1. Introduction
Human betaherpesvirus 5, also called human cytomegalovirus (HCMV), is a ubiquitous pathogen that rarely causes symptoms in healthy individuals but can lead to severe or even life-threatening diseases in immunocompromised patients and newborns (reviewed in [
1]). Characteristic of betaherpesviruses, HCMV has a slow replication cycle and shows a high level of host specificity since it cannot productively replicate in cells of other species [
2].
During lytic HCMV replication, viral proteins are expressed in three distinct phases termed immediate early (IE), early (E), and late (L) phase [
3]. The immediate early 1 (IE1) protein, expressed very early after infection, is important for initiating the lytic replication cycle, especially under low-multiplicity conditions [
4]. HCMV IE1 (hIE1, 491 amino acids (aa) is a multifunctional protein. Its structure can be subdivided into a short N-terminal domain harbouring a nuclear localisation sequence (NLS, aa 1-24), a core domain (aa 24-382), an acidic region (aa 373-475), and a chromatin tethering domain (CTD, aa 467-491) [
5,
6,
7,
8]. The NLS domain is required for nuclear transport. It has previously been demonstrated that the large IE1 core is essential to antagonizing promyelocytic leukemia nuclear bodies (PML-NBs), which are part of the intrinsic immune system [
7]. These multiprotein complexes appear as dot-like structures of the cell nucleus and can induce the silencing of viral gene expression. PML-NBs are formed by the main organiser promyelocytic leukemia (PML) protein and further restriction factors like ATRX, Sp100, and hDaxx [
9,
10,
11]. IE1 has been shown to inhibit the de novo SUMOylation of PML, followed by PML-NB disruption and inactivation of the intrinsic immune barrier [
12,
13].
Scherer et al. solved the structure of the rhesus CMV IE1 core, which revealed an unusual, all alpha-helical, femur-like fold [
7]. This structure of IE1 is conserved in different species: the core domains of rat CMV IE1 (rIE1) and rhesus CMV IE1 (rhesIE1) show high structural similarity to human IE1 (hIE1) core apart from small differences. All proteins contain a stalk between a C- and N-terminal head region. While the hIE1 core is built by eleven helices, the rIE1 core (aa 30-392) is formed by twelve. Furthermore, helix 2 of the rIE1 core is displaced, and helix 1 is shorter than in the hIE1 core. Interestingly, the folding is conserved, although the sequence identity between primate and rodent IE1 is very low [
14]. The core domains of hIE1 and rIE1 bind the natural host PML-NBs via coiled-coil interactions. Despite the structural similarity, this interaction and deSUMOylation and disruption of PML-NBs are species-dependent activities and do not occur during cross-species infection. These findings show that PML NBs contribute to a species barrier against CMVs [
7,
14].
In addition to PML antagonization, hIE1 plays a major role in the transactivation of viral promoters, as it enhances the transactivation capacity of IE2 [
15,
16]. It was shown that a possible transactivation site of IE1 is located in the core domain [
16]. While Sandford et al. observed that rat CMV IE2 (rIE2) can transactivate heterologous promotors, the role of rIE1 in transactivation has not been demonstrated yet [
17]. Furthermore, hIE1 also influences other cellular mechanisms in a pro-viral manner. Schilling et al. detected that the hIE1 core interacts with the flap endonuclease one protein (FEN1) that plays a role in the DNA damage response pathway. Due to this interaction, hIE1 stabilizes FEN1 and has a pro-viral effect on HCMV lytic replication [
18].
The third structural part of hIE1 is formed by an acidic domain (aa 373-475), a highly unordered region [
19]. Various groups showed that the SUMOylation site K450 and the interaction site for the signal transducer and activator of transcription (STAT2) are located within this domain [
20,
21,
22]. Paulus et al. revealed that IE1 antagonizes the interferon response by intranuclear relocalisation of STAT2. This results in a reduction of free STAT2 to form the transcription complex IFN stimulated gene factor 3 (ISGF3) together with STAT1 and IFN-regulatory factor 9 (IFN9). Consequently, the reduction of ISGF3 leads to impaired expression of interferon-stimulated genes (ISGs) [
23,
24]. Interestingly, rat IE1 also contains a highly unordered C-terminal region that could potentially harbour a STAT2 binding domain [
14]. Finally, the so-called chromatin tethering domain (CTD) is located at the very C-terminus of hIE1 (aa 476-491). This domain mediates IE1 co-localisation with condensed chromatin and interacts with histones in the chromatin structure [
25,
26]. Different groups showed that the CTD is not essential for CMV lytic replication. Muecke et al. speculated that the CTD probably plays a role during latency [
8,
26]. In particular, Tarrant-Elorza et al. reported that a smaller IE1 protein species (IE1x4) is expressed in hematopoietic progenitor cells that are latently infected with HCMV, and this protein appeared to be required for viral genome maintenance [
27]. The sequence of the CTD is conserved in primate species but cannot be detected in rat IE1 [
8,
17,
26].
In this study, we investigated whether the similarities in structure between hIE1 and rIE1 can be utilised to characterize the interaction surface of PML. Furthermore, we performed a comparative analysis to identify either species-specific or conserved functions of primate and rodent cytomegalovirus IE1 proteins. Our results demonstrate that while PML binding occurs in a highly species-specific manner, IE1 proteins possess a conserved function in cross-species antagonization of interferon signaling via relocalisation of STAT2.
2. Materials and Methods
2.1. Oligonucleotide, Plasmids, and Cloning
All oligonucleotide primers used in this study were purchased from Biomers GmbH (Ulm, Germany). All primers and cloned expression plasmids are listed in
Table S1. The hIE1_rH mutants were generated using Gibson assembly 1 Step Kit (Synthetic Genomics, La Jolla, CA, USA). Flag-tagged hIE1, hIE1 1-382, and rIE1 expression plasmids [
7,
14] were used as templates for the polymerase chain reaction (PCR). The helices of rIE1 were generated via PCR using Flag-tagged rIE1 as a template. As the backbone for cloning, Flag hIE1 1-382 was used. Mutants hIE1_rH1, hIE1_rH1/2; hIE1_rH4 and hIE1_rH8 were generated by cloning of one insert via PCR linearised vector. Mutants hIE1_rH2 and hIE1_rH10/11 were generated by cloning three inserts into the linearised Flag pcDNA3 vector [
28]. Full-length versions of the hIE1_rH mutants were generated by cleaving the expression plasmids using the fast digest (FD) restriction enzyme XhoI (Thermo Fisher, Waltham, MA, USA) and adding the C-terminus (aa 383-491) of Flag hIE1 via Gibson Assembly. Furthermore, the NanoBRET expression plasmids were purchased from Promega (#N1811). The plasmids were cleaved with the FD enzymes SacI and EcoRI (Thermo Fisher, Waltham, MA, USA). The cloning was also done via Gibson assembly. The hIE1 L174P mutant from Scherer et al. [
29], Myc PML [
30], and Flag rPML [
14] were also inserted in the expression plasmid for the NanoBRET assay. The mutant hIE1 cc172-176 published by Paulus, Harwardt, et al. [
31] was also generated. For this, a fragment of hIE1 that carries the cc172-176 mutation was purchased from Integrated DNA Technologies (IDT, Coralville, IA, USA). Then, the segment was integrated via Gibson assembly into Flag IE1 that was cleaved with FD EcoRV (Thermo Fisher, Waltham, MA, USA). Furthermore, besides expression plasmids with Flag-tagged hIE1, hIE1 core, rIE1, and rIE1 core [
14], an expression plasmid with a firefly luciferase gene under the control of the UL112 promotor (pHM 142, [
32]) and an expression plasmid for IE2 (pHM 134, [
33]) were used for the gaussia luciferase assay. Additionally, Schilling et al. generated the Flag-tagged FEN1 176-380 [
18] used for cloning the expression plasmids for the NanoBRET assay and the stabilization assay.
2.2. Cells and Transfection
HEK293T cells were cultivated in Dulbecco’s minimal essential medium (DMEM) (Gibco, Carlsbad, CA, USA) supplemented with glutamine, 10% fetal calf serum (FCS, Sigma-Aldrich, St. Louis, MO, USA), and 1% penicillin-streptomycin (Sigma-Aldrich, St. Louis, MO, USA). Primary human foreskin fibroblasts (HFF) and rat embryonic fibroblasts (REF), which were obtained from Sebastian Voigt (Berlin, Germany), were maintained in Eagle’s minimal essential medium (MEM, Gibco, Carlsbad, CA, USA) supplemented with 7% FCS, 1% Glutamax (Gibco, Carlsbad, CA, USA), and 1% penicillin-streptomycin (Sigma-Aldrich, St. Louis, MO, USA). HEK293T cells were transfected with 1–2 µg expression plasmid with Turbofect following the manufacturer’s instructions (Thermo Fisher, Waltham, MA, USA). The transfections for the luciferase and NanoBRET interaction assays were done with Lipofectamine 2000 (Invitrogen, Carlsbad, CA, USA) in HEK293T cells.
2.3. Stabilization Assay with Cycloheximide Treatment
HEK 293T cells were grown in six well dishes (7 × 105 cells/well). Cells were transfected with expression plasmids for IE1 homologs and human FEN1 aa 176-380 (hFEN1 aa 176-380), followed by the addition of cycloheximide (10 µg/mL, Sigma-Aldrich, St. Louis, MO, USA) at 14 h post-transfection. The cells were harvested after 0, 2, 4, and 6 h of incubation. The resulting lysates were then further analyzed by SDS-PAGE and Western Blotting.
2.4. NanoBRET Protein-Protein Interaction Assay
The protein-protein interaction assay was done using the NanoBRET-PPI Kit from Promega (#N1662, Fitchburg, MA, USA). The assay was generally performed following the instructions of the manufacturer. The following steps were changed: 6 × 105 HEK293T cells were seeded in six well dishes. One day later, the cells were transfected with plasmids encoding Halo- and Luc-tagged proteins using Lipofectamine 2000 reagent (Invitrogen, Carlsbad, CA, USA). The DNA ratio of IE1 to human PML and FEN1 aa 176-380 was 1:10, and to rat PML 1:100. After 24 h, the cells were washed once with 1 mL of PBS/EDTA and harvested with trypsin (Sigma-Aldrich, St. Louis, MO, USA). Then, cells were pelleted at 500 rpm for 5 min and resuspended in Opti-MEM™ I reduced serum medium without phenol red (Gibco, Carlsbad, CA, USA). Cell pools of 2.5 mL with 2 × 105 cells/mL were generated. One aliquot of the cells was used for the interaction assay; the other aliquot was used for Western Blot analysis. After the addition of the Halo ligand or DMSO to the pools, the cells were incubated for four h at 37 °C, and the detection of luminescence was done in a plate reader (Hidex Chameleon) for 5 s.
2.5. Generation of Lentiviruses and Transduction of HFFs and REFs
HFF and REF cells with doxycycline-inducible expression of human IE1 helix mutants were generated as described elsewhere [
14]. The cells were then cultivated in Dulbecco’s minimal essential medium (DMEM, Gibco, Carlsbad, CA, USA)) containing glutamine and supplemented with 10% Tet System Approved FCS (Takara Bio Inc., Shiga, Japan) and 1% penicillin-streptomycin (Sigma-Aldrich, St. Louis, MO, USA).
2.6. Indirect Immunofluorescence
HEK293T, HFF, and REF cells were seeded onto coverslips in six well dishes (8 × 104–1.5 × 105 cells/well). The cells were washed three times with 0.01 M PBS after transfection or 24 to 48 h after treatment with doxycycline (Sigma-Aldrich, St. Louis, MO, USA). The cells were fixed in a 4% paraformaldehyde solution (PFA) for 10 min at room temperature (RT). Afterward, the cells were washed four times with 0.01 M phosphate-buffered saline (PBS). The cells were permeabilized through incubation with 0.1% Triton X-100 in PBS for 10 min at 4 °C, followed by a washing step with PBS. After an additional incubation time of 5 min at RT, the respective first antibodies (diluted in PBS with 1% FCS) were added to the cells and incubated for 30 min at 37 °C. Unbound antibodies were removed by washing the cells three times with 0.01 M PBS. Afterwards, the cells were treated with the corresponding fluorescence-coupled secondary antibody diluted in 1% FCS in 0.01 M PBS for 30 min at 37 ˚C. The cells were mounted with DAPI-containing Vectashield mounting medium (Vector Laboratories, Newark, NJ, USA) and analyzed using a Zeiss Axio Observer Z1 with an Apotome2. The pictures were exported using the Zen software. For the quantification of PML foci, Z-series images of 50 cell nuclei per sample were taken, and the number of PML dots was assessed in maximum intensity projection images (0.24 μm distance). Following antibodies were used for immunofluorescence detection: mAb Flag M2 (F1804, Sigma-Aldrich, St. Louis, MO, USA), pAB Flag (F7425, Sigma-Aldrich, St. Louis, MO, USA); mAb PML 5E10 (kindly provided by Roel van Driel) to detect rat PML, and pAB PML A167 + A168 (#A301-167A, #A301-168A, Bethyl Laboratories, Montgomery, AL, USA) to detect human PML. Furthermore, pAB STAT2 from Santa Cruz Biotechnology (#H190, Dallas, TX, USA) was used.
2.7. Coimmunoprecipitation
HFF cells expressing human IE1, human IE1 1-382, rat IE1, and rat IE1 1-392 were grown in 10 cm dishes. The expression of the proteins was induced for 48 h with doxycycline (500 ng/mL). Afterwards, the cells were incubated with interferon β (IFNβ; 1000 U/mL; pbl assay science, NJ, USA) for 4 h and then harvested. For this, the cells were washed two times with PBS/EDTA and incubated with Trypsin (Sigma-Aldrich, St. Louis, MO, USA). The cells were harvested with PBS and pelleted at 1500 rpm for 5 min. Next, the cells were lysed in 800 µL lysis buffer (50 mM Tris-HCl [pH 8.0], 150 mM NaCl, 5 mM EDTA, 0.5% NP-40, 1 mM PMSF, 2 μg/mL of aprotinin, 2 μg/mL of leupeptin, and 2 μg/mL of pepstatin) for 30 min on ice. The cells were additionally sonicated for one minute using the QSonica Q700 Sonicator. One part of the lysed cells was used as input control. The other part of the lysates was incubated with anti-Flag antibody M2 (Sigma-Aldrich, St. Louis, MO, USA) coupled to protein-A-sepharose beads (Sigma-Aldrich, St. Louis, MO, USA) for 2 h at 4 °C. The beads were then centrifuged and washed four times with the lysis buffer. After the last step of centrifugation, the buffer was removed, and the beads were boiled in 4× SDS Buffer. The coimmunoprecipitation was then analyzed by SDS-PAGE and Western blotting.
2.8. Gaussia Luciferase Assay
HEK293T cells (6 × 10
5 cells/well) were grown in six well dishes and were transfected with a luciferase reporter plasmid containing the HCMV pUL112/113 promoter together with expression plasmids for IE2 and different IE1 homologs or mutants [
32]. The luciferase assay system of Promega (#E1501, Fitchburg, MA, USA) was used. The assay was performed as described by the manufacturer. The measurement was done using the plate reader SpectraMax iD5.
2.9. Western Blotting
Lysates from transfected cells were boiled with 4× SDS buffer for 10 min at 95 °C. The lysates were then sonicated for 1 min using the QSonica Q700 Sonicator (QSonica, Newton, MA, USA). Next, the samples were separated on 10% SDS polyacrylamide gels and transferred to PVDF membranes (BioRad, Feldkirchen, Grmany), followed by chemiluminescence detection using a FUSION FX7 imaging system (Vilber Lourmat, Eberhardzell, Germany). Following antibodies were used: mAB Flag M2 (Sigma-Aldrich, St. Louis, MO, USA), pAB human STAT2 H190 (Santa Cruz, Dallas, TX, USA), mAB β-Actin AC15 (Sigma-Aldrich, St. Louis, MO, USA), mAB Halo (G921A, Promega, Fitchburg, MA, USA), pAB IE2 (pAB178; [
28]).
2.10. RNA Isolation and qPCR
HFF cells expressing human IE1, human IE1 1-382, rat IE1, and rat IE1 1-392 and control cells were grown as triplicates in six well dishes. One day later, the expression of IE1 homologs was induced by adding doxycycline (10 µg/mL). After 24 h incubation, the cells were treated with IFNβ (1000 U/mL) for an additional 24 h. Next, RNA was isolated using the Direct-zol RNA Miniprep Kit (#R2052) from Zymo Research (Irvine, CA, USA) according to the manufacturer’s instructions. Afterward, the Maxima First Strand cDNA synthesis kit (#K1672, Thermo Fisher, Waltham, MA, USA) was used to generate the first strand cDNA. For the reaction, 0.1–0.5 µg RNA was used as a template. The cDNA was diluted 1:4, and 2 µL were used as a template for quantitative real-time PCR (qPCR). Therefore, a master mix was generated that included 10 µL Sso Advanced Univ SYBR Grn. Supermix (BioRad, Feldkirchen, Germany), 7.85 µL nuclease-free water, and 0.15 µL of the following qPCR Primer pair mix: CCL5, CCL8 (Biomol, Hamburg, Germany). Furthermore, the forward and reverse primer of ISG54 were purchased from Biomers (Ulm, Germany). Additionally, the housekeeping gene actin was used for normalisation of PCR values. For this, the primer pair mix for actin (Biomol, Hamburg, Germany) was used. The assay was performed in a 96-well plate, and the Agilent AriaMx Real-time PCR System together with the corresponding software Agilent Aria 1.5 (Agilent Technologies, Santa Clara, CA, USA) was used. The qPCR reaction started with activation of the polymerase for 30 s at 95°C. Afterward, primer binding and strand elongation were done at 60 °C for 30 s for 40 cycles. For the final step, the reaction specificity was verified by a dissociation stage.
2.11. Crystal Structure and Bioinformatic Analysis
3D structure predictions of IE1 helix swap mutants were generated using AlphaFold Colab in multimer mode (
https://colab.research.google.com/github/deepmind/alphafold/blob/main/notebooks/AlphaFold.ipynb (accessed on 13 June.2022)) [
34,
35]. As IE1 is known to dimerize [
7,
14], the models comprising the residues 14-382 of hIE1 were predicted using the multimer mode of AlphaFold; however, only monomers are shown. Structure illustrations were generated with PyMol (PyMOL Molecular Graphics System, Version 1.3. Schrödinger, LLC, New York, NY, USA).
4. Discussion
IE1 expressed at the beginning of an HCMV infection can counteract innate and intrinsic immune mechanisms. Previous studies demonstrated that PML-NBs form an important intrinsic immune barrier against cytomegaloviruses [
10,
12,
30]. Human IE1 interacts with the main organizer PML, followed by deSUMOylation and disruption of PML NBs. Schweininger et al. showed that the mechanism of antagonizing PML by IE1 seems to be conserved across different species during evolution [
14]. Until now, the exact interaction surface of IE1 and PML is unknown. Different groups analyzed the interaction of IE1 with PML using co-immunoprecipitation [
7,
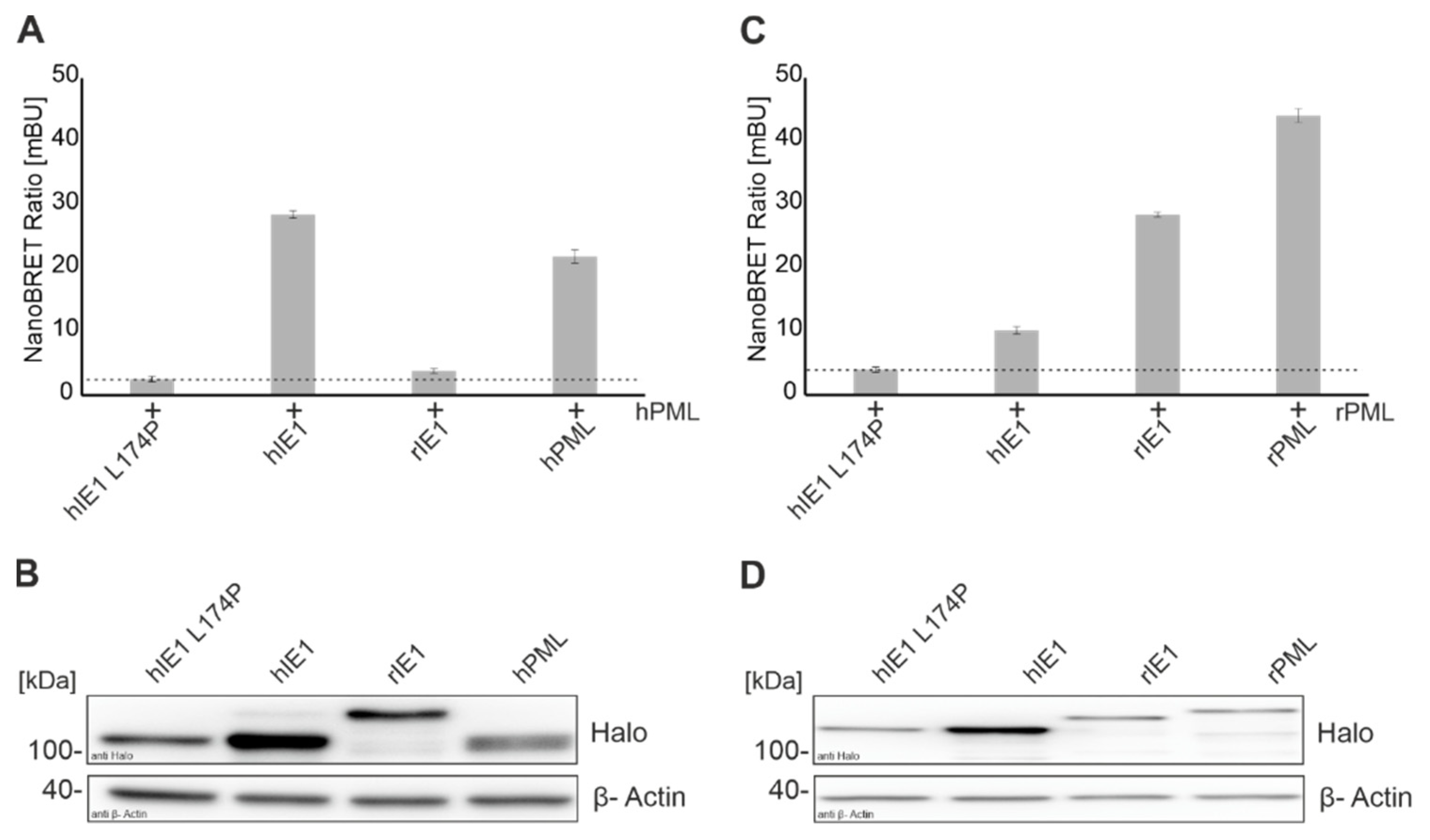
31]. In the present study, we successfully established a NanoBRET protein-protein interaction assay to measure the interaction of viral proteins with different cellular proteins in a sensitive and quantitative manner (
Figure 1). The interaction of IE1 homologs with their natural host PMLs that had already been detected by various groups with distinct methodology could be confirmed [
7,
14]. In the past years, different hIE1 mutants were generated in an attempt to define the interaction surface of IE1 with PML. NanoBRET analyses verified the observation that the hIE1 L174P mutant no longer interacts with hPML [
29]. Surprisingly, however, we detected that hIE1cc172-176, recently described by Paulus et al. as a mutant with wild-type characteristics except for lacking PML binding, still interacted with hPML in a manner comparable to unmodified hIE1 (
Figure 3A) [
31]. We conclude that clustered charge-to-alanine mutations at positions 172-176 do not abrogate PML binding. Consequently, results obtained with hIE1cc172-176 need to be interpreted with caution [
31].
Schweininger et al. solved the crystal structures of the human and rat IE1 core domain [
14]. We used the similarities and differences in structure and sequence to search for a potential interaction surface of IE1 with PML. We successfully generated five stable hIE1 core mutants that carry one or two helices of rIE1 (
Figure 2). Analyses with AlphaFold prediction suggested that swapping helices should not affect the overall folding of the respective chimeric proteins (
Figure 2B). Consistently, five out of six mutants were stably expressed. Only mutant hIE1_rH2 could not be detected in transient expression experiments (
Figure 2C). Since helix 2 in rIE1 is displaced by a rotation of 90°, this different angle in hIE1 might cause misfolding of the hIE1_rH2 mutant followed by degradation [
14]. Furthermore, immunofluorescence analyses showed that mutants that carry helix 1 of rIE1 were present both in the cytoplasm and nucleus (
Figure 2 D). The aa 1-24 in hIE1 function as a nuclear localisation signal [
12]. Due to helix exchange, results obtained with mutants hIE1_rH1 and hIE1_rH1/2 revealed that rat helix 1 does either not contain an NLS or the NLS activity contained within rat helix 1 is considerably weaker. This would reinforce the prediction of Sandford et al. for a potential NLS in the C-terminus of rIE1 [
17]. NanoBRET measurements demonstrated that the chimeric proteins were no longer able to interact with PML homologs (
Figure 3). Moreover, immunofluorescence analyses showed that loss of PML interaction correlated with a defect in PML-NB disruption (
Figure 4). Thus, based on these results, we assume that IE1 requires a large surface that comprises more than one helix for successful interaction with PML. This might also explain why the exchange of single amino acids or clustered mutations of IE1 was not able to abrogate PML binding.
In addition to PML, hFEN1 could recently be identified as a binding partner of hIE1 that is hijacked for promoting viral replication [
18]. NanoBRET analyses and cycloheximide stabilization assays revealed that rIE1 is neither able to interact with hFEN1 176-380 nor does it mediate hFEN1 protein stabilization (
Figure 5A, bar 1). These results indicate that the interaction between hFEN1 and hIE1 is also species-specific. Interestingly, however, we observed that the hIE1 chimeric proteins bearing either helices 4, 8, or 10/11 of rIE1 (hIE1_rH4, hIE1_rH8 and hIE1_rH10/11, respectively) were still able to bind hFEN1 (
Figure 5A, bars 5–7). Since mutants that carry helix 1 or helices 1/2 of rIE1 lost their interaction with hFEN1, we assume that the region comprising aa 1-82 of hIE1 may constitute the binding surface for FEN1 or may form an important part of it. In the previous analysis, we detected that mutation of residues located in helices H7 and H9 of hIE1 also interferes with FEN1 binding [
18]. One potential explanation for this may be that IE1 dimerization is a prerequisite for hFEN1 interaction, and this may be affected by mutations in helices H7 and H9. Mutants hIE1_rH1 and hIE1_rH4 were then tested for their ability to stabilize hFEN1 176-380. Surprisingly, both mutants exhibited a defect in hFEN1 stabilization while hIE1_rH4 was still able to bind hFEN1. We observed in previous experiments that the binding of hIE1 to hFEN1 not only increases the half-life of the protein but also leads to an accumulation of a hyperphosphorylated hFEN1 isoform [
18]. Thus, one may speculate that hIE1 needs to recruit additional cellular factors for hEN1 protein stabilization that interact with C-terminal regions of IE1 affected by the exchange of helix 4.
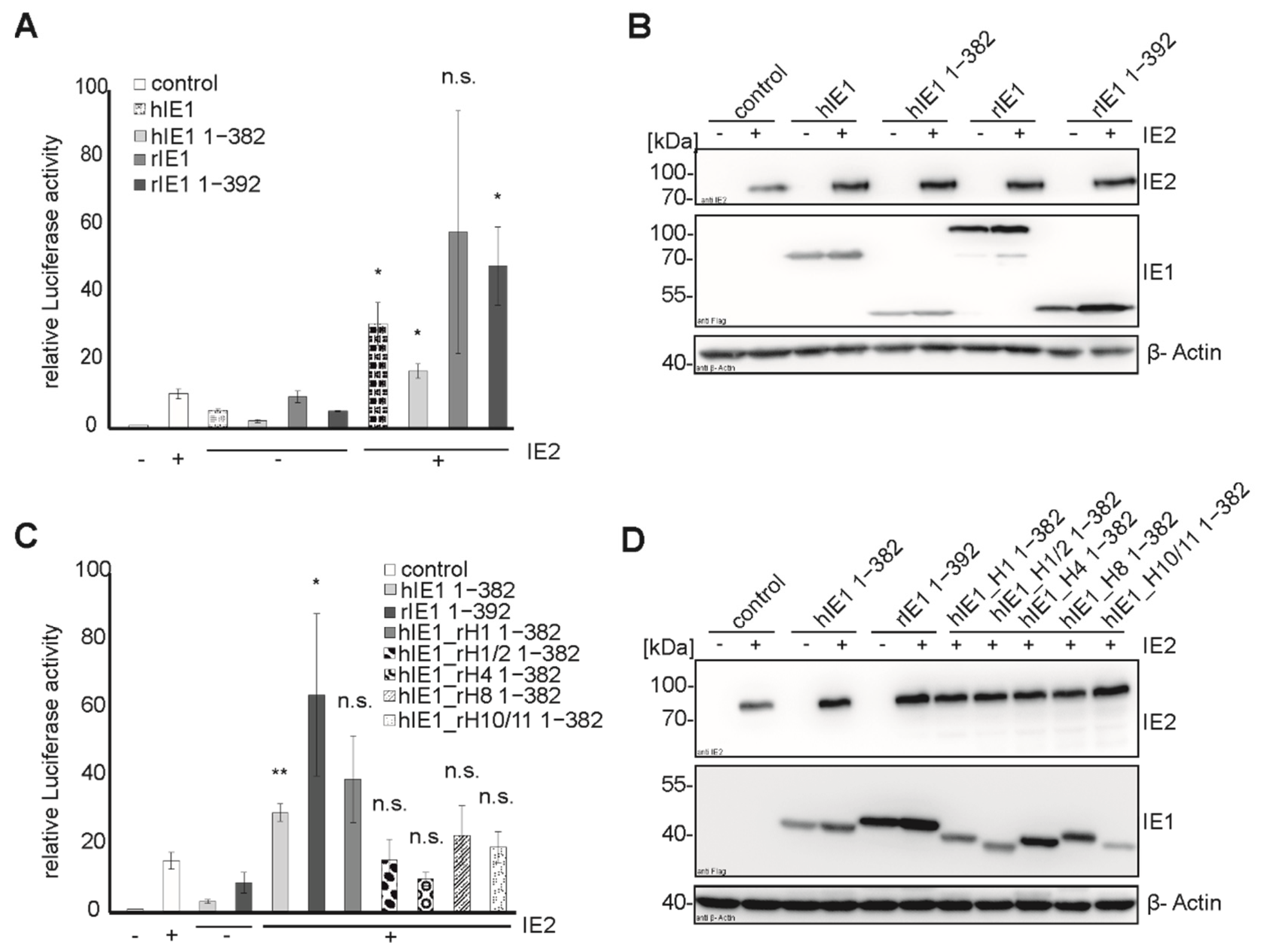
Concerning the transactivation capacity of the major immediate early proteins of rat CMV, Sandford et al. described that rIE2 could transactivate viral promotors due to similarities in sequence with hIE2 [
17]. Despite the low sequence identity of hIE1 and rIE1, we detected a transactivation activity for the core domain of rIE1, as also observed for hIE1, using luciferase assays (
Figure 6A,B). Tripathi et al. showed that a possible transactivation site of hIE1 is located in the core domain [
16]. Consequently, the integrity of transactivation of the hybrid proteins was tested. Mutant hIE1_rH1 1-382 could transactivate the promoter in the presence of hIE2 like the hIE1 1-382, while hIE1_rH1/2, hIE1_rH4, hIE1_rH8, and hIE1_H10/11 showed either reduced or no activity. The fact that hIE1_rH1 has a strong transactivation capacity suggests that this chimeric protein, which no longer interacts with FEN1, is expressed as a stable and functional protein. Furthermore, this observation supports the assumption that the interaction site of FEN1 is localized within helix 1 of hIE1. In summary, the exchange of helices at different positions of the IE1 core domain affected distinct functions of hIE1. Although all mutants failed to bind PML, other functions were not abrogated by swapping entire helices between hIE1 and rIE1.
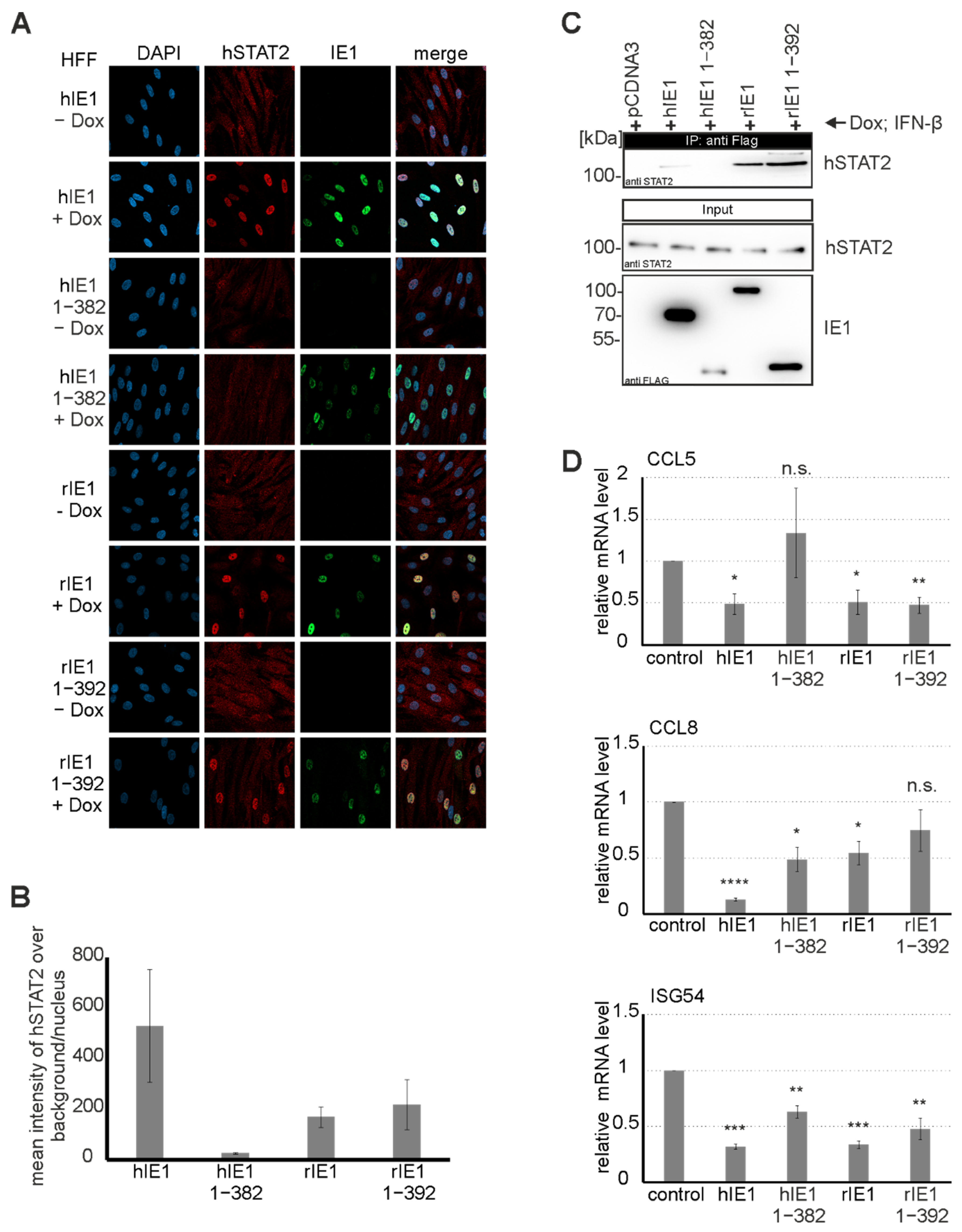
Since hIE1 is known to act as a potent interferon antagonist via STAT2 binding at C-terminal sequences, we were interested in investigating whether rIE1 can also modulate STAT2 [
23]. Previous studies demonstrated that hIE1 induces an intranuclear relocalisation of STAT2 that abates ISG expression [
23,
38]. Surprisingly, we observed that full-length rIE1 and rIE1 1-392 lacking the highly unordered C-terminus could induce STAT2 relocalization (
Figure 7A). Moreover, rIE1 1-392 exhibited a strong interaction with STAT2 in coimmunoprecipitation experiments indicating that the core domain of rIE1 is sufficient for STAT2 binding (
Figure 7B). Interaction with STAT2 correlated with significantly reduced expression levels of selected ISGs upon coexpression of the respective IE1 proteins. This suggests that antagonization of STAT2 by IE1 is conserved and occurs in a species-independent manner. However, primate and rodent CMV IE1 proteins may have evolved distinct binding surfaces to fulfill this task.
Furthermore, hIE1 has been shown to contain a C-terminal chromatin tethering domain (CTD) that plays a role in association with mitotic chromosomes [
25,
26,
39]. Immunofluorescence analyses revealed that rIE1 could neither bind to rats nor human mitotic chromatin (
Figure 8). This observation is in line with the observation that rIE1 lacks a sequence similar to the CTD of hIE1 [
8,
26]. Moreover, our results indicate that hIE1 can colocalize with chromatin of different species. Previous studies demonstrated that the CTD is dispensable for productive replication in asynchronous fibroblast cells [
8,
25]. However, as suggested by a recent publication using a CTD-deficient recombinant in the Towne strain of HCMV, this domain may be important for maintaining the viral genome during mitosis [
39]. If the CTD of hIE1 indeed acts as a genome maintenance factor, one may speculate that either RCMV lacks this function or encodes a different protein that serves to maintain viral genomes during mitosis.


 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}







