

Analysis of Viral and Host Factors on Immunogenicity of 2018, 2019, and 2020 Southern Hemisphere Seasonal Trivalent Inactivated Influenza Vaccine in Adults in Brazil
 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
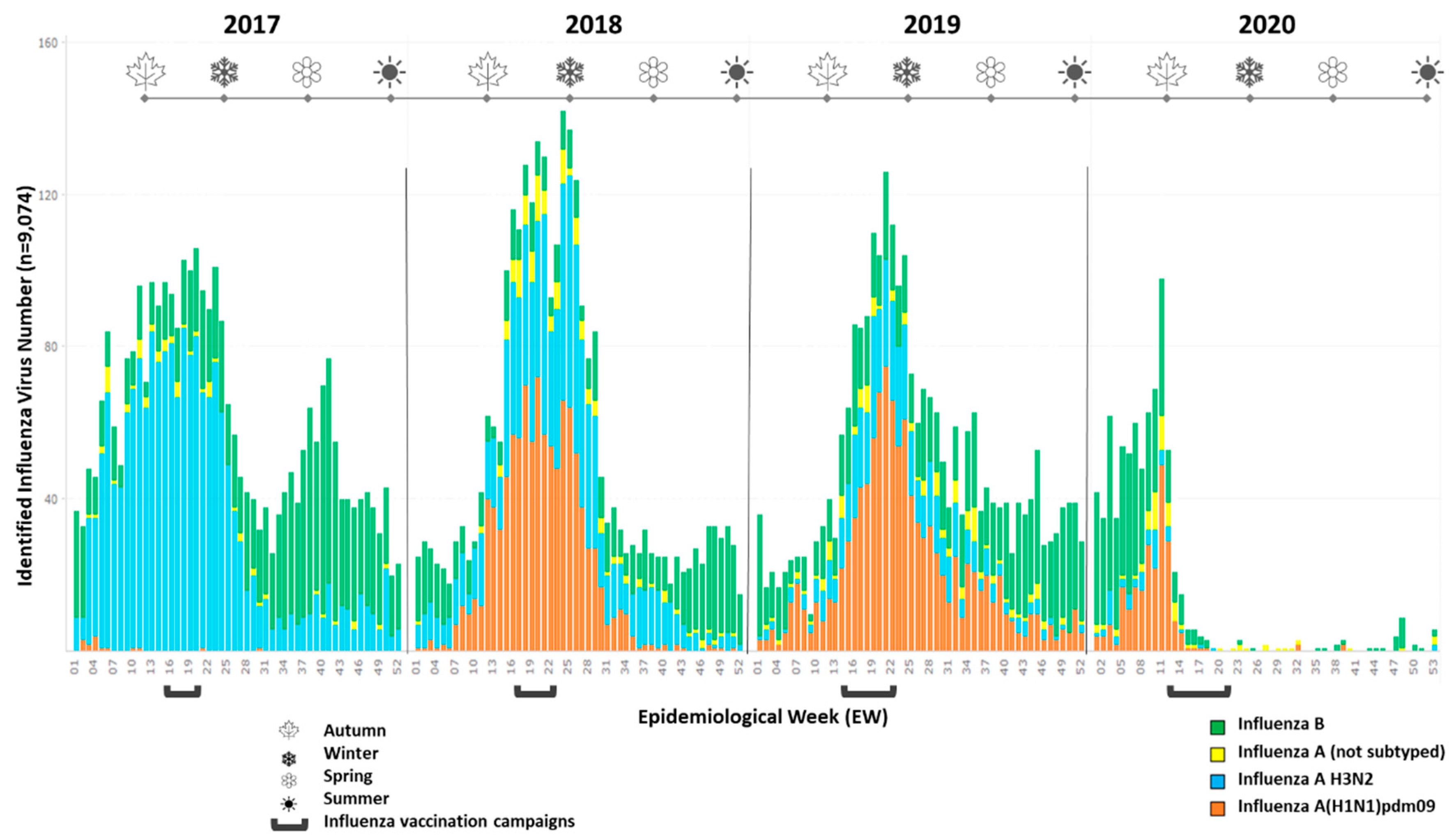
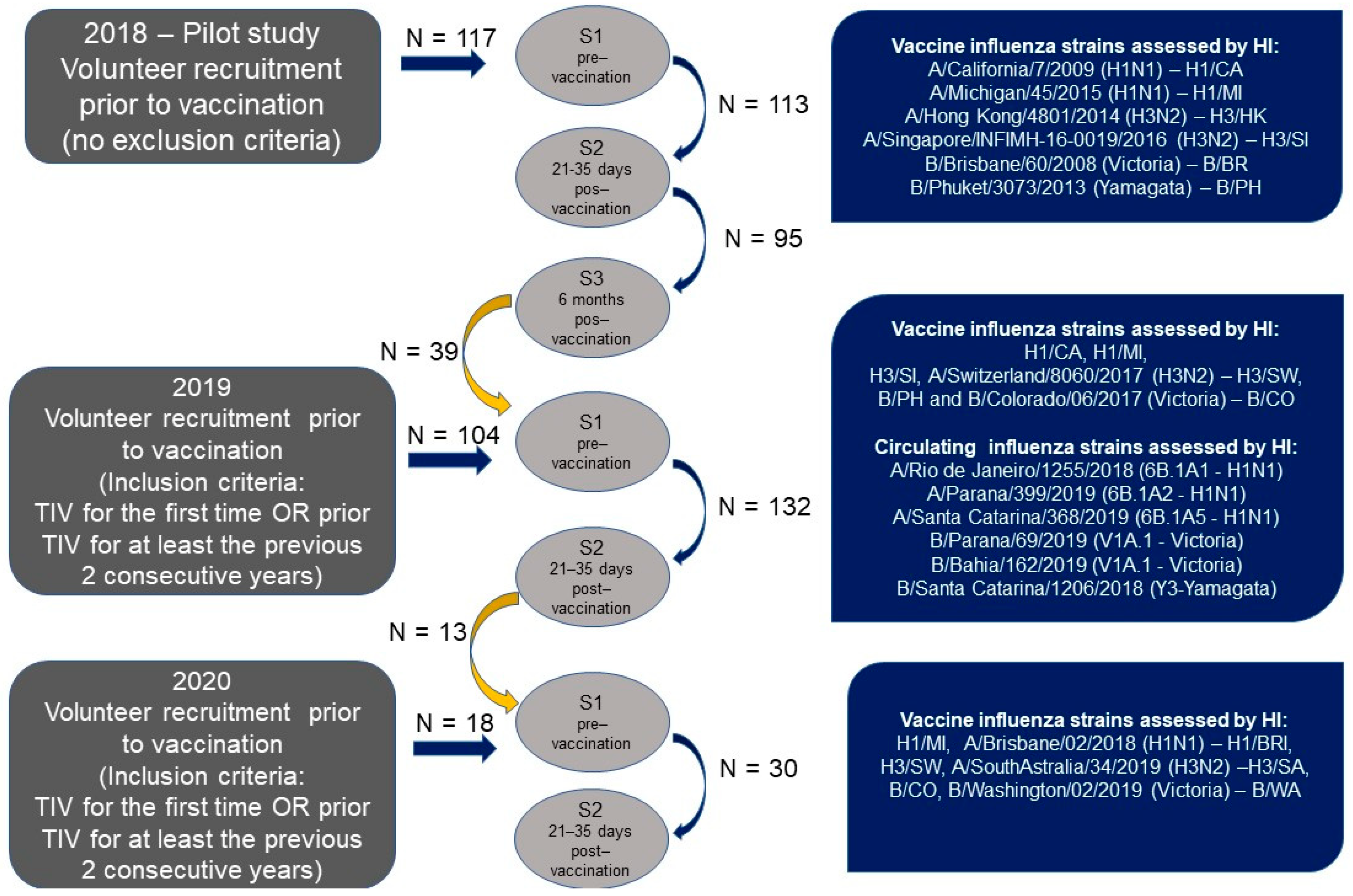
2.1. Study Design and Sample Collection
2.2. Virus Isolation
2.3. Serum Treatment and Hemagglutination Inhibition Assay
2.4. Patient Follow-Up
2.5. Statistical Analysis
3. Results
3.1. Study Population
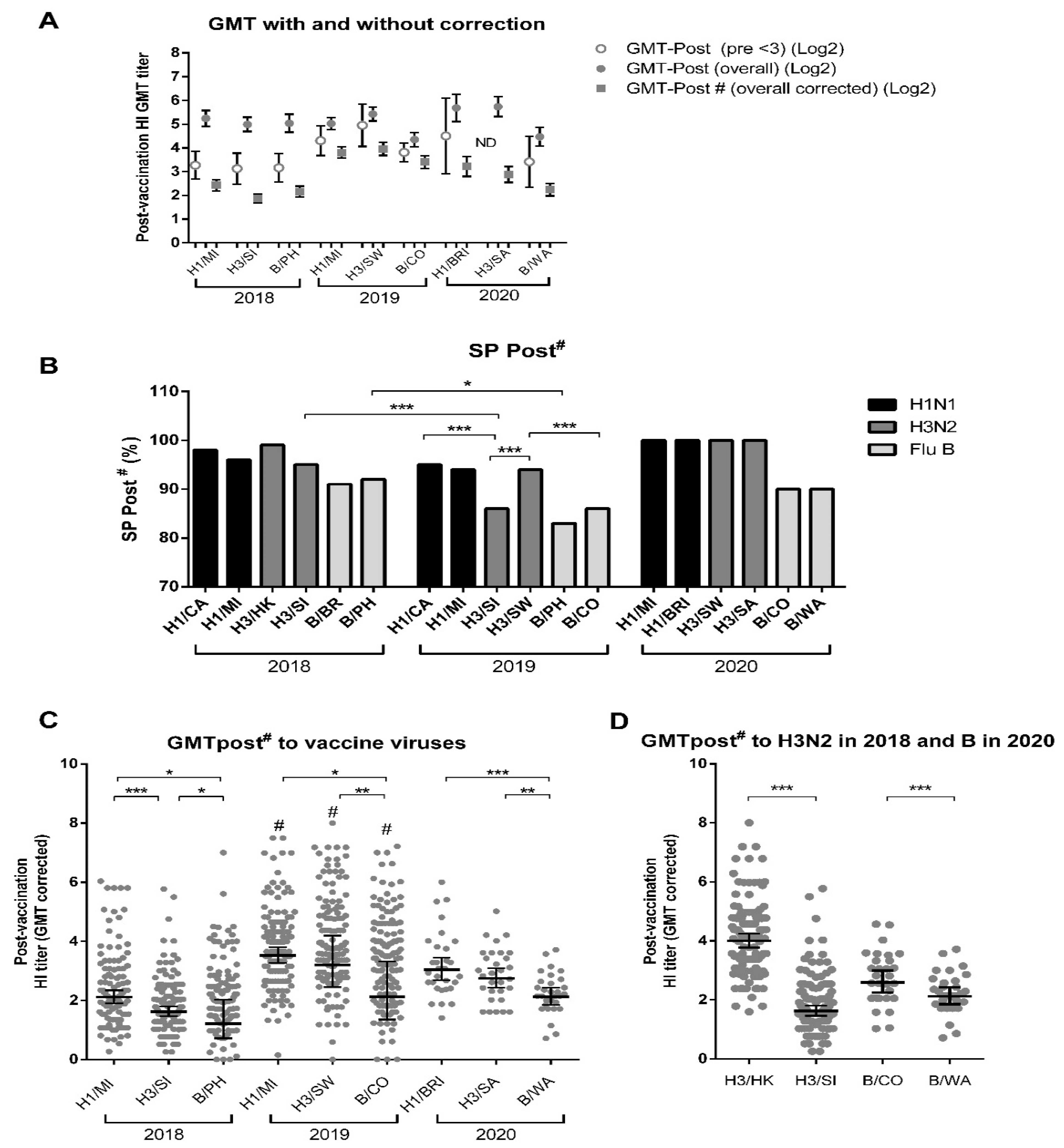
3.2. TIV Enhances HI Titers and Seroprotection for Current and Previous Years’ Vaccine Strains
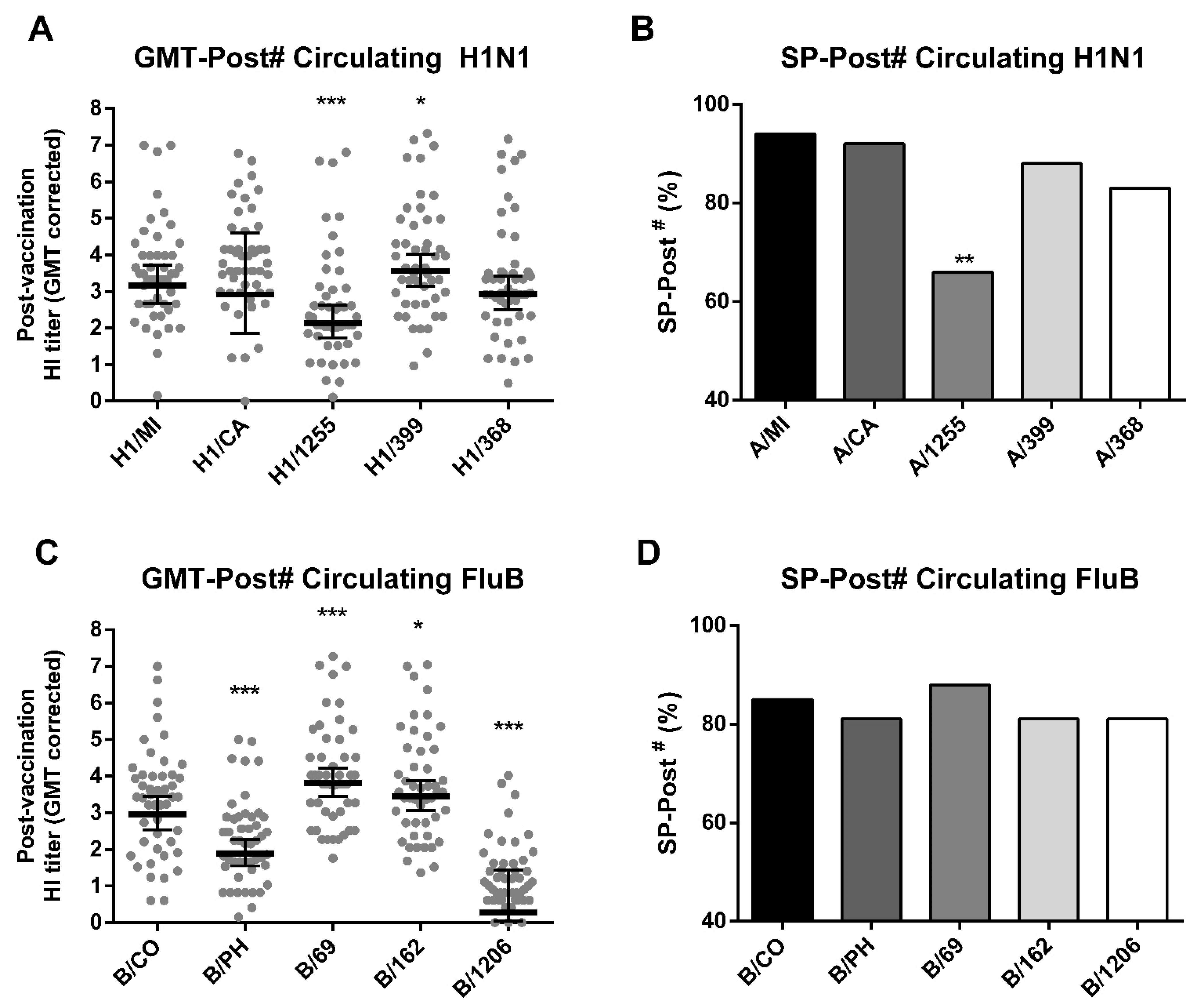
3.3. TIV Enhances HI Titers and Seroprotection for Matched and Other Contemporary Circulating Strains
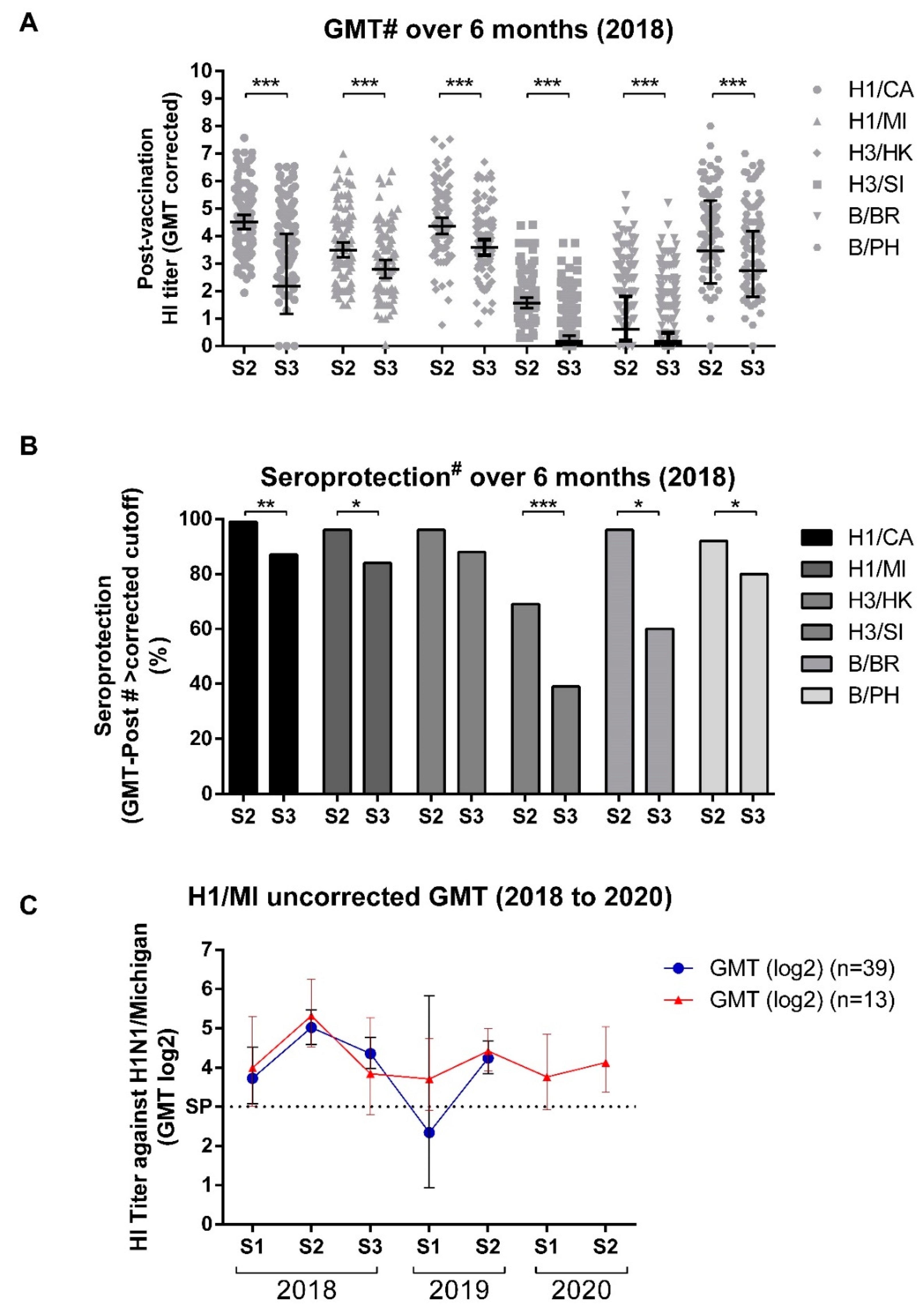
3.4. GMT Titers Wane over Time but Remain High through Most Influenza Seasons
3.5. Vaccine Efficacy Appears to Be High, but Does Not Exclude Infection
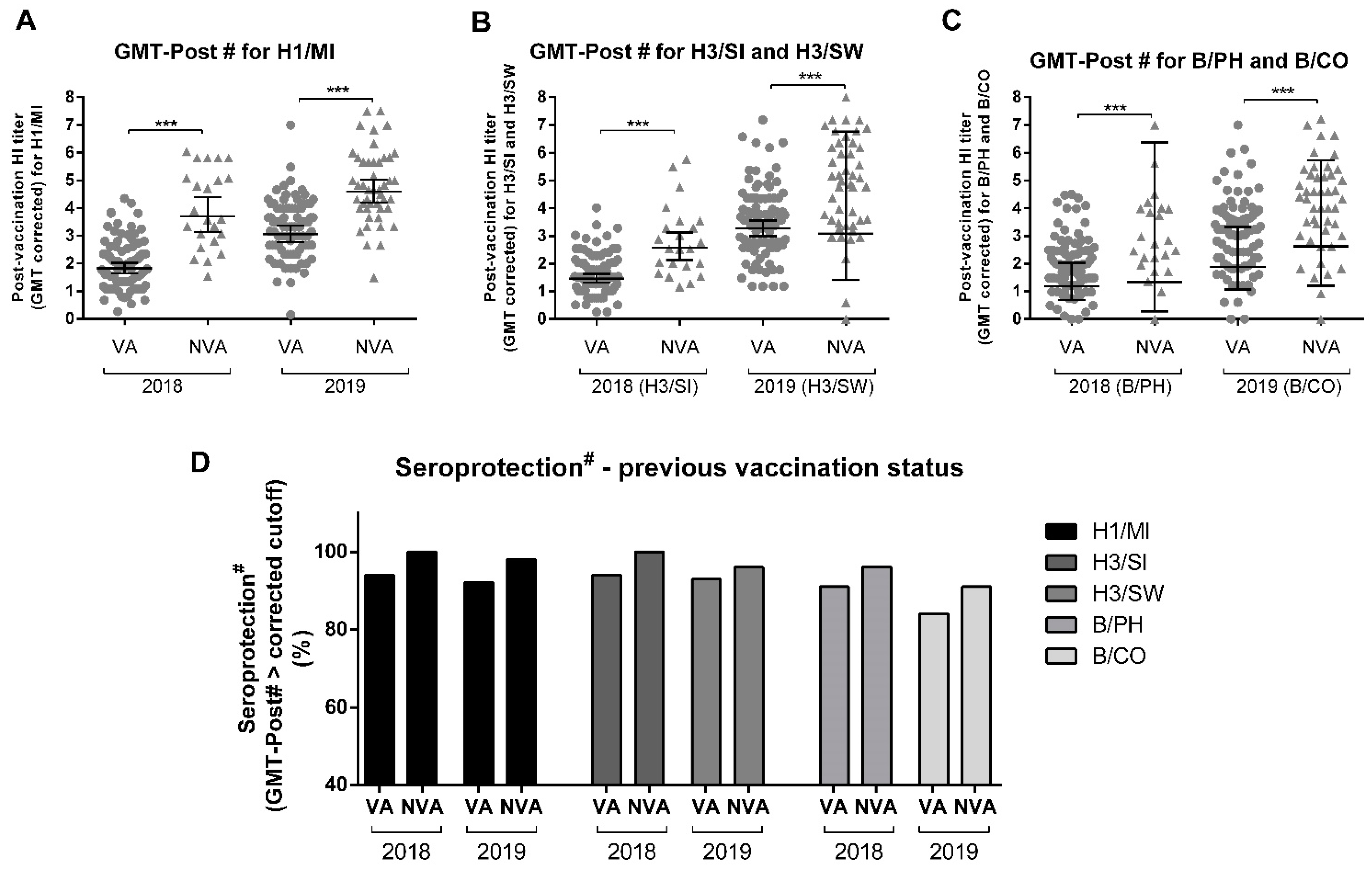
3.6. Previous Influenza Vaccine Influences Induction of Anti-HA Antibodies
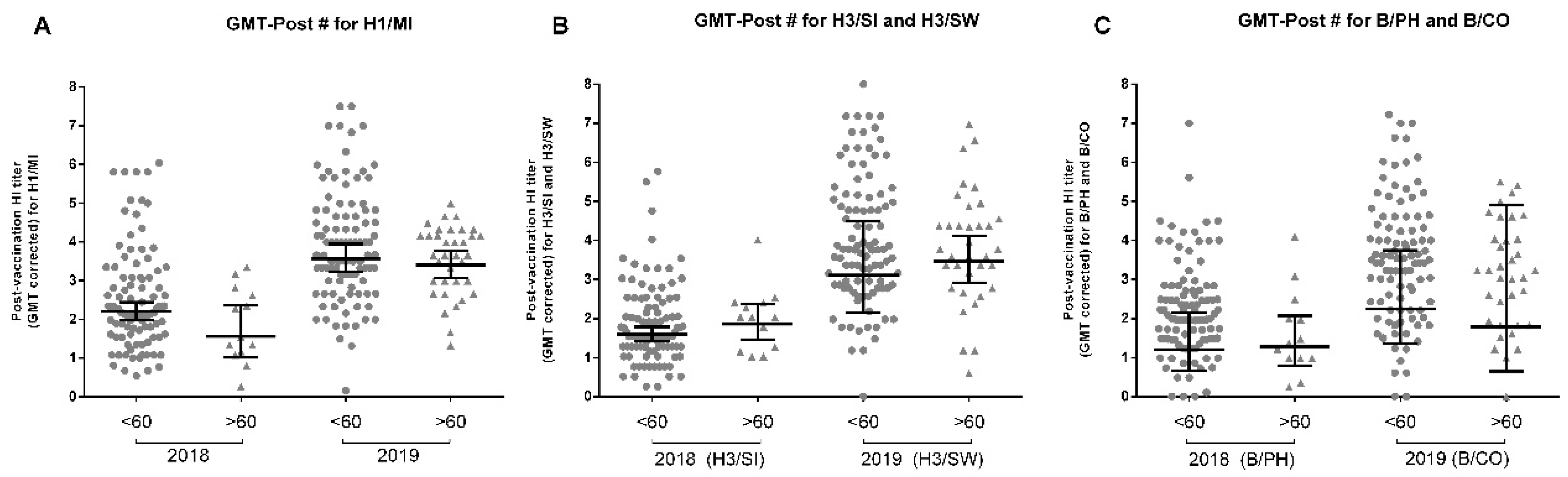
3.7. Age Influences Influenza Vaccine Response
3.8. Other Population Characteristics and Comorbidities Did Not Seem to Impact Vaccine Response
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- CDC. Disease Burden of Influenza. 2020. Available online: https://www.cdc.gov/flu/about/burden/index.html (accessed on 11 December 2019).
- Petrova, V.N.; Russell, C.A. The evolution of seasonal influenza viruses. Nat. Rev. Microbiol. 2017, 16, 47–60. [Google Scholar] [CrossRef]
- Haq, K.; McElhaney, J.E. Immunosenescence: Influenza vaccination and the elderly. Curr. Opin. Immunol. 2014, 29, 38–42. [Google Scholar] [CrossRef] [PubMed]
- Iuliano, A.D.; Roguski, K.M.; Chang, H.H.; Muscatello, D.J.; Palekar, R.; Tempia, S.; Cohen, C.; Gran, J.M.; Schanzer, D.; Cowling, B.J.; et al. Estimates of global seasonal influenza-associated respiratory mortality: A modelling study. Lancet 2018, 391, 1285–1300. [Google Scholar] [CrossRef]
- Belongia, E.A.; Skowronski, D.M.; McLean, H.Q.; Chambers, C.; Sundaram, M.E.; De Serres, G. Repeated annual influenza vaccination and vaccine effectiveness: Review of evidence. Expert Rev. Vaccines 2017, 16, 723–736. [Google Scholar] [CrossRef] [PubMed]
- Sheridan, P.A.; Paich, H.A.; Handy, J.; Karlsson, E.A.; Hudgens, M.G.; Sammon, A.B.; Holland, L.A.; Weir, S.; Noah, T.L.; Beck, M.A. Obesity is associated with impaired immune response to influenza vaccination in humans. Int. J. Obes. 2012, 36, 1072–1077. [Google Scholar] [CrossRef]
- Skowronski, D.M.; Chambers, C.; Sabaiduc, S.; De Serres, G.; Winter, A.L.; Dickinson, J.A.; Gubbay, J.B.; Drews, S.J.; Martineau, C.; Charest, H.; et al. Beyond Antigenic Match: Possible Agent-Host and Immuno-epidemiological Influences on Influenza Vaccine Effectiveness During the 2015–2016 Season in Canada. J. Infect. Dis. 2017, 216, 1487–1500. [Google Scholar] [CrossRef] [PubMed]
- Sacadura-Leite, E.; Sousa-Uva, A.; Rebelo-de-Andrade, H. Antibody response to the influenza vaccine in healthcare workers. Vaccine 2012, 30, 436–441. [Google Scholar] [CrossRef] [PubMed]
- Ray, G.T.; Lewis, N.; Klein, N.P.; Daley, M.F.; Wang, S.V.; Kulldorff, M.; Fireman, B. Intraseason Waning of Influenza Vaccine Effectiveness. Clin. Infect. Dis. 2019, 68, 1623–1630. [Google Scholar] [CrossRef]
- Ferdinands, J.M.; Fry, A.M.; Reynolds, S.; Petrie, J.G.; Flannery, B.; Jackson, M.L.; Belongia, E.A. Intraseason Waning of Influenza Vaccine Protection: Evidence From the US Influenza Vaccine Effectiveness. Clin. Infect. Dis. 2017, 64, 544–550. [Google Scholar] [CrossRef]
- Perdigão, A.C.B.; Siqueira, M.M.; Moura, F.E.A. Seasonality of Influenza in the Tropics: A Distinct Pattern in Northeastern Brazil. Am. J. Trop. Med. Hyg. 2009, 81, 180–183. [Google Scholar] [CrossRef]
- Caini, S.; Andrade, W.; Badur, S.; Balmaseda, A.; Barakat, A.; Bella, A.; Bimohuen, A.; Brammer, L.; Bresee, J.; Bruno, A.; et al. Temporal Patterns of Influenza A and B in Tropical and Temperate Countries: What Are the Lessons for Influenza Vaccination? PLoS ONE 2016, 11, e0152310. [Google Scholar] [CrossRef]
- Huang, K.Y.A.; Chang, S.C.; Huang, Y.C.; Chiu, C.H.; Lin, T.Y. Antibody Responses to Trivalent Inactivated Influenza Vaccine in Health Care Personnel Previously Vaccinated and Vaccinated for The First Time. Sci. Rep. 2017, 7, 40027. [Google Scholar] [CrossRef] [PubMed]
- Castrucci, M.R. Factors affecting immune responses to the influenza vaccine. Hum. Vaccines Immunother. 2017, 14, 637–646. [Google Scholar] [CrossRef]
- Coudeville, L.; Bailleux, F.; Riche, B.; Megas, F.; Andre, P.; Ecochard, R. Relationship between haemagglutination-inhibiting antibody titres and clinical protection against influenza: Development and application of a bayesian random-effects model. BMC Med. Res. Methodol. 2010, 10, 18. [Google Scholar] [CrossRef] [PubMed]
- CDC; WHO. CDC Protocol of Real-Time RT-PCR for Influenza A (H1N1). 2009. Available online: http://www.who.int/csr/resources/publications/swineflu/realtimeptpcr/en/ (accessed on 3 August 2015).
- Beyer, W.E.P.; Palache, A.M.; Lüchters, G.; Nauta, J.; Osterhaus, A.D.M.E. Seroprotection rate, mean fold increase, seroconversion rate: Which parameter adequately expresses seroresponse to influenza vaccination? Virus Res. 2004, 103, 125–132. [Google Scholar] [CrossRef] [PubMed]
- CDC. Defining Adult Overweight and Obesity. 2012; pp. 8–9. Available online: https://www.cdc.gov/obesity/adult/defining.html (accessed on 18 February 2020).
- Gomez Lorenzo, M.M.; Fenton, M.J. Immunobiology of Influenza Vaccines. Chest 2013, 143, 502–510. [Google Scholar] [CrossRef] [PubMed]
- Pera, A.; Campos, C.; López, N.; Hassouneh, F.; Alonso, C.; Tarazona, R.; Solana, R. Immunosenescence: Implications for response to infection and vaccination in older people. Maturitas 2015, 82, 50–55. [Google Scholar] [CrossRef]
- Louie, J.K.; Acosta, M.; Samuel, M.C.; Schechter, R.; Vugia, D.J.; Harriman, K.; Matyas, B.T. A Novel Risk Factor for a Novel Virus: Obesity and 2009 Pandemic Influenza A (H1N1). Clin. Infect. Dis. 2011, 52, 301–312. [Google Scholar] [CrossRef] [PubMed]
- Neidich, S.D.; Green, W.D.; Rebeles, J.; Karlsson, E.A.; Schultz-Cherry, S.; Noah, T.L.; Chakladar, S.; Hudgens, M.G.; Weir, S.S.; Beck, M.A. Increased risk of influenza among vaccinated adults who are obese. Int. J. Obes. 2017, 41, 1324–1330. [Google Scholar] [CrossRef]
- Xie, H.; Jing, X.; Li, X.; Lin, Z.; Plant, E.; Zoueva, O.; Yang, H.; Ye, Z. Immunogenicity and cross-reactivity of 2009–2010 inactivated seasonal influenza vaccine in US adults and elderly. PLoS ONE 2011, 6, e16650. [Google Scholar] [CrossRef] [PubMed]
- Mondini, G.; Braga, P.E.; Lopes, M.H.; Sartori, A.M.C.; Miyaji, K.T.; Infante, V.; Randi, B.A.; Timenetsky, M.D.C.S.T.; Ferreira, J.C.D.O.A.; Sakita, N.K.; et al. Prospective cohort studies to evaluate the safety and immunogenicity of the 2013, 2014, and 2015 seasonal influenza vaccines produced by Instituto Butantan. Rev. Do Inst. De Med. Trop. De São Paulo 2018, 60, e37. [Google Scholar] [CrossRef] [PubMed]
- Kieninger, D.; Sheldon, E.; Lin, W.Y.Y.; Yu, C.J.J.; Bayas, J.M.; Gabor, J.J.; Esen, M.; Fernandez Roure, J.L.; Narejos Perez, S.; Alvarez Sanchez, C.; et al. Immunogenicity, reactogenicity and safety of an inactivated quadrivalent influenza vaccine candidate versus inactivated trivalent influenza vaccine: A phase III, randomized trial in adults aged ≥18 years. BMC Infect. Dis. 2013, 13, 343. [Google Scholar] [CrossRef] [PubMed]
- Nuñez, I.A.; Carlock, M.A.; Allen, J.D.; Owino, S.O.; Moehling, K.K.; Nowalk, P.; Susick, M.; Diagle, K.; Sweeney, K.; Mundle, S.; et al. Impact of age and pre-existing influenza immune responses in humans receiving split inactivated influenza vaccine on the induction of the breadth of antibodies to influenza A strains. PLoS ONE 2017, 12, e0185666. [Google Scholar] [CrossRef] [PubMed]
- Young, B.; Zhao, X.; Cook, A.R.; Parry, C.M.; Wilder-Smith, A.; I-Cheng, M.C. Do antibody responses to the influenza vaccine persist year-round in the elderly? A systematic review and meta-analysis. Vaccine 2017, 35, 212–221. [Google Scholar] [CrossRef] [PubMed]
- WHO. Recommended Composition of Influenza Virus Vaccines for Use in the 2019 Southern Hemisphere Influenza Season. 2018. Available online: https://www.who.int/influenza/vaccines/virus/recommendations/201809_recommendation.pdf (accessed on 13 April 2021).
- WHO. Recommended Composition of Influenza Virus Vaccines for Use in the 2020 Southern Hemisphere Influenza Season. 2020. Available online: http://www.who.int/influenza/vaccines/virus/recommendations/2020_south/en/ (accessed on 27 March 2021).
- Belshe, R.B. The need for quadrivalent vaccine against seasonal influenza. Vaccine 2010, 28 (Suppl. 4), D45–D53. [Google Scholar] [CrossRef] [PubMed]
- Tricco, A.C.; Chit, A.; Soobiah, C.; Hallett, D.; Meier, G.; Chen, M.H.; Tashkandi, M.; Bauch, C.T.; Loeb, M. Comparing influenza vaccine efficacy against mismatched and matched strains: A systematic review and meta-analysis. BMC Med. 2013, 11, 1. [Google Scholar] [CrossRef] [PubMed]
- Skowronski, D.M.; Chambers, C.; De Serres, G.; Dickinson, J.A.; Winter, A.L.; Hickman, R.; Chan, T.; Jassem, A.N.; Drews, S.J.; Charest, H.; et al. Early season co-circulation of influenza A(H3N2) and B(Yamagata): Interim estimates of 2017/18 vaccine effectiveness, Canada, January 2018. Eurosurveillance 2018, 23, 18–00035. [Google Scholar] [CrossRef]
- Henry, C.; Palm, A.K.E.; Krammer, F.; Wilson, P.C. From Original Antigenic Sin to the Universal Influenza Virus Vaccine. Trends Immunol. 2018, 39, 70–79. [Google Scholar] [CrossRef]
- Zarnitsyna, V.I.; Ellebedy, A.H.; Davis, C.; Jacob, J.; Ahmed, R.; Antia, R. Masking of antigenic epitopes by antibodies shapes the humoral immune response to influenza. Philos. Trans. R. Soc. B Biol. Sci. 2015, 370, 20140248. [Google Scholar] [CrossRef]
- Cobey, S.; Hensley, S.E. Immune history and influenza virus susceptibility. Curr. Opin. Virol. 2017, 22, 105–111. [Google Scholar] [CrossRef]
- Linderman, S.L.; Hensley, S.E. Antibodies with ‘Original Antigenic Sin’ Properties Are Valuable Components of Secondary Immune Responses to Influenza Viruses. PLoS Pathog. 2016, 12, e1005806. [Google Scholar] [CrossRef] [PubMed]
- Park, M.S.; Kim, J.I.; Park, S.; Lee, I.; Park, M.S.M.S. Original Antigenic Sin Response to RNA Viruses and Antiviral Immunity. Immune Netw. 2016, 16, 261–270. [Google Scholar] [CrossRef] [PubMed]
- Zarnitsyna, V.I.; Lavine, J.; Ellebedy, A.; Ahmed, R.; Antia, R. Multi-epitope Models Explain How Pre-existing Antibodies Affect the Generation of Broadly Protective Responses to Influenza. PLoS Pathog. 2016, 12, e1005692. [Google Scholar] [CrossRef] [PubMed]
- Liu, F.; Gross, F.L.; Jefferson, S.N.; Holiday, C.; Bai, Y.; Wang, L.; Zhou, B.; Levine, M.Z. Age-specific effects of vaccine egg-adaptation and immune priming on A(H3N2) antibody responses following influenza vaccination. J. Clin. Investig. 2021, 131, e146138. [Google Scholar] [CrossRef] [PubMed]
- Kawakami, C.; Yamayoshi, S.; Akimoto, M.; Nakamura, K.; Miura, H.; Fujisaki, S.; Pattinson, D.J.; Shimizu, K.; Ozawa, H.; Momoki, T.; et al. Genetic and antigenic characterisation of influenza A(H3N2) viruses isolated in Yokohama during the 2016/17 and 2017/18 influenza seasons. Eurosurveillance 2019, 24, 1800467. [Google Scholar] [CrossRef]
- Kang, H.J.; Chu, K.B.; Yoon, K.W.; Eom, G.D.; Mao, J.; Dimitrov, S.; Quan, F.S. Cross-Protection Induced by Virus-like Particles Derived from the Influenza B Virus. Biomedicines 2022, 10, 1618. [Google Scholar] [CrossRef]
- Höpping, A.M.; Mcelhaney, J.; Fonville, J.M.; Powers, D.C.; Beyer, W.E.P.; Smith, D.J. The confounded effects of age and exposure history in response to influenza vaccination. Vaccine 2016, 34, 540–546. [Google Scholar] [CrossRef]
- Haralambieva, I.H.; Painter, S.D.; Kennedy, R.B.; Ovsyannikova, I.G.; Lambert, N.D.; Goergen, K.M.; Oberg, A.L.; Poland, G.A. The impact of immunosenescence on humoral immune response variation after influenza A/H1N1 vaccination in older subjects. PLoS ONE 2015, 10, e0122282. [Google Scholar] [CrossRef]
- Reber, A.J.; Chirkova, T.; Kim, J.H.; Cao, W.; Biber, R.; Shay, D.K.; Sambhara, S. Immunosenescence and Challenges of Vaccination against Influenza in the Aging Population. Aging Dis. 2012, 3, 68–90. [Google Scholar]
- Dugan, H.L.; Henry, C.; Wilson, P.C. Aging and influenza vaccine-induced immunity. Cell. Immunol. 2020, 348, 103998. [Google Scholar] [CrossRef]
- McElhaney, J.E.; Xie, D.; Hager, W.D.; Barry, M.B.; Wang, Y.; Kleppinger, A.; Ewen, C.; Kane, K.P.; Bleackley, R.C. T cell responses are better correlates of vaccine protection in the elderly. J. Immunol. 2006, 176, 6333–6339. [Google Scholar] [CrossRef] [PubMed]
- Wagar, L.E.; Gentleman, B.; Pircher, H. Influenza-Specific T Cells from Older People Are Enriched in the Late Effector Subset and Their Presence Inversely Correlates with Vaccine Response. PLoS ONE 2011, 6, e23698. [Google Scholar] [CrossRef] [PubMed]
- Thompson, W.W.; Shay, D.K.; Weintraub, E.; Cox, N.; Anderson, L.J.; Fukuda, K. Mortality associated with influenza and respiratory syncytial virus in the United States. JAMA 2003, 289, 179–186. [Google Scholar] [CrossRef] [PubMed]
- Ferreira, A.P.D.S.; Szwarcwald, C.L.; Damacena, G.N. Prevalência e fatores associados da obesidade na população brasileira: Estudo com dados aferidos da Pesquisa Nacional de Saúde, 2013. Rev. Bras. De Epidemiol. 2019, 22, e190024. [Google Scholar] [CrossRef] [PubMed]
- WHO. Obesity and Overweight. Available online: https://www.who.int/news-room/fact-sheets/detail/obesity-and-overweight (accessed on 15 February 2021).
- Green, W.D.; Beck, M.A. Obesity Impairs the Adaptive Immune Response to Influenza Virus. Ann. Am. Thorac. Soc. 2017, 14 (Suppl. 5), S406–S409. [Google Scholar] [CrossRef]
- Park, H.L.; Shim, S.H.; Lee, E.Y.; Cho, W.; Park, S.; Jeon, H.J.; Ahn, S.Y.; Kim, H.; Nam, J.H. Obesity-induced chronic inflammation is associated with the reduced efficacy of influenza vaccine. Hum. Vaccines Immunother. 2014, 10, 1181–1186. [Google Scholar] [CrossRef]
- Saltiel, A.R.; Olefsky, J.M. Inflammatory mechanisms linking obesity and metabolic disease. J. Clin. Investig. 2017, 127, 1–4. [Google Scholar] [CrossRef]
- Frasca, D.; Ferracci, F.; Diaz, A.; Romero, M.; Lechner, S.; Blomberg, B.B. Obesity decreases B cell responses in young and elderly individuals. Obesity 2016, 24, 615–625. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| H1N1 | H3N2 | Influenza B | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| H1/CA | H1/MI | H1/BRI | H3/HK | H3/SI | H3/SW | H3/SA | B/BR | B/PH | B/CO | B/WA | ||
| 2018 | GMT-Pre (Overall) (Log2) | 4.8 (1.8) | 3.9 (1.7) | 4.8 (1.8) | 4.2 (1.7) | 3.9 (1.4) | 3.8 (2.2) | |||||
| GMT-Post (Overall) (Log2) | 5.9 (1.8) | 5.2 (1.8) | 6.2 (1.4) | 4.9 (1.6) | 4.5 (1.6) | 5.0 (2.0) | ||||||
| GMT-Pre (Pre<3) (Log2) | 1.7 (0.7) | 1.6 (0.8) | 1.2 (1.1) | 1.5 (0.8) | 1.3 (0.8) | 1.3 (0.9) | ||||||
| GMT-Post (Pre<3) (Log2) | 3.6 (1.5) | 3.3 (1.5) | 6.3 (2.1) | 3.1 (1.4) | 2.5 (1.1) | 3.2 (1.8) | ||||||
| SC (fold-increase >4) (%) | 26 | 35 | 26 | 17 | 10 ** | 25 ** | ||||||
| SP-Pre (GMT>3) (%) | 87 | 75 | 92 | 82 | 75 | 67 | ||||||
| SP-Post (GMT>3) (%) | 97 | 92 | 99 | 92 | 83 | 85 | ||||||
| GMT-Post # (corrected) (Log2) | 2.3 (1.1) | 2.4 (1.2) | 4.2 (1.3) *** | 1.9 (0.9) *** | 1.3 (0.8) *** | 2.2 (1.2) *** | ||||||
| SP-Post # (GMT-Post#>corrected cutoff) (%) | 98 | 96 | 99 | 95 | 91 | 92 | ||||||
| 2019 | GMT-Pre (Overall) (Log2) | 4.2 (1.6) | 3.6 (1.5) | 3.2 (1.5) | 3.6 (1.3) | 3.4 (1.7) | 2.4 (1.9) | |||||
| GMT-Post (Overall) (Log2) | 5.7 (1.6) | 5.0 (1.4) | 4.4 (1.9) | 5.4 (1.7) | 4.3 (1.7) | 4.3 (1.7) | ||||||
| GMT-Pre (Pre<3) (Log2) | 1.7 (0.8) | 1.7 (0.7) | 1.6 (0.7) | 1.9 (0.6) | 1.8 (0.7) | 1.1 (0.8) | ||||||
| GMT-Post (Pre<3) (Log2) | 4.3 (2.3) | 4.3 (1.7) | 3.1 (1.9) | 4.9 (2.4) | 3.4 (1.6) | 3.8 (1.7) | ||||||
| SC (fold-increase >4) (%) | 36 | 32 | 25 ** | 41 ** | 21 *** | 46 *** | ||||||
| SP-Pre (GMT>3) (%) | 83 | 77 | 64 | 77 | 60 | 42 | ||||||
| SP-Post (GMT>3) (%) | 95 | 95 | 83 | 94 | 80 | 84 | ||||||
| GMT-Post # (corrected) (Log2) | 3.9 (1.5) | 3.8 (1.3) | 2.1 (1.5) *** | 3.9 (1.6) *** | 2.3 (1.4) *** | 3.4 (1.5) *** | ||||||
| SP-Post # (GMT-Post#>corrected cutoff) (%) | 95 | 94 | 86 *** | 94 *** | 83 | 86 | ||||||
| 2020 | GMT-Pre (Overall) (Log2) | 4.5 (1.8) | 4.7 (2.0) | 4.5 (1.1) | 4.8 (1.1) | 4.3 (1.5) | 3.9 (1.4) | |||||
| GMT-Post (Overall) (Log2) | 5.1 (1.7) | 5.7 (1.5) | 5.2 (1.0) | 5.7 (1.1) | 4.7 (1.4) | 4.5 (1.1) | ||||||
| GMT-Pre (Pre<3) (Log2) | 1.5 (1.0) | 1.1 (1.3) | - | - | 1.7 (0.6) | 1.8 (0.9) | ||||||
| GMT-Post (Pre<3) (Log2) | 3.0 (0.0) | 4.5 (1.0) | - | - | 3.7 (1.5) | 3.4 (1.0) | ||||||
| SC (fold-increase >4) (%) | 10 | 19 | 10 | 26 | 13 | 13 | ||||||
| SP-Pre (GMT>3) (%) | 87 | 87 | 100 | 100 | 90 | 80 | ||||||
| SP-Post (GMT>3) (%) | 100 | 100 | 100 | 100 | 97 | 97 | ||||||
| GMT-Post # (corrected) (Log2) | 2.9 (1.1) | 3.2 (1.0) | 1.9 (0.6) *** | 2.9 (0.9) *** | 2.8 (0.9) *** | 2.2 (0.7) *** | ||||||
| SP-Post # (GMT-Post#>corrected cutoff) (%) | 100 | 100 | 100 | 100 | 90 | 90 | ||||||
| A/Michigan/45/2015 (H1N1) | A/Switzerland/8060/2017 (H3N2) | B/Colorado/06/2017 (Victoria) | |
|---|---|---|---|
| VA | 9 (11%) | 23 (27%) | 26 (30%) |
| NVA | 33 (72%) | 31 (67%) | 35 (76%) |
| Odds-Ratio (CI) | 0.05 (0.02–0.12) *** | 0.18 (0.08–0.39) *** | 0.14 (0.06–0.31) *** |
| <60 | 35 (36%) | 38 (39%) | 48 (49%) |
| >60 | 7 (20%) | 16 (46%) | 13 (37%) |
| Odds-Ratio (CI) | 2.26 (0.89–5.70) | 0.89 (0.41–1.90) | 1.66 (0.75–3.66) |
| EG | 33 (34%) | 47 (19%) | 38 (39%) |
| OB | 6 (27%) | 10 (46%) | 7 (32%) |
| Odds-Ratio (CI) | 0.61 (0.22–1.68) | 0.89 (0.35–2.25) | 0.73 (0.27–1.94) |
| H1N1 | H3N2 | Influenza B | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| H1/CA | H1/MI | H3/HK | H3/SI | H3/SW | B/BR | B/PH | B/CO | |||
| 2018 | <60 | GMT-Post* (corrected) (Log2) | 2.4 (1.5) | 2.5 (1.3) | 4.2 (1.3) | 1.8 (1.0) | 1.3 (0.8) | 2.2 (1.2) | ||
| SP-Post (GMT-Post*>corrected cutoff) (%) | 98 | 96 | 99 | 94 | 90 | 93 | ||||
| >60 | GMT-Post* (corrected) (Log2) | 1.9 (0.9) | 1.9 (0.9) | 4.3 (1.3) | 2.0 (0.8) | 1.5 (0.7) | 1.6 (1.1) | |||
| SP-Post (GMT-Post*>corrected cutoff) (%) | 100 | 92 | 100 | 100 | 100 | 85 | ||||
| 2019 | <60 | GMT-Post* (corrected) (Log2) | 4.1 (1.6) | 3.9 (1.5) | 2.2 (1.5) | 4.0 (1.6) | 2.5 (1.4) | 3.5 (1.6) | ||
| SP-Post (GMT-Post*>corrected cutoff) (%) | 94 | 91 | 89 | 95 | 85 | 88 | ||||
| >60 | GMT-Post* (corrected) (Log2) | 3.5 (0.9) | 3.5 (0.9) | 1.6 (1.3) | 3.8 (1.4) | 1.9 (1.1) | 3.1 (1.4) | |||
| SP-Post (GMT-Post*>corrected cutoff) (%) | 97 | 94 | 77 | 91 | 77 | 83 | ||||
| 2019 | EG | GMT-Post* (corrected) (Log2) | 3.9 (1.6) | 3.9 (1.4) | 2.1 (1.5) | 3.9 (1.6) | 2.4 (1.4) | 3.5 (1.5) | ||
| SP-Post (GMT-Post*>corrected cutoff) (%) | 93 | 93 | 87 | 95 | 81 | 90 | ||||
| OB | GMT-Post* (corrected) (Log2) | 4.0 (1.4) | 3.8 (1.2) | 1.9 (1.5) | 4.1 (1.6) | 1.8 (1.1) | 2.9 (1.6) | |||
| SP-Post (GMT-Post*>corrected cutoff) (%) | 100 | 95 | 82 | 86 | 82 | 73 | ||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Capão, A.; Aguiar-Oliveira, M.L.; Caetano, B.C.; Neves, T.K.; Resende, P.C.; Almeida, W.A.F.; Miranda, M.D.; Martins-Filho, O.A.; Brown, D.; Siqueira, M.M.; et al. Analysis of Viral and Host Factors on Immunogenicity of 2018, 2019, and 2020 Southern Hemisphere Seasonal Trivalent Inactivated Influenza Vaccine in Adults in Brazil. Viruses 2022, 14, 1692. https://doi.org/10.3390/v14081692
Capão A, Aguiar-Oliveira ML, Caetano BC, Neves TK, Resende PC, Almeida WAF, Miranda MD, Martins-Filho OA, Brown D, Siqueira MM, et al. Analysis of Viral and Host Factors on Immunogenicity of 2018, 2019, and 2020 Southern Hemisphere Seasonal Trivalent Inactivated Influenza Vaccine in Adults in Brazil. Viruses. 2022; 14(8):1692. https://doi.org/10.3390/v14081692
Chicago/Turabian StyleCapão, Artur, Maria L. Aguiar-Oliveira, Braulia C. Caetano, Thayssa K. Neves, Paola C. Resende, Walquiria A. F. Almeida, Milene D. Miranda, Olindo A. Martins-Filho, David Brown, Marilda M. Siqueira, and et al. 2022. "Analysis of Viral and Host Factors on Immunogenicity of 2018, 2019, and 2020 Southern Hemisphere Seasonal Trivalent Inactivated Influenza Vaccine in Adults in Brazil" Viruses 14, no. 8: 1692. https://doi.org/10.3390/v14081692