses 2022, 14, 1490. [Google Scholar] [CrossRef]
- Chaïb, A.; Decossas, M.; Philippe, C.; Claisse, O.; Lambert, O.; Marrec, C.L. Isolation and CryoTEM of Phages Infecting Bacterial Wine Spoilers. BIO-Protocol 2020, 10, e3801. [Google Scholar] [CrossRef]
- Bankevich, A.; Nurk, S.; Antipov, D.; Gurevich, A.A.; Dvorkin, M.; Kulikov, A.S.; Lesin, V.M.; Nikolenko, S.I.; Pham, S.; Prjibelski, A.D.; et al. SPAdes: A new genome assembly algorithm and its applications tosingle-cell sequencing. J. Comput. Mol. Cell Biol. 2012, 19, 455–477. [Google Scholar] [CrossRef] [Green Version]
- McNair, K.; Aziz, R.K.; Pusch, G.D.; Overbeek, R.; Dutilh, B.E.; Edwards, R. Phage Genome Annotation Using the RAST Pipeline. Methods Mol. Biol. 2018, 1681, 231–238. [Google Scholar]
- Söding, J.; Biegert, A.; Lupas, A.N. The HHpred interactive server for protein homology detection and structure prediction. Nucleic Acids Res. 2005, 33, W244–W248. [Google Scholar] [CrossRef] [Green Version]
- Meier-Kolthoff, J.P.; Auch, A.F.; Klenk, H.-P.; Göker, M. Genome sequence-based species delimitation with confidence intervals and improved distance functions. BMC Bioinform. 2013, 14, 60. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meier-Kolthoff, J.P.; Göker, M. VICTOR: Genome-based phylogeny and classification of prokaryotic viruses. Bioinformatics 2017, 33, 3396–3404. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Untergasser, A.; Cutcutache, I.; Koressaar, T.; Ye, J.; Faircloth, B.C.; Remm, M.; Rozen, S.G. Primer3—New capabilities and interfaces. Nucleic Acids Res. 2012, 40, e115. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Couvin, D.; Bernheim, A.; Toffano-Nioche, C.; Touchon, M.; Michalik, J.; Néron, B.; Rocha, E.; Vergnaud, G.; Gautheret, D.; Pourcel, C. CRISPRCasFinder, an update of CRISRFinder, includes a portable version, enhanced performance and integrates search for Cas proteins. Nucleic Acids Res. 2018, 46, W246–W251. [Google Scholar] [CrossRef]
- Guindon, S.; Dufayard, J.-F.; Lefort, V.; Anisimova, M.; Hordijk, W.; Gascuel, O. New Algorithms and Methods to Estimate Maximum-Likelihood Phylogenies: Assessing the Performance of PhyML 3.0. Syst. Biol. 2010, 59, 307–321. [Google Scholar] [CrossRef] [Green Version]
- Huang, L.; Yang, B.; Yi, H.; Asif, A.; Wang, J.; Lithgow, T.; Zhang, H.; Minhas, F.A.; Yin, Y. AcrDB: A database of anti-CRISPR operons in prokaryotes and viruses. Nucleic Acids Res. 2020, 49, D622–D629. [Google Scholar] [CrossRef]
- Hidalgo-Cantabrana, C.; Goh, Y.J.; Pan, M.; Sanozky-Dawes, R.; Barrangou, R. Genome editing using the endogenous type I CRISPR-Cas system in Lactobacillus crispatus. Proc. Natl. Acad. Sci. USA 2019, 116, 15774–15783. [Google Scholar] [CrossRef] [Green Version]
- Börner, R.A.; Kandasamy, V.; Axelsen, A.M.; Nielsen, A.T.; Bosma, E.F. Genome editing of lactic acid bacteria: Opportunities for food, feed, pharma and biotech. FEMS Microbiol. Lett. 2019, 366, fny291. [Google Scholar] [CrossRef]
- Hidalgo-Cantabrana, C.; Crawley, A.B.; Sanchez, B.; Barrangou, R. Characterization and Exploitation of CRISPR Loci in Bifidobacterium longum. Front. Microbiol. 2017, 8, 1851. [Google Scholar] [CrossRef] [Green Version]
- Lee, S.H.; Jung, J.Y.; Lee, S.H.; Jeon, C.O. Complete genome sequence of Leuconostoc kimchii strain C2, isolated from Kimchi. J. Bacteriol. 2011, 193, 5548. [Google Scholar] [CrossRef] [Green Version]
- McDonald, N.D.; Regmi, A.; Morreale, D.P.; Borowski, J.D.; Boyd, E.F. CRISPR-Cas systems are present predominantly on mobile genetic elements in Vibrio species. BMC Genom. 2019, 20, 105. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Garrett, S.C. Pruning and Tending Immune Memories: Spacer Dynamics in the CRISPR Array. Front. Microbiol. 2021, 12, 664299. [Google Scholar] [CrossRef] [PubMed]
- Martynov, A.; Severinov, K.; Ispolatov, I. Optimal number of spacers in CRISPR arrays. PLoS Comput. Biol. 2017, 13, e1005891. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Misery, B.; Legendre, P.; Rue, O.; Bouchart, V.; Guichard, H.; Laplace, J.M.; Cretenet, M. Diversity and dynamics of bacterial and fungal communities in cider for distillation. Int. J. Food Microbiol. 2021, 339, 108987. [Google Scholar] [CrossRef] [PubMed]
- Ledormand, P.; Desmasures, N.; Bernay, B.; Goux, D.; Monnet, C.; Dalmasso, M. Molecular approaches to uncover phage-lactic acid bacteria interactions in a model community simulating fermented beverages. Food Microbiol. 2022, 107, 104069. [Google Scholar] [CrossRef]
- Ledormand, P.; Desmasures, N.; Midoux, C.; Rué, O.; Dalmasso, M. Investigation of the Phageome and Prophages in French Cider, a Fermented Beverage. Microorganisms 2022, 10, 1203. [Google Scholar] [CrossRef]
- Favier, M.; Bilhère, E.; Lonvaud, A.; Moine, V.; Lucas, P.M. Identification of pOENI-1 and Related Plasmids in Oenococcus oeni Strains Performing the Malolactic Fermentation in Wine. PLoS ONE 2012, 7, e49082. [Google Scholar] [CrossRef] [Green Version]
- Mesas, J.M.; Rodríguez, M.C.; Alegre, M.T. Nucleotide Sequence Analysis of pRS2 and pRS3, Two Small Cryptic Plasmids from Oenococcus oeni. Plasmid 2001, 46, 149–151. [Google Scholar] [CrossRef]
- Costello, P.J.; Kolouchova, R.; Jordans, C.; McCarthy, J.; Schmidt, S.A. Analysis of Transcriptomic Response to SO2 by Oenococcus oeni Growing in Continuous Culture. Microbiol. Spectr. 2021, 9, e0115421. [Google Scholar]
- Tisza, M.J.; Buck, C.B. A catalog of tens of thousands of viruses from human metagenomes reveals hidden associations with chronic diseases. Proc. Natl. Acad. Sci. USA 2021, 118, e2023202118. [Google Scholar] [CrossRef]
- Nobrega, F.L.; Walinga, H.; Dutilh, B.E.; Brouns, S. Prophages are associated with extensive CRISPR–Cas auto-immunity. Nucleic Acids Res. 2020, 48, 12074–12084. [Google Scholar] [CrossRef] [PubMed]
- Watson, B.N.J.; Easingwood, R.A.; Tong, B.; Wolf, M.; Salmond, G.P.C.; Staals, R.H.J.; Bostina, M.; Fineran, P.C. Different genetic and morphological outcomes for phages targeted by single or multiple CRISPR-Cas spacers. Philos. Trans. R. Soc. B Biol. Sci. 2019, 374, 20180090. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nicholson, T.J.; Jackson, S.A.; Croft, B.I.; Staals, R.H.J.; Fineran, P.C.; Brown, C.M. Bioinformatic evidenceof widespread priming in Type I and II CRISPR-Cassystems. RNA Biol. 2018, 18, 1–11. [Google Scholar]
- Bondy-Denomy, J.; Pawluk, A.; Maxwell, K.L.; Davidson, A.R. Bacteriophage genes that inactivate the CRISPR/Cas bacterial immune system. Nature 2013, 493, 429–432. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yin, Y.; Yang, B.; Entwistle, S. Bioinformatics Identification of Anti-CRISPR Loci by Using Homology, Guilt-by-Association, and CRISPR Self-Targeting Spacer Approaches. mSystems 2019, 4, e00455-19. [Google Scholar] [CrossRef] [Green Version]
- Göller, P.C.; Elsener, T.; Lorgé, D.; Radulovic, N.; Bernardi, V.; Naumann, A.; Amri, N.; Khatchatourova, E.; Hernandes Coutinho, F.; Loessner, M.J.; et al. Multi-species host range of staphylococcal phages isolated from wastewater. Nat. Commun. 2021, 12, 6965. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
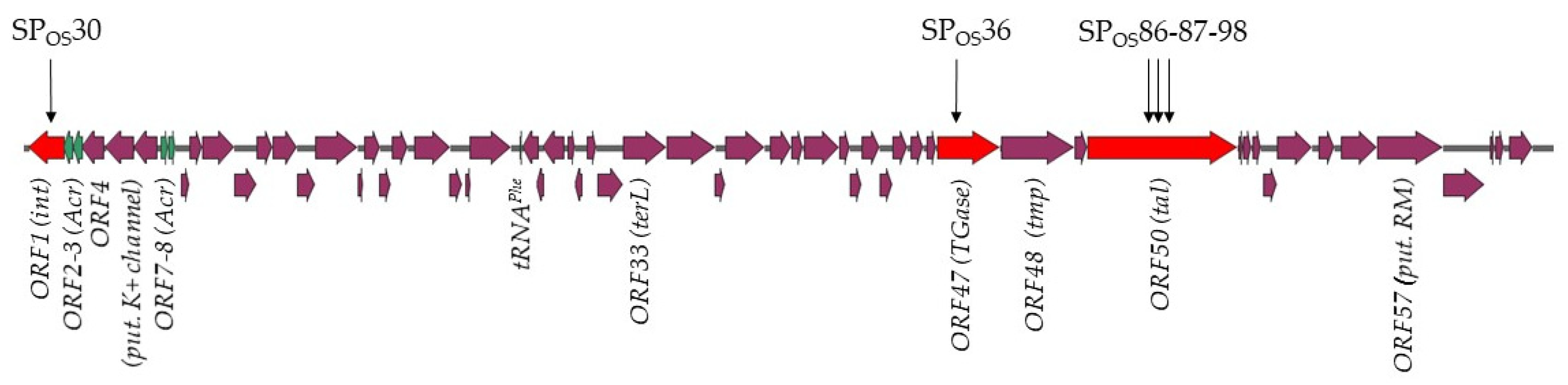{kind=link}
{kind=link}
| CRISPR Loci | DR | Spacers | |||||
|---|---|---|---|---|---|---|---|
| Species | Type | Size (bp) | Sequence | Size (bp) | Total | Unique | With Matches in O. oeni |
| O. kitaharae DSM17330T | II-A | 36 | GCTTCAGATGTGTGTCAGATCAATGAGGTAGAACCC | 30 | 57 | 56 | 3 |
| O. sicerae UCMA15228T | II-A | 36 | GGGTGTCACCCCATTAATCTGACATACAACTGAAGC | 29–31 | 23 | 21 | 2 |
| I-E | 29 | AGGATCACCCCCGCTTGTGCGGGGAATAC | 32–33 | 102 | 91 | 5 | |
| O. alcoholitolerans UFRJ-M7.2.18T | II-A | 35 | GCTTCAGATGTGTGTCAGATCAATGAGGTAGAACC | 30–31 | 15 | 15 | 0 |
| Type | Sequences of Spacers (SP), Protospacers and Relevant Phages | Phage/Plasmid Functions Targeted by CRISPR Systems | |
|---|---|---|---|
| II Ok | SPOKII2 IOEB1491ProA |  | Dit (phage baseplate protein, WP_032824876) |
| SPOKII9 IOEB1491ProA |  | ||
| SPOKII15 pOeni1/2; pAWRIB429 |  | Intergenic sequence | |
| II Os | SPOSII1 Vinitor27 |  | Rep5 (Replication inititation protein QNO11543) |
| SPOSII21 S161ProA |  | Tal (peptidase, KGH92654) | |
| I Os | SPOSI5 IOEB0501ProA |  | Head closure protein (WP_002820745) |
| SPOSI7 S161ProA |  | TMP2 (KGH92652) | |
| SPOSI43 IOEB0205ProA |  | HP (WP_002825766) | |
| SPOSI46 IOEB0501ProA |  | Portal protein (WP_032822098) | |
| SPOSI78 S161 ProA |  | Bifunctional DNA primase/polymerase (WP_002824107) | |
| SPOSI89 S161ProA |  | ||
| STSOSI30 Resident prophage | AATGTGCTTGATTCAAGCTATGGCAATGACTC | Transglycosylase soluble domain- containing protein | |
| STSOSI36 Resident prophage | CCCACCAGCTGTCTGGCATTGAGATAGTTCGT | Tyrosine-type recombinase/integrase (QAS70227) | |
| STSOSI86 Resident prophage | ATTTATTAGGACGAGGGACTCCGATGGTGAAC | Tal (WP_243148548) | |
| STSOSI87 Resident prophage | ACCATCGTTTGATAAGTCGAGCTCTCAACTTT | ||
| STSOSI9 Resident prophage | CAAGAAGTTCCACTAATCACTGTCGTCGCAGC | ||
| Phages | Bacteria, Species and Origin | ||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| O. oeni | O. k | O. s | O. a | ||||||||||||
| W | C | Ko | Kef | S | C | E | |||||||||
| Host species and habitat | Name (type of lysate) | Clus-ter | IOEB S277 | CRBO 1381 | CRBO 1384 | BL4 | CRBO 2176 | NRIC 0649 | UCMA 15228 | 736 | |||||
| O. oeni | W | OE33PA (P) | I.1 | ++ | ++ | + | ++ | − | − | − | − | ||||
| Krappator27 (P) | II.1 | ++ | ++ | - | ++ | − | − | − | − | ||||||
| Vinitor 162 (P) | II.2 | ++ | − * | + | − * | − | − | − | − | ||||||
| C | CRBO1384ProE (MC) | I.2 | ++ ** | − | − | − | − | − | − | ||||||
| Ko | BL4 ProF (MC) | I.2 | − | − | − | − | − | − | − | ||||||
| O. c | C | UCMA15228 (MC) | nd | − | − | − | − | − | − | − | |||||
| O. a | E | O. alcoholitolerans JP736 (MC) | nd | − | − | − | − | − | − | − | |||||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Barchi, Y.; Philippe, C.; Chaïb, A.; Oviedo-Hernandez, F.; Claisse, O.; Le Marrec, C. Phage Encounters Recorded in CRISPR Arrays in the Genus Oenococcus. Viruses 2023, 15, 15. https://doi.org/10.3390/v15010015
Barchi Y, Philippe C, Chaïb A, Oviedo-Hernandez F, Claisse O, Le Marrec C. Phage Encounters Recorded in CRISPR Arrays in the Genus Oenococcus. Viruses. 2023; 15(1):15. https://doi.org/10.3390/v15010015
Chicago/Turabian StyleBarchi, Yasma, Cécile Philippe, Amel Chaïb, Florencia Oviedo-Hernandez, Olivier Claisse, and Claire Le Marrec. 2023. "Phage Encounters Recorded in CRISPR Arrays in the Genus Oenococcus" Viruses 15, no. 1: 15. https://doi.org/10.3390/v15010015