
Female Genital Fibroblasts Diminish the In Vitro Efficacy of PrEP against HIV

 , , , ,
, , , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Isolation and Culture of Endometrial Stromal Fibroblasts (eSFs)
2.2. Isolation and Culture of Peripheral Blood Mononuclear Cells (PBMCs)
2.3. Virus Preparation
2.4. Fibroblast Co-Culture Infection Assays
2.5. Flow Cytometry
2.6. Quantitation of Intracellular Tenofovir-Diphosphate (TVF-dp) by Mass Spectometry
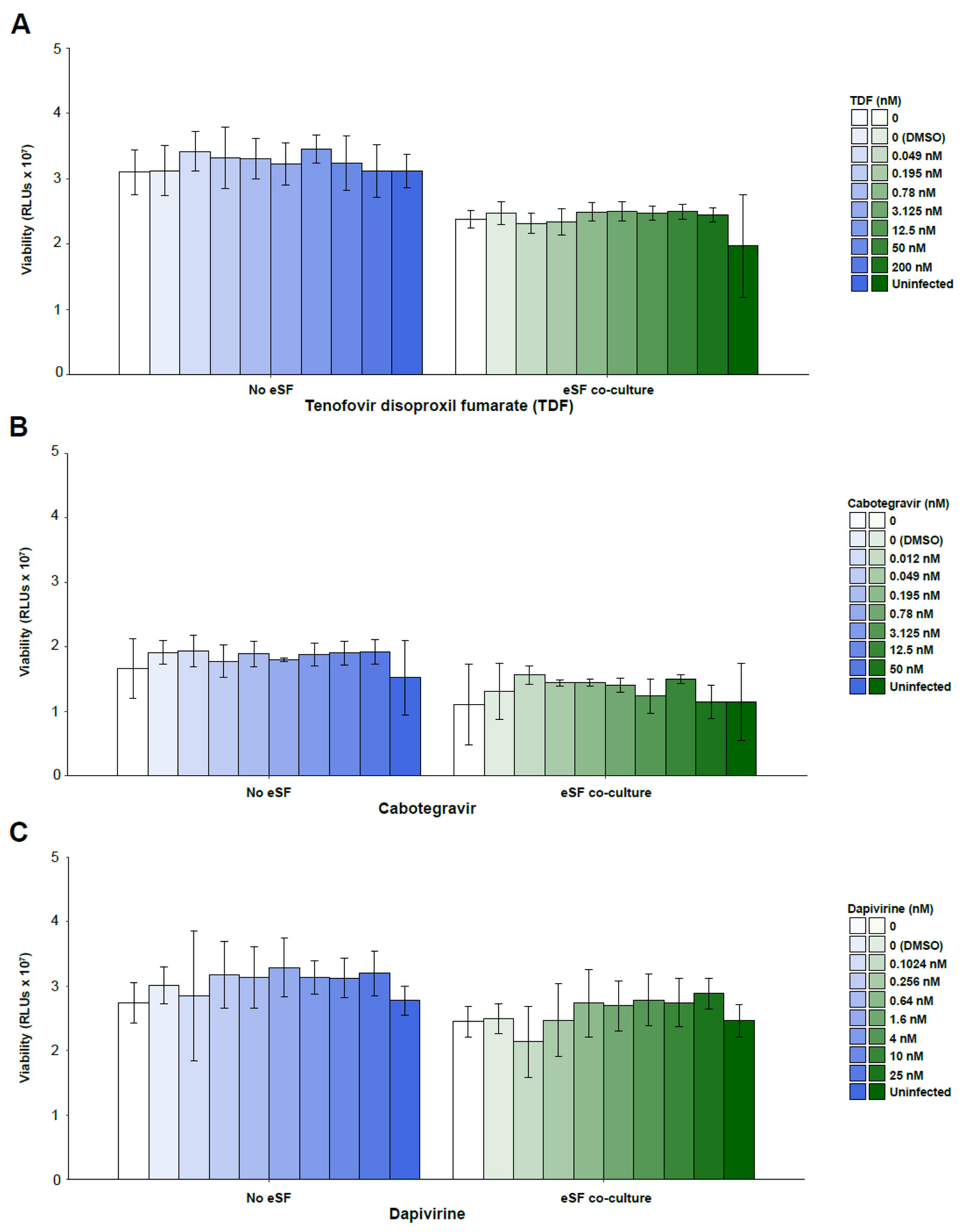
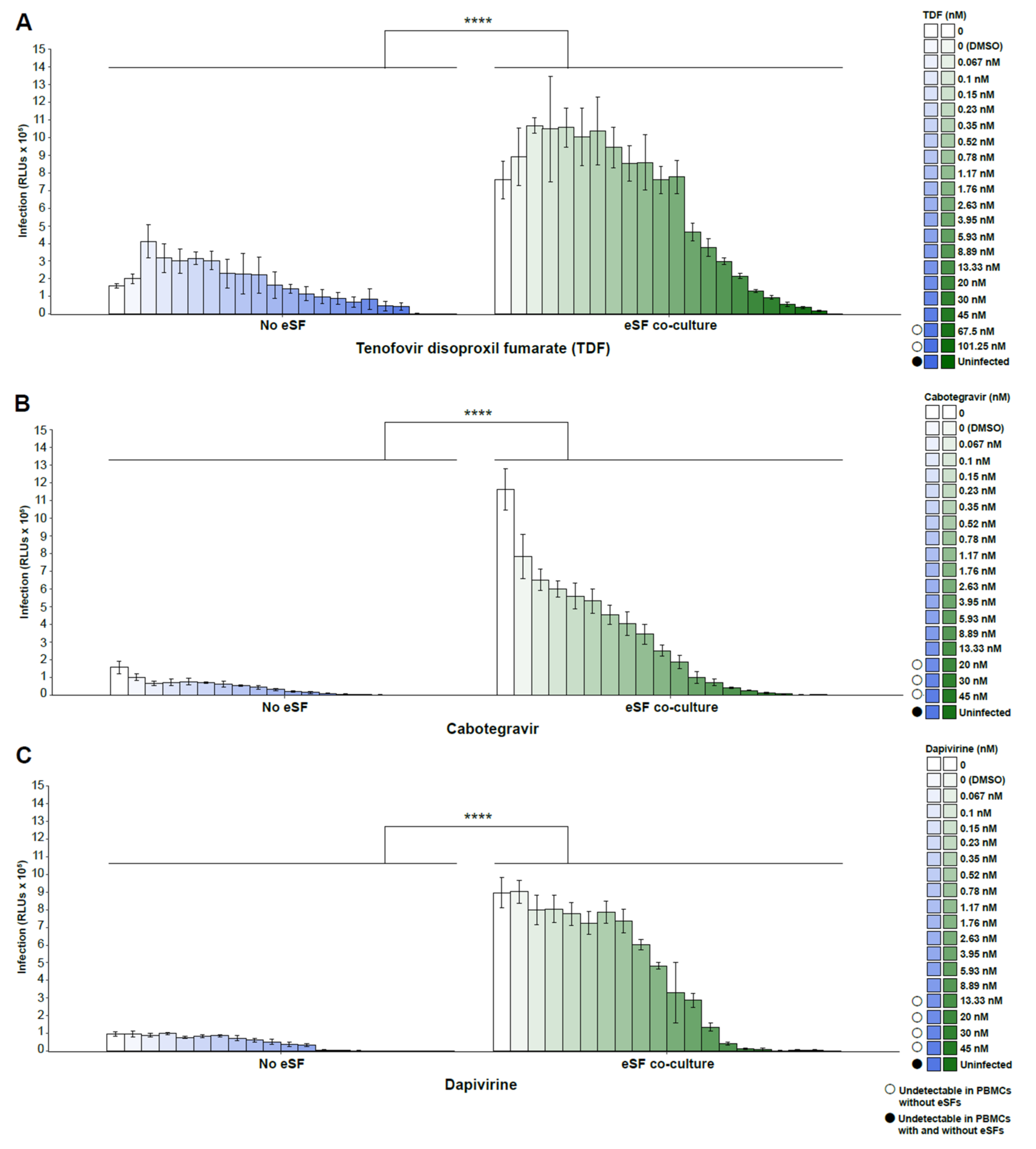
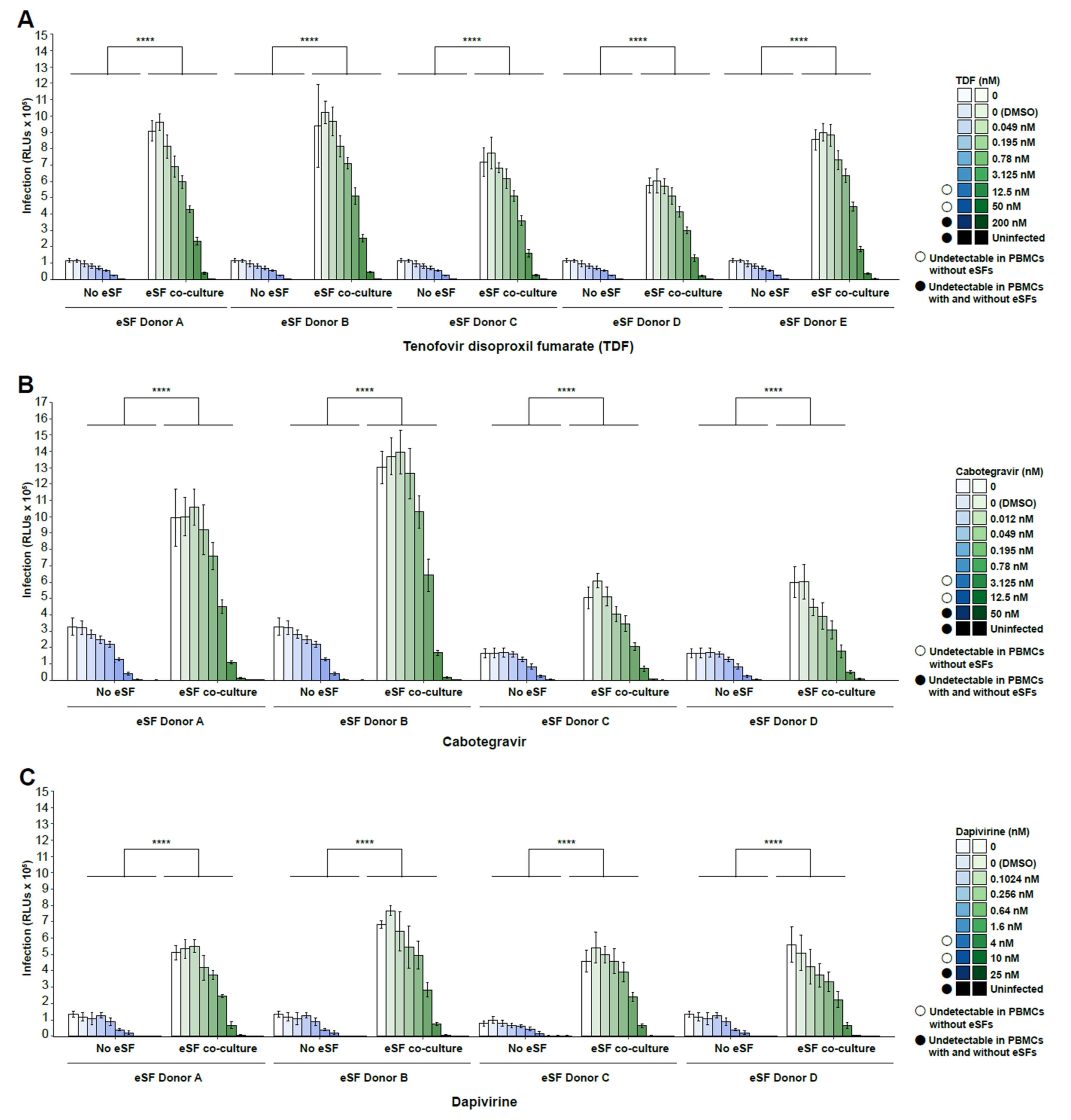
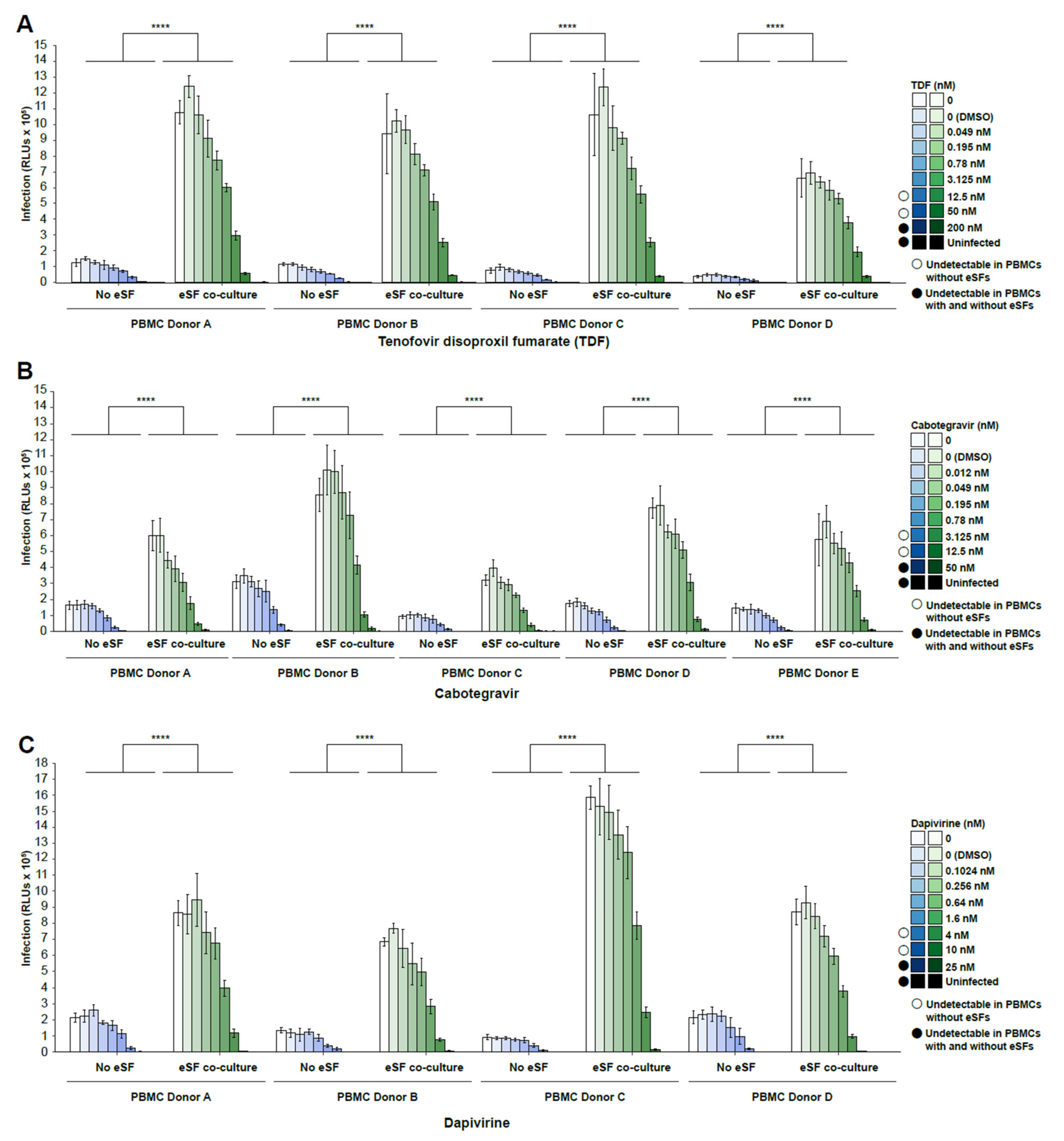
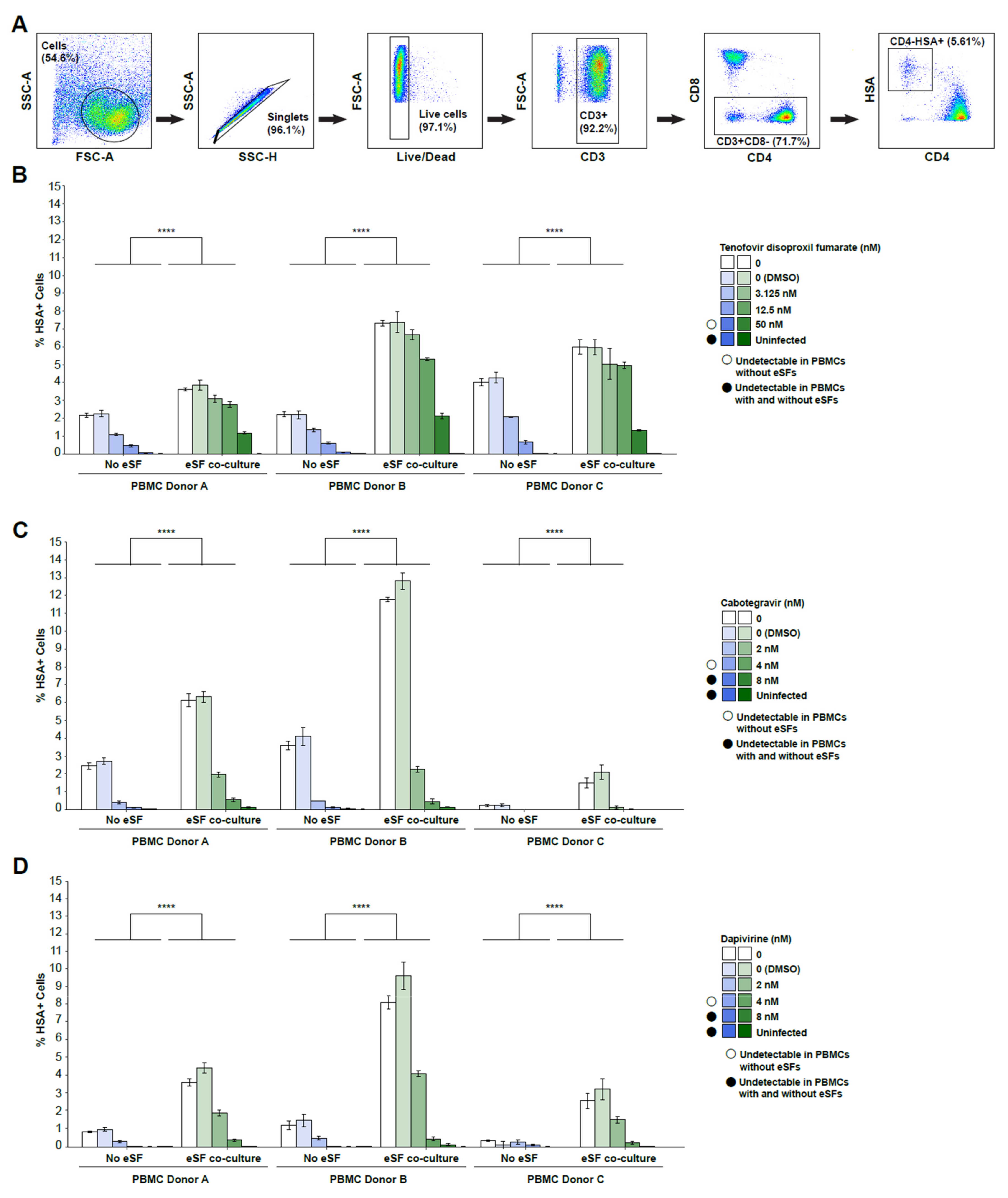
3. Results
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- UNAIDS Global Factsheets 2020. Available online: https://aidsinfo.unaids.org/ (accessed on 21 July 2022).
- Delany-Moretlwe, S.; Hughes, J.P.; Bock, P.; Ouma, S.G.; Hunidzarira, P.; Kalonji, D.; Kayange, N.; Makhema, J.; Mandima, P.; Mathew, C.; et al. Cabotegravir for the prevention of HIV-1 in women: Results from HPTN 084, a phase 3, randomised clinical trial. Lancet 2022, 399, 1779–1789. [Google Scholar] [CrossRef]
- Thomson, K.A.; Baeten, J.M.; Mugo, N.R.; Bekker, L.G.; Celum, C.L.; Heffron, R. Tenofovir-based oral preexposure prophylaxis prevents HIV infection among women. Curr. Opin. HIV AIDS 2016, 11, 18–26. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abdool Karim, Q.; Abdool Karim, S.S.; Frohlich, J.A.; Grobler, A.C.; Baxter, C.; Mansoor, L.E.; Kharsany, A.B.; Sibeko, S.; Mlisana, K.P.; Omar, Z.; et al. Effectiveness and safety of tenofovir gel, an antiretroviral microbicide, for the prevention of HIV infection in women. Science 2010, 329, 1168–1174. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cottrell, M.L.; Yang, K.H.; Prince, H.M.; Sykes, C.; White, N.; Malone, S.; Dellon, E.S.; Madanick, R.D.; Shaheen, N.J.; Hudgens, M.G.; et al. A Translational Pharmacology Approach to Predicting Outcomes of Preexposure Prophylaxis Against HIV in Men and Women Using Tenofovir Disoproxil Fumarate With or Without Emtricitabine. J. Infect. Dis. 2016, 214, 55–64. [Google Scholar] [CrossRef] [PubMed]
- Anderson, P.L.; Glidden, D.V.; Liu, A.; Buchbinder, S.; Lama, J.R.; Guanira, J.V.; McMahan, V.; Bushman, L.R.; Casapia, M.; Montoya-Herrera, O.; et al. Emtricitabine-tenofovir concentrations and pre-exposure prophylaxis efficacy in men who have sex with men. Sci. Transl. Med. 2012, 4, 151ra125. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hendrix, C.W. Exploring concentration response in HIV pre-exposure prophylaxis to optimize clinical care and trial design. Cell 2013, 155, 515–518. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baeten, J.M.; Palanee-Phillips, T.; Mgodi, N.M.; Mayo, A.J.; Szydlo, D.W.; Ramjee, G.; Gati Mirembe, B.; Mhlanga, F.; Hunidzarira, P.; Mansoor, L.E.; et al. Safety, uptake, and use of a dapivirine vaginal ring for HIV-1 prevention in African women (HOPE): An open-label, extension study. Lancet HIV 2021, 8, e87–e95. [Google Scholar] [CrossRef]
- Baeten, J.M.; Palanee-Phillips, T.; Brown, E.R.; Schwartz, K.; Soto-Torres, L.E.; Govender, V.; Mgodi, N.M.; Matovu Kiweewa, F.; Nair, G.; Mhlanga, F.; et al. Use of a Vaginal Ring Containing Dapivirine for HIV-1 Prevention in Women. N. Engl. J. Med. 2016, 375, 2121–2132. [Google Scholar] [CrossRef]
- Nel, A.; van Niekerk, N.; Van Baelen, B.; Malherbe, M.; Mans, W.; Carter, A.; Steytler, J.; van der Ryst, E.; Craig, C.; Louw, C.; et al. Safety, adherence, and HIV-1 seroconversion among women using the dapivirine vaginal ring (DREAM): An open-label, extension study. Lancet HIV 2021, 8, e77–e86. [Google Scholar] [CrossRef]
- Hendrix, C.W.; Chen, B.A.; Guddera, V.; Hoesley, C.; Justman, J.; Nakabiito, C.; Salata, R.; Soto-Torres, L.; Patterson, K.; Minnis, A.M.; et al. MTN-001: Randomized pharmacokinetic cross-over study comparing tenofovir vaginal gel and oral tablets in vaginal tissue and other compartments. PLoS ONE 2013, 8, e55013. [Google Scholar] [CrossRef] [Green Version]
- Thurman, A.R.; Schwartz, J.L.; Cottrell, M.L.; Brache, V.; Chen, B.A.; Cochon, L.; Ju, S.; McGowan, I.; Rooney, J.F.; McCallister, S.; et al. Safety and Pharmacokinetics of a Tenofovir Alafenamide Fumarate-Emtricitabine based Oral Antiretroviral Regimen for Prevention of HIV Acquisition in Women: A Randomized Controlled Trial. eClinicalMedicine 2021, 36, 100893. [Google Scholar] [CrossRef]
- Klatt, N.R.; Cheu, R.; Birse, K.; Zevin, A.S.; Perner, M.; Noel-Romas, L.; Grobler, A.; Westmacott, G.; Xie, I.Y.; Butler, J.; et al. Vaginal bacteria modify HIV tenofovir microbicide efficacy in African women. Science 2017, 356, 938–945. [Google Scholar] [CrossRef] [Green Version]
- Cheu, R.K.; Gustin, A.T.; Lee, C.; Schifanella, L.; Miller, C.J.; Ha, A.; Kim, C.; Rodriguez, V.J.; Fischl, M.; Burgener, A.D.; et al. Impact of vaginal microbiome communities on HIV antiretroviral-based pre-exposure prophylaxis (PrEP) drug metabolism. PLoS Pathog. 2020, 16, e1009024. [Google Scholar] [CrossRef]
- Masson, L.; Mlisana, K.; Little, F.; Werner, L.; Mkhize, N.N.; Ronacher, K.; Gamieldien, H.; Williamson, C.; McKinnon, L.R.; Walzl, G.; et al. Defining genital tract cytokine signatures of sexually transmitted infections and bacterial vaginosis in women at high risk of HIV infection: A cross-sectional study. Sex. Transm. Infect. 2014, 90, 580–587. [Google Scholar] [CrossRef]
- Galvin, S.R.; Cohen, M.S. The role of sexually transmitted diseases in HIV transmission. Nat. Rev. Microbiol. 2004, 2, 33–42. [Google Scholar] [CrossRef]
- Masson, L.; Passmore, J.A.; Liebenberg, L.J.; Werner, L.; Baxter, C.; Arnold, K.B.; Williamson, C.; Little, F.; Mansoor, L.E.; Naranbhai, V.; et al. Genital inflammation and the risk of HIV acquisition in women. Clin. Infect. Dis. 2015, 61, 260–269. [Google Scholar] [CrossRef] [Green Version]
- Passmore, J.A.; Jaspan, H.B.; Masson, L. Genital inflammation, immune activation and risk of sexual HIV acquisition. Curr. Opin. HIV AIDS 2016, 11, 156–162. [Google Scholar] [CrossRef]
- Munch, J.; Rucker, E.; Standker, L.; Adermann, K.; Goffinet, C.; Schindler, M.; Wildum, S.; Chinnadurai, R.; Rajan, D.; Specht, A.; et al. Semen-derived amyloid fibrils drastically enhance HIV infection. Cell 2007, 131, 1059–1071. [Google Scholar] [CrossRef] [Green Version]
- Kim, K.A.; Yolamanova, M.; Zirafi, O.; Roan, N.R.; Staendker, L.; Forssmann, W.G.; Burgener, A.; Dejucq-Rainsford, N.; Hahn, B.H.; Shaw, G.M.; et al. Semen-mediated enhancement of HIV infection is donor-dependent and correlates with the levels of SEVI. Retrovirology 2010, 7, 55. [Google Scholar] [CrossRef] [Green Version]
- Roan, N.R.; Muller, J.A.; Liu, H.; Chu, S.; Arnold, F.; Sturzel, C.M.; Walther, P.; Dong, M.; Witkowska, H.E.; Kirchhoff, F.; et al. Peptides Released by Physiological Cleavage of Semen Coagulum Proteins Form Amyloids that Enhance HIV Infection. Cell Host Microbe 2011, 10, 541–550. [Google Scholar] [CrossRef] [Green Version]
- Roan, N.R.; Liu, H.; Usmani, S.M.; Neidleman, J.; Muller, J.A.; Avila-Herrera, A.; Gawanbacht, A.; Zirafi, O.; Chu, S.; Dong, M.; et al. Liquefaction of Semen Generates and Later Degrades a Conserved Semenogelin Peptide That Enhances HIV Infection. J. Virol. 2014, 88, 7221–7234. [Google Scholar] [CrossRef] [Green Version]
- Usmani, S.M.; Zirafi, O.; Muller, J.A.; Sandi-Monroy, N.L.; Yadav, J.K.; Meier, C.; Weil, T.; Roan, N.R.; Greene, W.C.; Walther, P.; et al. Direct visualization of HIV-enhancing endogenous amyloid fibrils in human semen. Nat. Commun. 2014, 5, 3508. [Google Scholar] [CrossRef] [Green Version]
- Roan, N.R.; Jakobsen, M.R. Friend or Foe: Innate Sensing of HIV in the Female Reproductive Tract. Curr. HIV/AIDS Rep. 2016, 13, 53–63. [Google Scholar] [CrossRef] [Green Version]
- Leyendecker, G.; Kunz, G.; Wildt, L.; Beil, D.; Deininger, H. Uterine hyperperistalsis and dysperistalsis as dysfunctions of the mechanism of rapid sperm transport in patients with endometriosis and infertility. Hum. Reprod. 1996, 11, 1542–1551. [Google Scholar] [CrossRef] [Green Version]
- Barnhart, K.T.; Stolpen, A.; Pretorius, E.S.; Malamud, D. Distribution of a spermicide containing Nonoxynol-9 in the vaginal canal and the upper female reproductive tract. Hum. Reprod. 2001, 16, 1151–1154. [Google Scholar] [CrossRef] [Green Version]
- Stieh, D.J.; Maric, D.; Kelley, Z.L.; Anderson, M.R.; Hattaway, H.Z.; Beilfuss, B.A.; Rothwangl, K.B.; Veazey, R.S.; Hope, T.J. Vaginal challenge with an SIV-based dual reporter system reveals that infection can occur throughout the upper and lower female reproductive tract. PLoS Pathog. 2014, 10, e1004440. [Google Scholar] [CrossRef]
- Stieh, D.J.; Matias, E.; Xu, H.; Fought, A.J.; Blanchard, J.L.; Marx, P.A.; Veazey, R.S.; Hope, T.J. Th17 Cells Are Preferentially Infected Very Early after Vaginal Transmission of SIV in Macaques. Cell Host Microbe 2016, 19, 529–540. [Google Scholar] [CrossRef] [Green Version]
- Li, Q.; Estes, J.D.; Schlievert, P.M.; Duan, L.; Brosnahan, A.J.; Southern, P.J.; Reilly, C.S.; Peterson, M.L.; Schultz-Darken, N.; Brunner, K.G.; et al. Glycerol monolaurate prevents mucosal SIV transmission. Nature 2009, 458, 1034–1038. [Google Scholar] [CrossRef]
- Quillay, H.; El Costa, H.; Marlin, R.; Duriez, M.; Cannou, C.; Chretien, F.; Fernandez, H.; Lebreton, A.; Ighil, J.; Schwartz, O.; et al. Distinct endometrial and decidual macrophage characteristics and regulation of their permissivity to HIV-1 infection by SAMHD1. J. Virol. 2015, 89, 1329–1339. [Google Scholar] [CrossRef] [Green Version]
- Neidleman, J.A.; Chen, J.C.; Kohgadai, N.; Muller, J.A.; Laustsen, A.; Thavachelvam, K.; Jang, K.S.; Sturzel, C.M.; Jones, J.J.; Ochsenbauer, C.; et al. Mucosal stromal fibroblasts markedly enhance HIV infection of CD4+ T cells. PLoS Pathog. 2017, 13, e1006163. [Google Scholar] [CrossRef]
- Krjutskov, K.; Katayama, S.; Saare, M.; Vera-Rodriguez, M.; Lubenets, D.; Samuel, K.; Laisk-Podar, T.; Teder, H.; Einarsdottir, E.; Salumets, A.; et al. Single-cell transcriptome analysis of endometrial tissue. Hum. Reprod. 2016, 31, 844–853. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Spitzer, T.L.; Rojas, A.; Zelenko, Z.; Aghajanova, L.; Erikson, D.W.; Barragan, F.; Meyer, M.; Tamaresis, J.S.; Hamilton, A.E.; Irwin, J.C.; et al. Perivascular human endometrial mesenchymal stem cells express pathways relevant to self-renewal, lineage specification, and functional phenotype. Biol. Reprod. 2012, 86, 58. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.C.; Roan, N.R. Isolation and culture of human endometrial epithelial cells and stromal fibroblasts. Bio-Protocol 2015, 5, e1623. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hiebenthal-Millow, K.; Kirchhoff, F. The most frequent naturally occurring length polymorphism in the HIV-1 LTR has little effect on proviral transcription and viral replication. Virology 2002, 292, 169–175. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cavrois, M.; Banerjee, T.; Mukherjee, G.; Raman, N.; Hussien, R.; Rodriguez, B.A.; Vasquez, J.; Spitzer, M.H.; Lazarus, N.H.; Jones, J.J.; et al. Mass Cytometric Analysis of HIV Entry, Replication, and Remodeling in Tissue CD4+ T Cells. Cell Rep. 2017, 20, 984–998. [Google Scholar] [CrossRef] [Green Version]
- Egedal, J.H.; Xie, G.; Packard, T.A.; Laustsen, A.; Neidleman, J.; Georgiou, K.; Pillai, S.K.; Greene, W.C.; Jakobsen, M.R.; Roan, N.R. Hyaluronic acid is a negative regulator of mucosal fibroblast-mediated enhancement of HIV infection. Mucosal Immunol. 2021, 14, 1203–1213. [Google Scholar] [CrossRef]
- Garcia, J.V.; Miller, A.D. Serine phosphorylation-independent downregulation of cell-surface CD4 by nef. Nature 1991, 350, 508–511. [Google Scholar] [CrossRef]
- Vincent, M.J.; Raja, N.U.; Jabbar, M.A. Human immunodeficiency virus type 1 Vpu protein induces degradation of chimeric envelope glycoproteins bearing the cytoplasmic and anchor domains of CD4: Role of the cytoplasmic domain in Vpu-induced degradation in the endoplasmic reticulum. J. Virol. 1993, 67, 5538–5549. [Google Scholar] [CrossRef] [Green Version]
- Zirafi, O.; Kim, K.A.; Roan, N.R.; Kluge, S.F.; Muller, J.A.; Jiang, S.; Mayer, B.; Greene, W.C.; Kirchhoff, F.; Munch, J. Semen enhances HIV infectivity and impairs the antiviral efficacy of microbicides. Sci. Transl. Med. 2014, 6, 262ra157. [Google Scholar] [CrossRef] [Green Version]
- Sharkey, D.J.; Macpherson, A.M.; Tremellen, K.P.; Mottershead, D.G.; Gilchrist, R.B.; Robertson, S.A. TGF-beta mediates proinflammatory seminal fluid signaling in human cervical epithelial cells. J. Immunol. 2012, 189, 1024–1035. [Google Scholar] [CrossRef] [Green Version]
- Sharkey, D.J.; Macpherson, A.M.; Tremellen, K.P.; Robertson, S.A. Seminal plasma differentially regulates inflammatory cytokine gene expression in human cervical and vaginal epithelial cells. Mol. Hum. Reprod. 2007, 13, 491–501. [Google Scholar] [CrossRef] [Green Version]
- Sharkey, D.J.; Tremellen, K.P.; Jasper, M.J.; Gemzell-Danielsson, K.; Robertson, S.A. Seminal fluid induces leukocyte recruitment and cytokine and chemokine mRNA expression in the human cervix after coitus. J. Immunol. 2012, 188, 2445–2454. [Google Scholar] [CrossRef] [Green Version]
- Marques de Menezes, E.G.; Jang, K.; George, A.F.; Nyegaard, M.; Neidleman, J.; Inglis, H.C.; Danesh, A.; Deng, X.; Afshari, A.; Kim, Y.H.; et al. Seminal Plasma-Derived Extracellular-Vesicle Fractions from HIV-Infected Men Exhibit Unique MicroRNA Signatures and Induce a Proinflammatory Response in Cells Isolated from the Female Reproductive Tract. J. Virol. 2020, 94, e00525-20. [Google Scholar] [CrossRef]
- Chen, J.C.; Johnson, B.A.; Erikson, D.W.; Piltonen, T.T.; Barragan, F.; Chu, S.; Kohgadai, N.; Irwin, J.C.; Greene, W.C.; Giudice, L.C.; et al. Seminal plasma induces global transcriptomic changes associated with cell migration, proliferation and viability in endometrial epithelial cells and stromal fibroblasts. Hum. Reprod. 2014, 29, 1255–1270. [Google Scholar] [CrossRef] [Green Version]
- Shannon, B.; Gajer, P.; Yi, T.J.; Ma, B.; Humphrys, M.S.; Thomas-Pavanel, J.; Chieza, L.; Janakiram, P.; Saunders, M.; Tharao, W.; et al. Distinct Effects of the Cervicovaginal Microbiota and Herpes Simplex Type 2 Infection on Female Genital Tract Immunology. J. Infect. Dis. 2017, 215, 1366–1375. [Google Scholar] [CrossRef] [Green Version]
- Liebenberg, L.J.P.; McKinnon, L.R.; Yende-Zuma, N.; Garrett, N.; Baxter, C.; Kharsany, A.B.M.; Archary, D.; Rositch, A.; Samsunder, N.; Mansoor, L.E.; et al. HPV infection and the genital cytokine milieu in women at high risk of HIV acquisition. Nat. Commun. 2019, 10, 5227. [Google Scholar] [CrossRef] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample | TDF Concentration | TVF-dp Concentration Median (IQR) a |
|---|---|---|
| PBMCs | 50 nM | 0.79 ng/mL (0.62, 1.04) |
| 200 nM | 3.50 ng/mL (1.81, 3.82) | |
| 800 nM | 18.00 ng/mL (11.46, 20.50) | |
| PBMCs (co-cultured with eSFs) | 50 nM | 0.56 ng/mL (0.26, 1.30) |
| 200 nM | 2.71 ng/mL (1.60, 6.04) | |
| 800 nM | 12.77 ng/mL (6.56, 29.73) | |
| eSFs (co-cultured with PBMCs) | 50 nM | 0.53 ng/mL (<LLOQ, 0.58) |
| 200 nM | 0.87 ng/mL (<LLOQ, 1.81) | |
| 800 nM | 3.98 ng/mL (0.79, 9.27) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
George, A.F.; McGregor, M.; Gingrich, D.; Neidleman, J.; Marquez, R.S.; Young, K.C.; Thanigaivelan, K.L.; Greene, W.C.; Tien, P.C.; Deitchman, A.N.; et al. Female Genital Fibroblasts Diminish the In Vitro Efficacy of PrEP against HIV. Viruses 2022, 14, 1723. https://doi.org/10.3390/v14081723
George AF, McGregor M, Gingrich D, Neidleman J, Marquez RS, Young KC, Thanigaivelan KL, Greene WC, Tien PC, Deitchman AN, et al. Female Genital Fibroblasts Diminish the In Vitro Efficacy of PrEP against HIV. Viruses. 2022; 14(8):1723. https://doi.org/10.3390/v14081723
Chicago/Turabian StyleGeorge, Ashley F., Matthew McGregor, David Gingrich, Jason Neidleman, Rebecca S. Marquez, Kyrlia C. Young, Kaavya L. Thanigaivelan, Warner C. Greene, Phyllis C. Tien, Amelia N. Deitchman, and et al. 2022. "Female Genital Fibroblasts Diminish the In Vitro Efficacy of PrEP against HIV" Viruses 14, no. 8: 1723. https://doi.org/10.3390/v14081723