
Early or Simultaneous Infection with Infectious Pancreatic Necrosis Virus Inhibits Infectious Hematopoietic Necrosis Virus Replication and Induces a Stronger Antiviral Response during Co-infection in Rainbow Trout (Oncorhynchus mykiss)

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cell and Virus Culture
2.2. Fish Samples and Virus Infection Design
2.2.1. Infectious Dose Selection
2.2.2. Single-Infection
2.2.3. Co-Infection
2.3. The Survival Rate of the Challenged Fish
2.4. Determination of Viral Target Gene Expression
2.5. Expression of Immune-Related Genes Determined by qRT-PCR
2.6. Statistical Analysis
3. Results
3.1. Effect of Co-Infection on Survival Rate
3.2. Distribution and Expression of Viral Target Genes
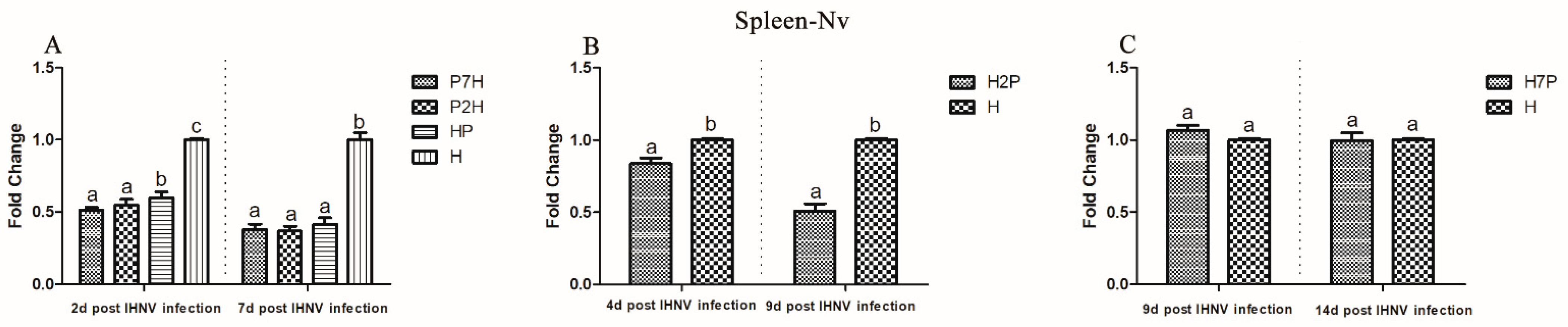
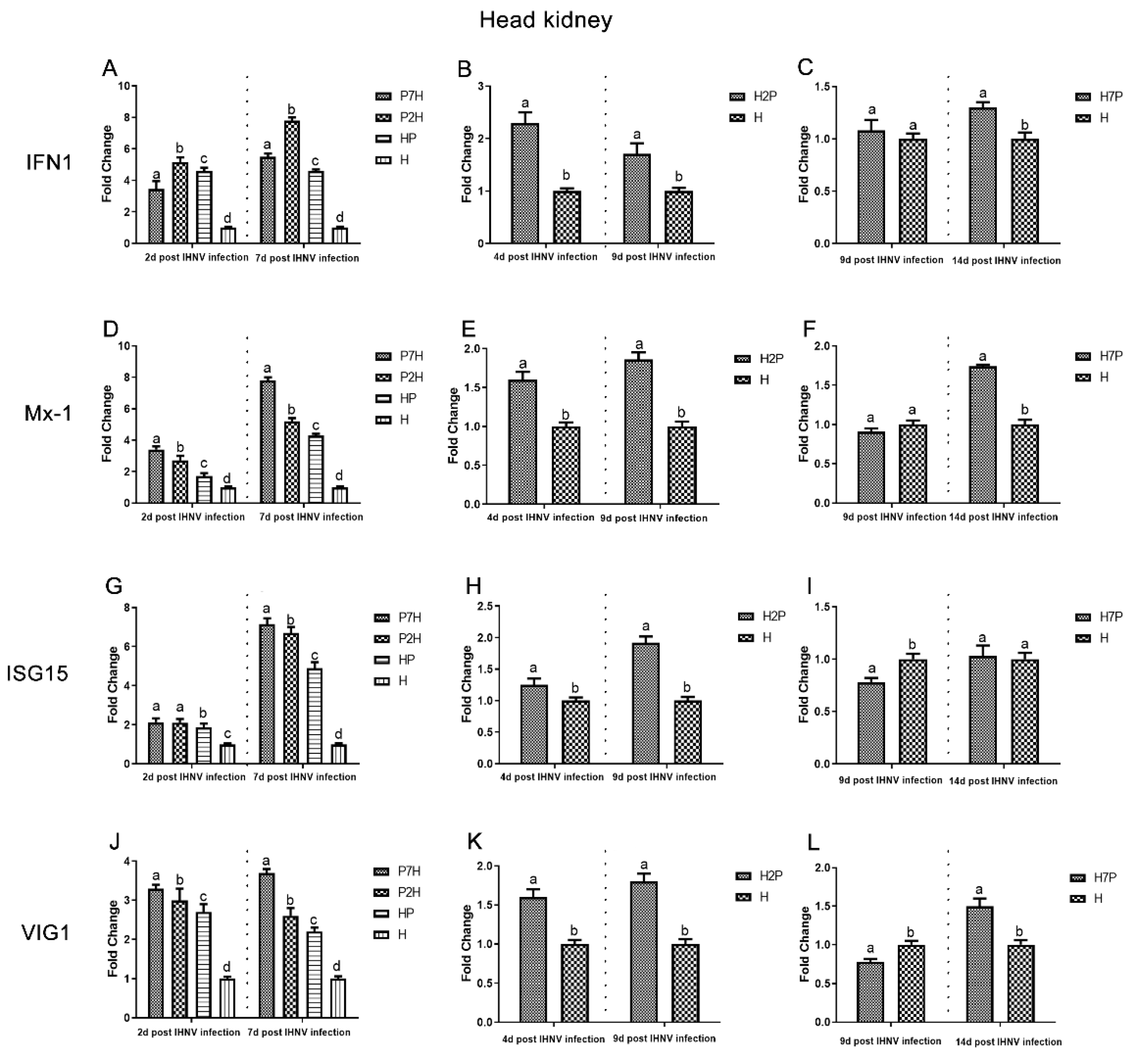
3.3. Expression Level of Immune-Related Factors
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Informed Consent Statement
Conflicts of Interest
References
- Bakaletz, L.O. Developing animal models for polymicrobial diseases. Nat. Rev. Microbiol. 2004, 2, 552–568. [Google Scholar] [CrossRef] [PubMed]
- Fox, F.E. Concomitant infections, parasites and immune responses. Parasitology 2001, 122, S23–S38. [Google Scholar]
- Emmenegger, E.J.; Biacchesi, S.; Mérour, E.; Glenn, J.A.; Palmer, A.D.; Brémont, M.; Kurath, G. Virulence of a chimeric recombinant infectious haematopoietic necrosis virus expressing the spring viraemia of carp virus glycoprotein in salmonid and cyprinid fish. J. Fish Dis. 2018, 41, 67–78. [Google Scholar] [CrossRef] [PubMed]
- Zhao, J.Z.; Xu, L.M.; Ren, G.M.; Dong, Y.; Cao, Y.S.; Liu, H.B.; Yin, J.S.; Lu, T.Y. Complete Genome Sequence and Phylogenetic Analysis of Sn1203 Strain of Infectious Hematopoietic Necrosis Virus. Chin. J. Fish. 2020, 33, 1–9. (In Chinese) [Google Scholar]
- Dixon, P.; Paley, R.; Alegria-Moran, R.; Oidtmann, B. Epidemiological characteristics of infectious hematopoietic necrosis virus (IHNV): A review. Vet. Res. 2016, 47, 63. [Google Scholar] [CrossRef]
- Julin, K.; Sommer, A.; Johansen, L.; Jorgensen, J.B. Persistent infections with infectious pancreatic necrosis virus (IPNV) of different virulence in Atlantic salmon, Salmo salar L. J. Fish Dis. 2015, 38, 1005–1019. [Google Scholar] [CrossRef]
- Dopazo, C.P. The Infectious Pancreatic Necrosis Virus (IPNV) and its Virulence Determinants: What is Known and What Should be Known. Pathogens 2020, 9, 94. [Google Scholar] [CrossRef]
- Rodríguez Saint-Jean, S.; Alonso, M.; Pérez-Prieto, S.I. Comparison of two birnavirus-rhabdovirus coinfections in fish cell lines. Dis. Aquat. Organ. 2005, 67, 183–190. [Google Scholar] [CrossRef]
- Guez, S.R.; Vilas, M.P.; Alonso, M.; Rez, S.P. Study of a viral-dual infection in rainbow trout (Oncorhynchus mykiss) by seroneutralization, western blot and polymerase chain reaction assays. Microbiologia 1995, 11, 461–470. [Google Scholar]
- Alonso, M.; Rodríguez Saint-Jean, S.; Pérez-Prieto, S.I. Virulence of infectious hematopoietic necrosis virus and Infectious pancreatic necrosis virus coinfection in rainbow trout (Oncorhynchus mykiss) and nucleotide sequence analysis of the IHNV glycoprotein gene. Arch. Virol. 2003, 148, 1507–1521. [Google Scholar] [CrossRef]
- Byrne, N.; Castric, J.; Lamour, F.; Cabon, J.; Quentel, C. Study of the viral interference between infectious pancreatic necrosis virus (IPNV) and infectious haematopoietic necrosis virus (IHNV) in rainbow trout (Oncorhynchus mykiss). Fish Shellfish Immun. 2008, 24, 489–497. [Google Scholar] [CrossRef] [PubMed]
- Xu, L.M.; Zhao, J.Z.; Ren, G.M.; Dong, Y.; Lin, J.N.; Cao, Y.S.; Yin, J.S.; Liu, H.B.; Lu, T.Y.; Zhang, Q.Y. Co-infection of infectious hematopoietic necrosis virus (IHNV) and infectious pancreatic necrosis virus (IPNV) caused high mortality in farmed rainbow trout (Oncorhynchus mykiss) in China. Aquaculture 2019, 512, 734286. [Google Scholar] [CrossRef]
- Kim, H.J.; Oseko, N.; Nishizawa, T.; Yoshimizu, M. Protection of rainbow trout from infectious hematopoietic necrosis (IHN) by injection of infectious pancreatic necrosis virus (IPNV) or poly(I:C). Dis. Aquat. Organ. 2009, 83, 105–113. [Google Scholar] [CrossRef] [PubMed]
- Xu, L.M.; Liu, M.; Zhao, J.Z.; Ren, G.M.; Dong, Y.; Shao, Y.Z.; Lu, T.Y.; Zhang, Q.Y. Infectious pancreatic necrosis virus inhibits infectious hematopoietic necrosis virus at the early stage of infection in a time dependent manner during Co-infection in Chinook salmon embryo cell lines. Fish Shellfish Immun. 2020, 102, 361–367. [Google Scholar] [CrossRef] [PubMed]
- Duan, K.Y.; Zhao, J.Z.; Ren, G.M.; Shao, Y.Z.; Lu, T.Y.; Xu, L.P.; Tang, X.; Zhao, W.W.; Xu, L.M. Molecular Evolution of Infectious Pancreatic Necrosis Virus in China. Viruses 2021, 13, 488. [Google Scholar] [CrossRef]
- Xu, L.M.; Zhao, J.Z.; Liu, M.; Ren, G.M.; Feng, J.; Yin, J.S.; Ji, F.; Liu, H.B.; Lu, T.Y. Bivalent DNA vaccine induces significant immune responses against infectious hematopoietic necrosis virus and infectious pancreatic necrosis virus in rainbow trout. Sci. Rep. 2017, 7, 5700. [Google Scholar] [CrossRef] [PubMed]
- Tapia, D.; Kuznar, J.; Farlora, R.; Yáñez, J.M. Differential Transcriptomic Response of Rainbow Trout to Infection with Two Strains of IPNV. Viruses 2021, 14, 21. [Google Scholar] [CrossRef]
- Soltani, M.; Ahmadivand, S.; Behdani, M.; Hassanzadeh, R.; Rahmati-Holasoo, H.; Taheri-Mirghaed, A. Transcription of adaptive-immune genes upon challenge with infectious pancreatic necrosis virus (IPNV) in DNA vaccinated rainbow trout. Int. J. Aquat. Biol. 2016, 4, 353–359. [Google Scholar]
- Zhao, J.Z.; Liu, M.; Xu, L.M.; Zhang, Z.Y.; Cao, Y.S.; Shao, Y.Z.; Yin, J.S.; Liu, H.B.; Lu, T.Y. A chimeric recombinant infectious hematopoietic necrosis virus induces protective immune responses against infectious hematopoietic necrosis and infectious pancreatic necrosis in rainbow trout. Mol. Immunol. 2019, 116, 180–190. [Google Scholar] [CrossRef]
- Chinchar, V.G.; Logue, O.; Antao, A.; Chinchar, G.D. Channel catfish reovirus (CRV) inhibits replication of channel catfish herpesvirus (CCV) by two distinct mechanisms: Viral interference and induction of an anti-viral factor. Dis. Aquat. Organ. 1998, 33, 77–85. [Google Scholar] [CrossRef]
- Brudeseth, B.E.; Castric, J.; Evensen, O. Studies on pathogenesis following single and double infection with viral hemorrhagic septicemia virus and infectious hematopoietic necrosis virus in rainbow trout (Oncorhynchus mykiss). Vet. Pathol. 2002, 39, 180–189. [Google Scholar] [CrossRef] [PubMed]
- López-Vázquez, C.; Alonso, M.C.; Dopazo, C.P.; Bandín, I. In vivo study of viral haemorrhagic septicaemia virus and infectious pancreatic necrosis virus coexistence in Senegalese sole (Solea senegalensis). J. Fish Dis. 2016, 40, 1129–1139. [Google Scholar] [CrossRef]
- Cyrille, D.; Nick, S.L.; Ted, K.; David, Z.; Reza, K.; Damian, C.E.; Jeong, H.L.; Zoltan, M.; Miriam, V.B.; Mandy, J.; et al. Highly conserved protective epitopes on influenza B viruses. Science 2012, 337, 1343–1348. [Google Scholar]
- Bonnichon, V.; Lightner, D.V.; Bonami, J.R. Viral interference between infectious hypodermal and hematopoietic necrosis virus and white spot syndrome virus in Litopenaeus vannamei. Dis. Aquat. Organ. 2006, 72, 179–184. [Google Scholar] [CrossRef] [PubMed]
- Imre, K.; Tibor, B. Therapeutic Exploitation of Viral Interference. Infect. Disord. Drug Targets 2020, 20, 423–432. [Google Scholar]
- Alonso, M.; Rodríguez, S.; Pérez-Prieto, S.I. Viral coinfection in salmonids: Infectious pancreatic necrosis virus interferes with infectious hematopoietic necrosis virus. Arch. Virol. 1999, 144, 657–673. [Google Scholar] [CrossRef]
- Wolf, K. Fish Viruses and Fish Viral Diseases; Cornell University Press: Ithaca, NY, USA, 1988. [Google Scholar]
- Woo, P.T.; Bruno, D. Viral, bacterial and fungal infections. In Fish Diseases and Disorders; CABI: Oxfordshire, UK, 1999; Volume 3. [Google Scholar]
- Sinha, A.K.; Liew, H.J.; Diricx, M.; Blust, R.; Gudrun, D.B. The interactive effects of ammonia exposure, nutritional status and exercise on metabolic and physiological responses in gold fish (Carassius auratus L.). Aquat. Toxicol. 2012, 109, 33–46. [Google Scholar] [CrossRef]
- Graham, A.L.; Cattadori, I.M.; Lloyd-Smith, J.O.; Ferrari, M.J.; Bjørnstad, O.N. Transmission consequences of coinfection: Cytokines writ large? Trends Parasitol. 2007, 23, 284–291. [Google Scholar] [CrossRef]
- Telfer, S.; Birtles, R.; Bennett, M.; Lambin, X.; Paterson, S.; Begon, M. Parasite interactions in natural populations: Insights from longitudinal data. Parasitology 2008, 135, 767–781. [Google Scholar] [CrossRef]
- Lello, J.; Boag, B.; Fenton, A.; Stevenson, I.R.; Hudson, P.J. Competition and mutualism among the gut helminths of a mammalian host. Nature 2004, 428, 840–844. [Google Scholar] [CrossRef]
- García, I.; Galiana, A.; Falcó, A.; Estepa, A.; Perez, L. Characterization of an infectious pancreatic necrosis (IPN) virus carrier cell culture with resistance to superinfection with heterologous viruses. Vet. Microbiol. 2011, 149, 48–55. [Google Scholar] [CrossRef] [PubMed]
- Kotob, M.H.; Menanteau-Ledouble, S.; Kumar, G.; Abdelzaher, M.; El-Matbouli, M. The impact of co-infections on fish: A review. Vet. Res. 2016, 47, 98. [Google Scholar] [CrossRef]
- Bootland, L.M.; Leong, J. Infectious haematopoietic necrosis virus. Fish Dis. Disord. 2011, 3, 66–110. [Google Scholar]
- So, S.K.; Kwang, K.; Hae, K.Y.; Yu, S.H.; Myeong, E.J.; Soon, G.B.; Hyun, J.L.; Jang, S.P.; Yung, J.K. Differential virulence of infectious hematopoietic necrosis virus (IHNV) isolated from salmonid fish in Gangwon Province, Korea. Fish Shellfish Immunol. 2021, 119, 490–498. [Google Scholar]
- Zou, J.; Gorgoglione, B.; Taylor, N.G.H.; Summathed, T.; Lee, O.T.; Panigrahi, A.; Genet, C.; Chen, Y.M.; Chen, T.Y.; Hassan, M.U.; et al. Salmonids Have an Extraordinary Complex Type I IFN System: Characterization of the IFN Locus in Rainbow Trout Oncorhynchus mykiss Reveals Two Novel IFN Subgroups. J. Immunol. 2014, 193, 2273–2286. [Google Scholar] [CrossRef] [PubMed]
- Schoggins, J.W.; Rice, C.M. Interferon-stimulated genes and their antiviral effector functions. Curr. Opin. Virol. 2011, 1, 519–525. [Google Scholar] [CrossRef] [PubMed]
- Durfee, L.A.; Lyon, N.; Seo, K.; Huibregtse, J.M. The ISG15 Conjugation System Broadly Targets Newly Synthesized Proteins: Implications for the Antiviral Function of ISG15. Mol. Cell. 2010, 38, 722–732. [Google Scholar] [CrossRef] [PubMed]
- Nombela, I.; Carrion, A.; Puente-Marin, S.; Chico, V.; Mercado, L.; Perez, L.; Coll, J.; Ortega-Villaizan, M.D.M. Infectious pancreatic necrosis virus triggers antiviral immune response in rainbow trout red blood cells, despite not being infective. F1000Research 2017, 6, 1968. [Google Scholar] [CrossRef]
- Crosse, K.M.; Monson, E.A.; Beard, M.R.; Helbig, K.J. Interferon-Stimulated Genes as Enhancers of Antiviral Innate Immune Signaling. J. Innate. Immun. 2018, 10, 85–93. [Google Scholar] [CrossRef]











Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shao, Y.; Zhao, J.; Ren, G.; Lu, T.; Chen, X.; Xu, L. Early or Simultaneous Infection with Infectious Pancreatic Necrosis Virus Inhibits Infectious Hematopoietic Necrosis Virus Replication and Induces a Stronger Antiviral Response during Co-infection in Rainbow Trout (Oncorhynchus mykiss). Viruses 2022, 14, 1732. https://doi.org/10.3390/v14081732
Shao Y, Zhao J, Ren G, Lu T, Chen X, Xu L. Early or Simultaneous Infection with Infectious Pancreatic Necrosis Virus Inhibits Infectious Hematopoietic Necrosis Virus Replication and Induces a Stronger Antiviral Response during Co-infection in Rainbow Trout (Oncorhynchus mykiss). Viruses. 2022; 14(8):1732. https://doi.org/10.3390/v14081732
Chicago/Turabian StyleShao, Yizhi, Jingzhuang Zhao, Guangming Ren, Tongyan Lu, Xiaoyu Chen, and Liming Xu. 2022. "Early or Simultaneous Infection with Infectious Pancreatic Necrosis Virus Inhibits Infectious Hematopoietic Necrosis Virus Replication and Induces a Stronger Antiviral Response during Co-infection in Rainbow Trout (Oncorhynchus mykiss)" Viruses 14, no. 8: 1732. https://doi.org/10.3390/v14081732