
Hemagglutinin Subtype Specificity and Mechanisms of Highly Pathogenic Avian Influenza Virus Genesis

 , ,
, ,
Abstract
:1. Introduction
2. Historical Emergences of HPAIVs

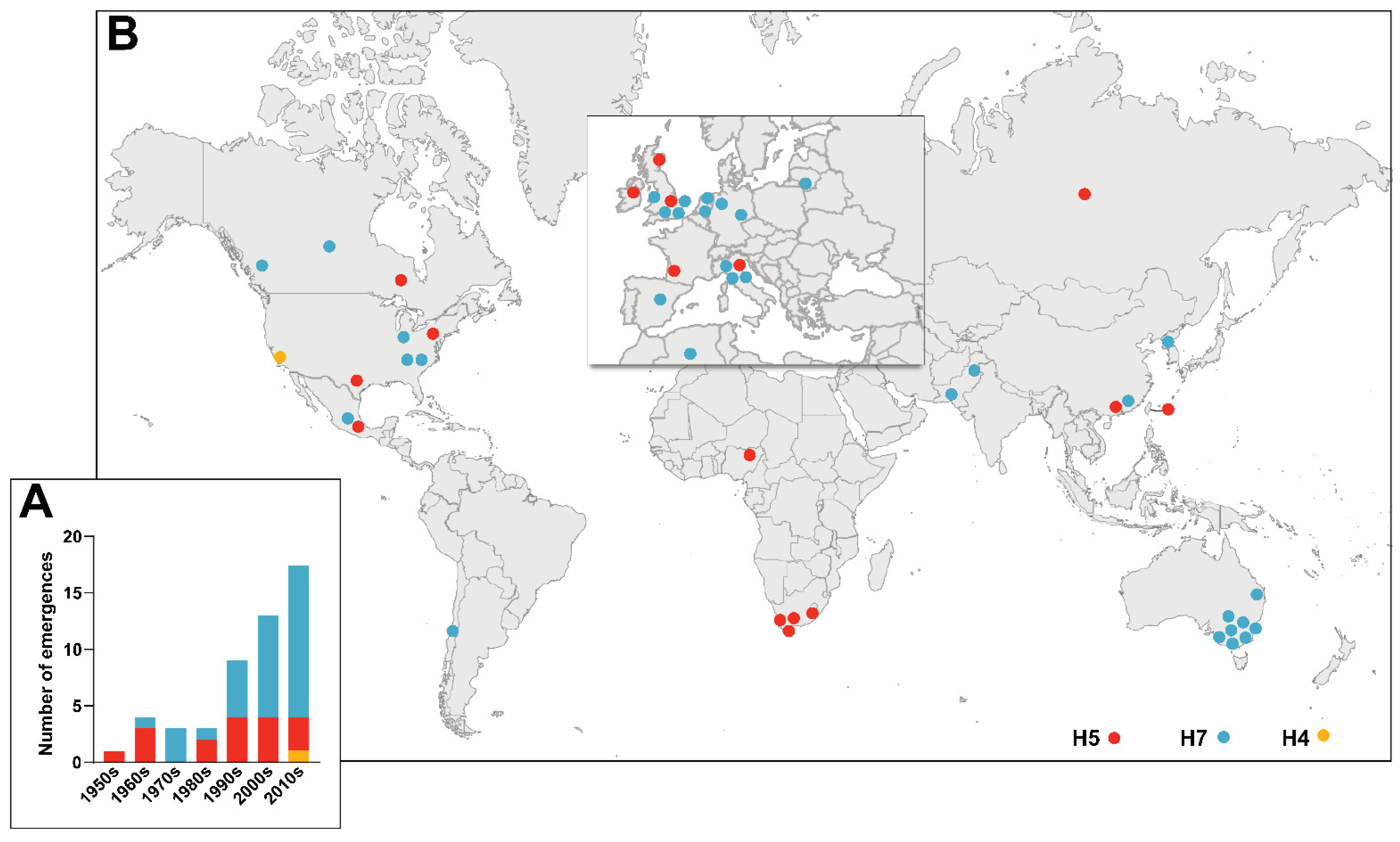{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Year | Country | Subtype | Cleavage Site Sequence | HA Accession Number | IVPI (Phenotype a) | Progenitor Detection b | HA Accession Number Progenitor | Number of Affected Premises (Species) | Ref. |
|---|---|---|---|---|---|---|---|---|---|
| 1959 | Scotland | H5N1 | PQRKKR*G c | GU052518 | N.D. d (HP) | No e | N.A. f | 2 (unknown) | [14] |
| 1961 | South Africa | H5N3 | PQRETRRQKR*G | GU052822 | N.D. (HP) | No | N.A. | N.A. g (common terns) | [34,35,36] |
| 1963 | England | H7N3 | PKRRRR*G | AF202238 | N.D. (HP) | No | N.A. | 3 (turkeys) | [37,38] |
| 1966 | Canada | H5N9 | PQRRKKR*G | CY107859 | N.D. (HP) | Yes | CY087808 | 1 (turkeys) | [39,40] |
| 1967 | USSR | H5N1 | unknown | N.A. | N.D. (HP) | No | N.A. | 1 (chickens) | [41] |
| 1976 | Australia | H7N7 | PEIPKKKEKR*G | CY024786 | N.D. (HP) | No h | N.A. | 1 (chickens) | [42,43] |
| 1979 | England | H7N7 | PEIPKKRKR*G PEIPKRRRR*G PEIPKKREKR*G | N.A. | N.D. (HP) | No | N.A. | 3 (turkeys) | [44] |
| 1979 | Germany | H7N7 | PEIPKKKKR*G PEIPKRKKR*G PEIPKKKKKKR*G PEIPKKRKKR*G PETPKKKKKKR*G | U20459 L43913 CY107844 L43915 L43914 | N.D. (HP) | No i | N.A. | 2 (chickens, geese) | [45] |
| 1983–1984 | USA | H5N2 | PQKKKR*G | GU052771 | 2.37 (HP) | Yes | J04325 | 356 (chickens, turkeys, guinea fowl, chuckar) | [46,47,48] |
| 1983 | Ireland | H5N8 | PQRKRKKR*G | M18451 | 2.76 (HP) | No | N.A. | 4 (turkeys, ducks, chickens) | [49] |
| 1985 | Australia | H7N7 | PEIPKKREKR*G | M17735 | N.D. (HP) | No | N.A. | 1 (chickens) | [50,51] |
| 1991 | England | H5N1 | PQRKRKTR*G | GU052510 | 3 (HP) | No | N.A. | 1 (turkeys) | [52] |
| 1992 | Australia | H7N3 | PEIPKKKKR*G | AF202227 | 2.71 (HP) | No | N.A. | 1 (chickens, ducks) | [53,54] |
| 1994–1995 | Australia | H7N3 | PEIPRKRKR*G | CY022685 | N.D. (HP) | No | N.A. | 1 (chickens) | [54] |
| 1994–1995 | Mexico | H5N2 | PQRKRKTR*G PQRKRKRKTR*G | AB558473 U85390 | N.D. (HP) | Yes | GU186573 | 360 (chickens) | [55,56,57] |
| 1994–1995 | Pakistan | H7N3 | PETPKRKRKR*G PETPKRRKR*G | AF202226 AF202230 | N.D. (HP) | No | N.A. | Many (chickens) | [58,59] |
| 1996– now | China | H5N1 + H5Nx j | PQRERRRKKR*G ( + many variations) | AF144305 | 2.1 (HP) | No | N.A. | Many (many) | [60] |
| 1997 | Australia | H7N4 | PEIPRKRKR*G PEIPRKRKR*G | AY943924 CY022701 | 2.52 k (HP) | No | N.A. | 3 (chickens, emu) | [61] |
| 1997–1998 | Italy | H5N2 | PQRRRKKR*G | CY017403 | 2.98–3 (HP) | No | N.A. | 8 outbreaks (chickens, ducks, geese, turkeys, guinea fowl, pigeons) | [62] |
| 1999–2000 | Italy | H7N1 | PKGSRVRR*G | CY021405 | 3 (HP) | Yes | GU052999 | 413 outbreaks (chickens, turkeys, guinea fowl, ducks, pheasants, quails, ostriches) | [63,64] |
| 2002 | Chile | H7N3 | PEKPKTCSPLSRCRETR*G PEKPKTCSPLSRCRKTR*G | AY303631 AY303632 | 2.24–3 (HP) | Yes | AY303630 | 4 (chickens, turkeys) | [65,66,67] |
| 2003 | Netherlands | H7N7 | PEIPKRRRR*G | AY338458 | 2.93 (HP) | No l | N.A. | 255 (chickens) | [68,69] |
| 2003 | Netherlands | H7N3 | PEIPKGSRVRR*G | EPI1595425 | 2.4 (HP) | Yes | EPI1595417 | N.A. m (turkeys) | [70,71] |
| 2003 | Pakistan | H7N3 | PETPKRRKR*G | HM346493 | 2.8 (HP) | No | N.A. | 522 (chickens) | [59,72] |
| 2004 | USA | H5N2 | PQRKKR*G | AY849793 | 0 (LP) | No | N.A. | 6 (chickens) | [73] |
| 2004 | Canada | H7N3 | PENPKQAYRKRMTR*G (+ many variations) | AY648287 | 2.87 (HP) | Yes | AY650270 | 42 (chickens) | [74] |
| 2004 | South Africa | H5N2 | PQREKRRKKR*G | FJ519983 | 0.63 n (HP) | No | N.A. | 38 (ostriches) | [75] |
| 2005 | North Korea | H7N7 | PEIPKGRHRRPKR*G | N.A. | N.D. (HP) | No | N.A. | 3 (chickens | [76] |
| 2006 | South Africa | H5N2 | PQRRKKR*G | EF591749 | N.D. (HP) | Yes | EF591757 | 24 (ostriches) | [77] |
| 2007 | Canada | H7N3 | PENPKTTKPRPRR*G | EU500860 | 3 (HP) | No | N.A. | 1 (chickens) | [78] |
| 2007 | Nigeria | H5N2 | KEKRRKKR*G REKRRKKR*G | N.A. | N.D. (LPo) | No | N.A. | N.A. o (duck, geese) | [79] |
| 2008 | England | H7N7 | PEIPKRKKR*G PEIPKKKKR*G PEIPKKKKKKR*G | FJ476173 | 2.95–3 (HP) | No | N.A. | 1 (chickens) | [80] |
| 2009 | Spain | H7N7 | PKGTKPRPRR*G | GU121458 | N.D. (HP) | No | N.A. | 1 (chickens) | [81] |
| 2011–2013 | South Africa | H5N2 | PQRRKKR*G PQRRRKKR*G PQRKRKKR*G PQRRRKR*G | JX069081 | 0.8–1.37 p (HP) | No | N.A. | 50 (ostriches) | [82] |
| 2012 | Australia | H7N7 | PEIPRKRKR*G | N.A. | N.D. (HP) | No | N.A. | 1 (chickens) | [83] |
| 2012–2013 | Taiwan | H5N2 | PQRKKR*G PQRRKR*G | KJ162620 KF193394 | 2.91 (HP) | No q | N.A. | 5 (chickens) | [84] |
| 2012–now | Mexico | H7N3 | PENPKDRKSRHRRTR*G | JX908509 | N.D. (HP) | No | N.A. | Many (chickens) | [85,86] |
| 2012 | USA | H4N2 | PEKRRTR*G | KF986862 | 0 (LP) | No | N.A. | 1 (quail) | [17] |
| 2013 | Italy | H7N7 | PKRKRR*G PKRKRR*G PKRRERR*G PKRKRR*G | KF569186 | N.D. (HP) | No | N.A. | 6 (chickens, turkeys) | [87,88] |
| 2013 | Australia | H7N2 | PEIPRKRKR*G | N.A. | N.D. (HP) | No | N.A. | 2 (chickens) | [89] |
| 2015 | England | H7N7 | PEIPRHRKGR*G PEIPRHRKRR*G | EPI623939 | 2.52 (HP) | No | N.A. | 1 (chickens) | [90] |
| 2015 | Germany | H7N7 | PEIPKRKRR*G | EPI634885 | N.D. (HP) | Yes | EPI624526 | 1 (chickens) | [91] |
| 2015–2016 | France | H5(N1, N2,N9) | HQRRKR*G | H5N1:KU310447 H5N2:KX014878 H5N9:KX014886 | H5N1: 2.9 (HP) | No | N.A. | 81 (chickens, ducks, geese, guinea fowl) | [92,93] |
| 2016 | Algeria | H7N1 | unknown | N.A. | N.D. (HP) | No | N.A. | N.A. r (many wild bird species) | [94] |
| 2016 | USA | H7N8 | PKKRKTR*G | KU558906 | N.D. (HP) | Yes | EPI709576 | 1 (turkeys) | [95,96] |
| 2016 | Italy | H7N7 | PELPKGRKRR*G PELPKRRERR*G | EPI756028 | N.D. (HP) | No | N.A. | 2 (chickens, turkeys) | [97] |
| 2016–now | China | H7N9 | PEVPKGKRTAR*G PEVPKRKRTAR*G PEVPKGKRIAR*G | EPI919533 EPI917102 | 2.92 (HP) | No s | N.A. | Many (primarily chickens) | [29,30] |
| 2017 | USA | H7N9 | PENPKTDRKSRHRRIR*G | MF357740 | N.D. (HP) | Yes | MF357732 | 2 (chickens) | [98] |
| 2020 | USA | H7N3 | PENPKTDRKSRHRRIR*G | EPI1775733 | 2.46 (HP) | Yes | MT444363 | 13 (turkeys) | [99] |
| 2020 | Australia | H7N7 | unknown | N.A. | N.D. (HP) | No | N.A. | 3 (chickens) | [100] |
| 2021 | Lithuania | H7N7 | unknown | N.A. | N.D. (HP) | No | N.A. | N.A.t (mute swan) | [101] |
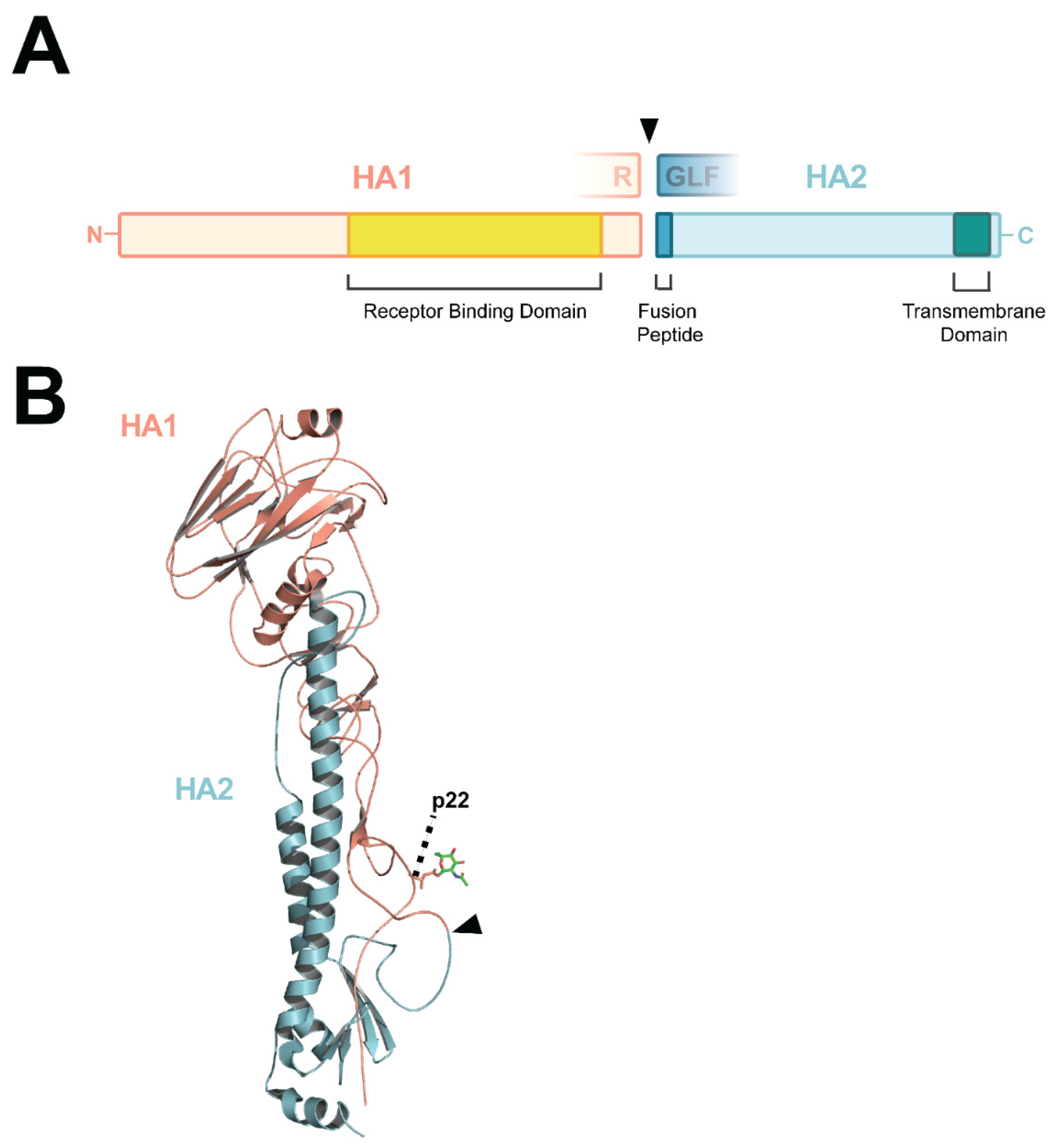
3. HA Cleavage and Virulence
4. Proteases That Activate HA
5. MBCS Amino Acid Composition
| Viral Strain | Cleavage Site Sequence | Trypsin-Independent HA Cleavage | Putative GS p22 a | IVPI b (Phenotype c) | Ref. |
|---|---|---|---|---|---|
| A/chicken/Scotland/1959 (H5N1) | PQRKKR*G d | + | - | N.D. e (HP) | [171] |
| A/chicken/Pennsylvania/1/1983 (H5N2) | PQKKKR*G | - | + | 0 (LP) | [171,172] |
| A/chicken/Pennsylvania/1370/1983 (H5N2) | PQKKKR*G | + | - | 2.37 (HP) | [171,172] |
| A/chicken/Texas/298313/2004 (H5N2) | PQRKKR*G | - | + | 0 (LP) | [73] |
| A/chicken/Taiwan/A703-1/2008 (H5N2) | PQRKKR*G | + | + | 0.89 (LP) | [174,175] |
| H5N2 HPAIV from Taiwan 2012 f | PQRRKR*G | N.D. | + | 2.91 (HP) | [176] |
| A/chicken/France/150169a/2015 (H5N1) | HQRRKR*G | N.D. | + | 2.9 (HP) | [93] |
6. Subtype Restriction of MBCS Acquisition: Compatibility of an MBCS at the Protein Level
7. Mechanisms of MBCS Acquisition
7.1. Nucleotide Substitutions
7.2. Influenza Virus RdRp Stuttering and Backtracking
7.2.1. Influence of the Nucleotide Sequence on Influenza Virus RdRp Stuttering and Backtracking
7.2.2. Influence of RNA Structure on Influenza Virus RdRp Stuttering and Backtracking
7.3. Non-Homologous Recombination
8. Concluding Remarks
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Stallknecht, D.E.; Shane, S.M. Host range of avian influenza virus in free-living birds. Vet. Res. Commun. 1988, 12, 125–141. [Google Scholar] [CrossRef] [PubMed]
- Olsen, B.; Munster, V.J.; Wallensten, A.; Waldenström, J.; Osterhaus, A.D.M.E.; Fouchier, R.A.M. Global patterns of influenza A virus in wild birds. Science 2006, 312, 384–388. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Daoust, P.Y.; Kibenge, F.S.B.; Fouchier, R.A.M.; van de Bildt, M.W.G.; Van Riel, D.; Kuiken, T. Replication of low pathogenic avian influenza virus in naturally infected mallard ducks (Anas platyrhynchos) causes no morphologic lesions. J. Wildl. Dis. 2011, 47, 401–409. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kuiken, T. Is low pathogenic avian influenza virus virulent for wild waterbirds? Proc. R. Soc. B Biol. Sci. 2013, 280, 20130990. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Webster, R.G.; Yakhno, M.; Hinshaw, V.S.; Bean, W.J.; Copal Murti, K. Intestinal influenza: Replication and characterization of influenza viruses in ducks. Virology 1978, 84, 268–278. [Google Scholar] [CrossRef]
- Slemons, R.D.; Easterday, B.C. Virus replication in the digestive tract of ducks exposed by aerosol to type-A influenza. Avian Dis. 1978, 22, 367–377. [Google Scholar] [CrossRef]
- Munster, V.J.; Baas, C.; Lexmond, P.; Bestebroer, T.M.; Guldemeester, J.; Beyer, W.E.P.; De Wit, E.; Schutten, M.; Rimmelzwaan, G.F.; Osterhaus, A.D.M.E.; et al. Practical considerations for high-throughput influenza A virus surveillance studies of wild birds by use of molecular diagnostic tests. J. Clin. Microbiol. 2009, 47, 666–673. [Google Scholar] [CrossRef] [Green Version]
- Pepin, K.M.; Spackman, E.; Brown, J.D.; Pabilonia, K.L.; Garber, L.P.; Weaver, J.T.; Kennedy, D.A.; Patyk, K.A.; Huyvaert, K.P.; Miller, R.S.; et al. Using quantitative disease dynamics as a tool for guiding response to avian influenza in poultry in the United States of America. Prev. Vet. Med. 2014, 113, 376–397. [Google Scholar] [CrossRef] [Green Version]
- Bergervoet, S.A.; Pritz-Verschuren, S.B.E.; Gonzales, J.L.; Bossers, A.; Poen, M.J.; Dutta, J.; Khan, Z.; Kriti, D.; van Bakel, H.; Bouwstra, R.; et al. Circulation of low pathogenic avian influenza (LPAI) viruses in wild birds and poultry in the Netherlands, 2006–2016. Sci. Rep. 2019, 9, 13681. [Google Scholar] [CrossRef]
- França, M.S.; Brown, J.D. Influenza pathobiology and pathogenesis in avian species. Curr. Top. Microbiol. Immunol. 2014, 385, 221–242. [Google Scholar] [CrossRef]
- Office International des Épizooties. Avian Influenza (Including Infection with High Pathogenicity Avian Influenza Viruses); OIE: Paris, France, 2021; Chapter 3.3.4; Available online: https://www.oie.int/fileadmin/Home/eng/Health_standards/tahm/3.03.04_AI.pdf (accessed on 5 August 2021).
- Perroncito, E. Epizoozia tifoide nei gallinacei. Ann. Accad. Agric. 1878, 21, 87–126. [Google Scholar]
- Kaleta, E.F.; Rülke, C.P.A. The Beginning and Spread of Fowl Plague (H7 High Pathogenicity Avian Influenza) across Europe and Asia (1878–1955). In Avian Influenza; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 2009; pp. 145–189. ISBN 9780813820477. [Google Scholar]
- Pereira, H.G.; Tůmová, B.; Law, V.G. Avian influenza A viruses. Bull. World Health Organ. 1965, 32, 855–860. [Google Scholar] [PubMed]
- Lee, D.H.; Criado, M.F.; Swayne, D.E. Pathobiological origins and evolutionary history of highly pathogenic avian influenza viruses. Cold Spring Harb. Perspect. Med. 2021, 11, 1–22. [Google Scholar] [CrossRef] [Green Version]
- Dhingra, M.S.; Artois, J.; Dellicour, S.; Lemey, P.; Dauphin, G.; Von Dobschuetz, S.; Van Boeckel, T.; Castellan, D.M.; Morzaria, S.; Gilbert, M. Geographical and Historical Patterns in the Emergences of Novel Highly Pathogenic Avian Influenza (HPAI) H5 and H7 Viruses in Poultry. Front. Vet. Sci. 2018, 5, 84. [Google Scholar] [CrossRef] [Green Version]
- Wong, S.S.; Yoon, S.W.; Zanin, M.; Song, M.S.; Oshansky, C.; Zaraket, H.; Sonnberg, S.; Rubrum, A.; Seiler, P.; Ferguson, A.; et al. Characterization of an H4N2 influenza virus from Quails with a multibasic motif in the hemagglutinin cleavage site. Virology 2014, 468–470, 72–80. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Röhm, C.; Horimoto, T.; Kawaoka, Y.; Süss, J.; Webster, R.G. Do Hemagglutinin Genes of Highly Pathogenic Avian influenza Viruses Constitute Unique Phylogenetic Lineages? Virology 1995, 209, 664–670. [Google Scholar] [CrossRef] [Green Version]
- Chan, P.K.S. Outbreak of avian influenza A(H5N1) virus infection in Hong Kong in 1997. Clin. Infect. Dis. 2002, 34, 58–64. [Google Scholar] [CrossRef]
- De Jong, J.C.; Claas, E.C.J.; Osterhaus, A.D.M.E.; Webster, R.G.; Lim, W.L. A pandemic warning? Nature 1997, 389, 554. [Google Scholar] [CrossRef]
- Li, K.S.; Guan, Y.; Wang, J.; Smith, G.J.D.; Xu, K.M.; Duan, L.; Rahardjo, A.P.; Puthavathana, P.; Buranathai, C.; Nguyen, T.D.; et al. Genesis of a highly pathogenic and potentially pandemic H5N1 influenza virus in eastern Asia. Nature 2004, 430, 209–213. [Google Scholar] [CrossRef]
- Kilpatrick, A.M.; Chmura, A.A.; Gibbons, D.W.; Fleischer, R.C.; Marra, P.P.; Daszak, P. Predicting the global spread of H5N1 avian influenza. Proc. Natl. Acad. Sci. USA 2006, 103, 19368–19373. [Google Scholar] [CrossRef] [Green Version]
- Lycett, S.J.; Bodewes, R.; Pohlmann, A.; Banks, J.; Bányai, K.; Boni, M.F.; Bouwstra, R.; Breed, A.C.; Brown, I.H.; Chen, H.; et al. Role for migratory wild birds in the global spread of avian influenza H5N8. Science 2016, 354, 213. [Google Scholar] [CrossRef] [Green Version]
- Ducatez, M.F.; Olinger, C.M.; Owoade, A.A.; De Landtsheer, S.; Ammerlaan, W.; Niesters, H.G.M.; Osterhaus, A.D.M.E.; Fouchier, R.A.M.; Muller, C.P. Avian flu: Multiple introductions of H5N1 in Nigeria. Nature 2006, 442, 37. [Google Scholar] [CrossRef] [PubMed]
- World Health Organization. Avian Influenza Weekly Update Number 832. Available online: https://www.who.int/docs/default-source/wpro---documents/emergency/surveillance/avian-influenza/ai_20220218.pdf?sfvrsn=5f006f99_87 (accessed on 4 March 2022).
- Youk, S.; Lee, D.H.; Ferreira, H.L.; Afonso, C.L.; Absalon, A.E.; Swayne, D.E.; Suarez, D.L.; Pantin-Jackwood, M.J. Rapid evolution of Mexican H7N3 highly pathogenic avian influenza viruses in poultry. PLoS ONE 2019, 14, 0222457. [Google Scholar] [CrossRef] [Green Version]
- Lopez-Martinez, I.; Balish, A.; Barrera-Badillo, G.; Jones, J.; Nuñez-García, T.E.; Jang, Y.; Aparicio-Antonio, R.; Azziz-Baumgartner, E.; Belser, J.A.; Ramirez-Gonzalez, J.E.; et al. Highly pathogenic avian influenza A(H7N3) virus in poultry workers, Mexico, 2012. Emerg. Infect. Dis. 2013, 19, 1531–1534. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Jiang, H.; Wu, P.; Uyeki, T.M.; Feng, L.; Lai, S.; Wang, L.; Huo, X.; Xu, K.; Chen, E.; et al. Epidemiology of avian influenza A H7N9 virus in human beings across five epidemics in mainland China, 2013–2017: An epidemiological study of laboratory-confirmed case series. Lancet. Infect. Dis. 2017, 17, 822–832. [Google Scholar] [CrossRef]
- Yang, L.; Zhu, W.; Li, X.; Chen, M.; Wu, J.; Yu, P.; Qi, S.; Huang, Y.; Shi, W.; Dong, J.; et al. Genesis and Spread of Newly Emerged Highly Pathogenic H7N9 Avian Viruses in Mainland China. J. Virol. 2017, 91, e01277-17. [Google Scholar] [CrossRef] [Green Version]
- Qi, W.; Jia, W.; Liu, D.; Li, J.; Bi, Y.; Xie, S.; Li, B.; Hu, T.; Du, Y.; Xing, L.; et al. Emergence and Adaptation of a Novel Highly Pathogenic H7N9 Influenza Virus in Birds and Humans from a 2013 Human-Infecting Low-Pathogenic Ancestor. J. Virol. 2018, 92, e00921-17. [Google Scholar] [CrossRef] [Green Version]
- Shi, J.; Deng, G.; Kong, H.; Gu, C.; Ma, S.; Yin, X.; Zeng, X.; Cui, P.; Chen, Y.; Chen, H.; et al. H7N9 virulent mutants detected in chickens in China pose an increased threat to humans. Cell Res. 2017, 27, 1409–1421. [Google Scholar] [CrossRef] [Green Version]
- Zeng, X.; Tian, G.; Shi, J.; Deng, G.; Li, C.; Chen, H. Vaccination of poultry successfully eliminated human infection with H7N9 virus in China. Science China. Life Sci. 2018, 61, 1465–1473. [Google Scholar] [CrossRef]
- Chen, J.; Liu, Z.; Li, K.; Li, X.; Xu, L.; Zhang, M.; Wu, Y.; Liu, T.; Wang, X.; Xie, S.; et al. Emergence of novel avian origin H7N9 viruses after introduction of H7-Re3 and rLN79 vaccine strains to China. Transbound. Emerg. Dis. 2021. online ahead of print. [Google Scholar] [CrossRef]
- Becker, W.B. The isolation and classification of Tern virus: Influenza Virus A/Tern/South Africa/1961. J. Hyg. 1966, 64, 309–320. [Google Scholar] [CrossRef] [PubMed]
- Rowan, M.K. Mass mortality among European Common Terns in South Africa in April-May 1961. Br. Birds 1961, 55, 103–114. [Google Scholar]
- Uys, C.J.; Becker, W.B. Experimental infection of chickens with influenza A/Tern/South Africa/1961 and Chicken/Scotland/1959 viruses: II. Pathology. J. Comp. Pathol. 1967, 77, 167–173. [Google Scholar] [CrossRef]
- Wells, R.J.H. An outbreak of fowl plague in turkeys. Vet. Rec. 1963, 75, 783–786. [Google Scholar]
- Alexander, D.J.; Allan, W.H.; Parsons, G. Characterisation of influenza viruses isolated from turkeys in Great Britain during 1963–1977. Res Vet Sci. 1979, 26, 17–20. [Google Scholar] [CrossRef]
- Ping, J.; Selman, M.; Tyler, S.; Forbes, N.; Keleta, L.; Brown, E.G. Low-pathogenic avian influenza virus A/turkey/Ontario/6213/1966 (H5N1) is the progenitor of highly pathogenic A/turkey/Ontario/7732/1966 (H5N9). J. Gen. Virol. 2012, 93, 1649–1657. [Google Scholar] [CrossRef]
- Lang, G.; Narayan, O.; Rouse, B.T.; Ferguson, A.E.; Connell, M.C. A new influenza A virus infection in turkeys II. A highly pathogenic variant, a/turkey/ontario 772/66. Can. Vet. J. 1968, 9, 151–160. [Google Scholar]
- Lvov, D.K.; Shchelkanov, M.Y.; Alkhovsky, S.V.; Deryabin, P.G. Single-Stranded RNA Viruses. In Zoonotic Viruses in Northern Eurasia; Academic Press: Cambridge, MA, USA, 2015; pp. 135–392. [Google Scholar]
- Turner, A.J. The isolation of fowl plague virus in Victoria. Aust. Vet. J. 1976, 52, 384. [Google Scholar] [CrossRef]
- Westbury, H.A.; Turner, A.J.; Kovesdy, L. The pathogenicity of three Australian fowl plague viruses for chickens, turkeys and ducks. Vet. Microbiol. 1979, 4, 223–234. [Google Scholar] [CrossRef]
- Alexander, D.J.; Spackman, D. Characterisation of influenza A viruses isolated from turkeys in England during March-May 1979. Avian Pathol. 1981, 10, 281–293. [Google Scholar] [CrossRef] [Green Version]
- Röhm, C.; Süss, J.; Pohle, V.; Webster, R.G. Different hemagglutinin cleavage site variants of H7N7 in an influenza outbreak in chickens in Leipzig, Germany. Virology 1996, 218, 253–257. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kawaoka, Y.; Webster, R.G. Evolution of the A/Chicken/Pennsylvania/83 (H5N2) Influenza Virus. Virology 1985, 146, 130–137. [Google Scholar] [CrossRef]
- Bean, W.J.; Kawaoka, Y.; Wood, J.M.; Pearson, J.E.; Webster, R.G. Characterization of virulent and avirulent A/chicken/Pennsylvania/83 influenza A viruses: Potential role of defective interfering RNAs in nature. J. Virol. 1985, 54, 151–160. [Google Scholar] [CrossRef] [Green Version]
- Webster, R.G.; Kawaoka, Y.; Bean, W.J. Molecular changes in A/Chicken/Pennsylvania/83 (H5N2) influenza virus associated with acquisition of virulence. Virology 1986, 149, 165–173. [Google Scholar] [CrossRef]
- McNulty, M.S.; McParland, P.J.; Allan, G.M.; McCracken, R.M. Isolation of a highly pathogenic influenza virus from turkeys. Avian Pathol. 1985, 14, 173–176. [Google Scholar] [CrossRef]
- Barr, D.A.; Kelly, A.P.; Badman, R.T.; Campey, A.R.; O’Rourke, M.D.; Grix, D.C.; Reece, R.L. Avian influenza on a multi-age chicken farm. Aust. Vet. J. 1986, 63, 195–196. [Google Scholar] [CrossRef]
- Nestorowicz, A.; Kawaoka, Y.; Bean, W.J.; Webster, R.G. Molecular analysis of the hemagglutinin genes of Australian H7N7 influenza viruses: Role of passerine birds in maintenance or transmission? Virology 1987, 160, 411–418. [Google Scholar] [CrossRef]
- Alexander, D.J.; Lister, S.A.; Johnson, M.J.; Randall, C.J.; Thomas, P.J. An outbreak of highly pathogenic avian influenza in turkeys in Great Britain in 1991. Vet. Rec. 1993, 132, 535–536. [Google Scholar] [CrossRef]
- Selleck, P.W.; Gleeson, L.J.; Hooper, P.T.; Westbury, H.A.; Hansson, E. Identification and characterisation of an H7N3 influenza A virus from an outbreak of virulent avian influenza in Victoria. Aust. Vet. J. 1997, 75, 289–292. [Google Scholar] [CrossRef]
- Westbury, H.A. History of Highly Pathogenic Avian Influenza in Australia. Avian Dis. 2003, 47, 23–30. [Google Scholar]
- García, M.; Crawford, J.M.; Latimer, J.W.; Rivera-Cruz, E.; Perdue, M.L. Heterogeneity in the haemagglutinin gene and emergence of the highly pathogenic phenotype among recent H5N2 avian influenza viruses from Mexico. J. Gen. Virol. 1996, 77, 1493–1504. [Google Scholar] [CrossRef] [PubMed]
- Horimoto, T.; Rivera, E.; Pearson, J.; Senne, D.; Krauss, S.; Kawaoka, Y.; Webster, R.G. Origin and molecular changes associated with emergence of a highly pathogenic H5N2 influenza virus in Mexico. Virology 1995, 213, 223–230. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Swayne, D.E. Pathobiology of H5N2 Mexican avian influenza virus infections of chickens. Vet. Pathol. 1997, 34, 557–567. [Google Scholar] [CrossRef]
- Naeem, K.; Hussain, M. An outbreak of avian influenza in poultry in Pakistan. Vet. Rec. 1995, 137, 439. [Google Scholar] [CrossRef] [PubMed]
- Abbas, M.A.; Spackman, E.; Swayne, D.E.; Ahmed, Z.; Sarmento, L.; Siddique, N.; Naeem, K.; Hameed, A.; Rehmani, S. Sequence and phylogenetic analysis of H7N3 avian influenza viruses isolated from poultry in Pakistan 1995-2004. Virol. J. 2010, 7, 137. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, X.; Subbarao, K.; Cox, N.J.; Guo, Y. Genetic characterization of the pathogenic influenza A/Goose/Guangdong/1/96 (H5N1) virus: Similarity of its hemagglutinin gene to those of H5N1 viruses from the 1997 outbreaks in Hong Kong. Virology 1999, 261, 15–19. [Google Scholar] [CrossRef] [Green Version]
- Selleck, P.W.; Arzey, G.; Kirkland, P.D.; Reece, R.L.; Gould, A.R.; Daniels, P.W.; Westbury, H.A. An outbreak of highly pathogenic avian influenza in Australia in 1997 caused by an H7N4 virus. Avian Dis. 2003, 47, 806–811. [Google Scholar] [CrossRef]
- Capua, I.; Marangon, S.; Selli, L.; Alexander, D.J.; Swayne, D.E.; Dalla Pozza, M.; Parenti, E.; Cancellotti, F.M. Outbreaks of highly pathogenic avian influenza (H5N2) in Italy during October 1997 to January 1998. Avian Pathol. 1999, 28, 455–460. [Google Scholar] [CrossRef]
- Capua, I.; Mutinelli, F.; Pozza, M.D.; Donatelli, I.; Puzelli, S.; Cancellotti, F.M. The 1999–2000 avian influenza (H7N1) epidemic in Italy: Veterinary and human health implications. Acta Trop. 2002, 83, 7–11. [Google Scholar] [CrossRef]
- Monne, I.; Fusaro, A.; Nelson, M.I.; Bonfanti, L.; Mulatti, P.; Hughes, J.; Murcia, P.R.; Schivo, A.; Valastro, V.; Moreno, A.; et al. Emergence of a highly pathogenic avian influenza virus from a low-pathogenic progenitor. J. Virol. 2014, 88, 4375–4388. [Google Scholar] [CrossRef] [Green Version]
- Suarez, D.L.; Senne, D.A.; Banks, J.; Brown, I.H.; Essen, S.C.; Lee, C.W.; Manvell, R.J.; Mathieu-Benson, C.; Moreno, V.; Pedersen, J.C.; et al. Recombination Resulting in Virulence Shift in Avian Influenza Outbreak, Chile. Emerg. Infect. Dis. 2004, 10, 693–699. [Google Scholar] [CrossRef] [PubMed]
- Rojas, H.; Moreira, R.; Avalos, P.; Capua, I.; Marangon, S. Avian influenza in poultry in Chile. Vet. Rec. 2002, 151, 188. [Google Scholar] [PubMed]
- Jones, Y.L.; Swayne, D.E. Comparative pathobiology of low and high pathogenicity H7N3 Chilean avian influenza viruses in chickens. Avian Dis. 2004, 48, 119–128. [Google Scholar] [CrossRef] [PubMed]
- Elbers, A.R.W.; Fabri, T.H.F.; De Vries, T.S.; De Wit, J.J.; Pijpers, A.; Koch, G. The highly pathogenic avian influenza A (H7N7) virus epidemic in The Netherlands in 2003—Lessons learned from the first five outbreaks. Avian Dis. 2004, 48, 691–705. [Google Scholar] [CrossRef] [PubMed]
- Stegeman, A.; Bouma, A.; Elbers, A.R.W.; De Jong, M.C.M.; Nodelijk, G.; De Klerk, F.; Koch, G.; Van Boven, M. Avian influenza A virus (H7N7) epidemic in The Netherlands in 2003: Course of the epidemic and effectiveness of control measures. J. Infect. Dis. 2004, 190, 2088–2095. [Google Scholar] [CrossRef]
- Beerens, N.; Heutink, R.; Harders, F.; Bossers, A.; Koch, G.; Peeters, B. Emergence and Selection of a Highly Pathogenic Avian Influenza H7N3 Virus. J. Virol. 2020, 94, e01818-19. [Google Scholar] [CrossRef]
- Velkers, F.C.; Bouma, A.; Matthijs, M.G.R.; Koch, G.; Westendorp, S.T.; Stegeman, J.A. Outbreak of avian influenza H7N3 on a turkey farm in the Netherlands. Vet. Rec. 2006, 159, 403–405. [Google Scholar] [CrossRef] [Green Version]
- Naeem, K.; Siddique, N.; Ayaz, M.; Jalalee, M.A. Avian influenza in Pakistan: Outbreaks of low- and high-pathogenicity avian influenza in Pakistan during 2003–2006. Avian Dis. 2007, 51, 189–193. [Google Scholar] [CrossRef]
- Lee, C.-W.; Swayne, D.E.; Linares, J.A.; Senne, D.A.; Suarez, D.L. H5N2 Avian Influenza Outbreak in Texas in 2004: The First Highly Pathogenic Strain in the United States in 20 Years? J. Virol. 2005, 79, 11412–11421. [Google Scholar] [CrossRef] [Green Version]
- Hirst, M.; Astell, C.R.; Griffith, M.; Coughlin, S.M.; Moksa, M.; Zeng, T.; Smailus, D.E.; Holt, R.A.; Jones, S.; Marra, M.A.; et al. Novel avian influenza H7N3 strain outbreak, British Columbia. Emerg. Infect. Dis. 2004, 10, 2192–2195. [Google Scholar] [CrossRef]
- Abolnik, C.; Londt, B.Z.; Manvell, R.J.; Shell, W.; Banks, J.; Gerdes, G.H.; Akol, G.; Brown, I.H. Characterisation of a highly pathogenic influenza A virus of subtype H5N2 isolated from ostriches in South Africa in 2004. Influenza Other Respi. Viruses 2009, 3, 63–68. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alexander, D.J.; Brown, I.H. History of highly pathogenic avian influenza. Rev. Sci. Tech. 2009, 28, 19–38. [Google Scholar] [CrossRef] [PubMed]
- Abolnik, C. Molecular characterization of H5N2 avian influenza viruses isolated from South African ostriches in 2006. Avian Dis. 2007, 51, 873–879. [Google Scholar] [CrossRef] [PubMed]
- Berhane, Y.; Hisanaga, T.; Kehler, H.; Neufeld, J.; Manning, L.; Argue, C.; Handel, K.; Hooper-McGrevy, K.; Jonas, M.; Robinson, J.; et al. Highly pathogenic avian influenza virus A (H7N3) in domestic poultry, Saskatchewan, Canada, 2007. Emerg. Infect. Dis. 2009, 15, 1492–1495. [Google Scholar] [CrossRef]
- Gaidet, N.; Cattoli, G.; Hammoumi, S.; Newman, S.H.; Hagemeijer, W.; Takekawa, J.Y.; Cappelle, J.; Dodman, T.; Joannis, T.; Gil, P.; et al. Evidence of infection by H5N2 highly pathogenic avian influenza viruses in healthy wild waterfowl. PLoS Pathog. 2008, 4, e1000127. [Google Scholar] [CrossRef] [Green Version]
- Seekings, A.H.; Slomka, M.J.; Russell, C.; Howard, W.A.; Choudhury, B.; Nuñéz, A.; Löndt, B.Z.; Cox, W.; Ceeraz, V.; Thorén, P.; et al. Direct evidence of H7N7 avian influenza virus mutation from low to high virulence on a single poultry premises during an outbreak in free range chickens in the UK, 2008. Infect. Genet. Evol. 2018, 64, 13–31. [Google Scholar] [CrossRef]
- Iglesias, I.; Martínez, M.; Muñoz, M.J.; De La Torre, A.; Sánchez-Vizcaíno, J.M. First case of highly pathogenic avian influenza in poultry in Spain. Transbound. Emerg. Dis. 2010, 57, 282–285. [Google Scholar] [CrossRef]
- Abolnik, C.; Olivier, A.J.; Grewar, J.; Gers, S.; Romito, M. Molecular analysis of the 2011 HPAI H5N2 outbreak in ostriches, South Africa. Avian Dis. 2012, 56, 865–879. [Google Scholar] [CrossRef]
- Office International des Épizooties. Report Id. FUR-12739, Australia. 2012. Available online: https://wahis.oie.int/#/report-info?reportId=4656 (accessed on 27 March 2022).
- Lee, C.D.; Zhu, H.; Huang, P.; Peng, L.; Chang, Y.; Yip, C.; Li, Y.; Cheung, C.; Compans, R.; Yang, C.; et al. Emergence and Evolution of Avian H5N2 Influenza Viruses in Chickens in Taiwan. J. Virol. 2014, 88, 5677. [Google Scholar] [CrossRef] [Green Version]
- Maurer-Stroh, S.; Lee, R.T.; Gunalan, V.; Eisenhaber, F. The highly pathogenic H7N3 avian influenza strain from July 2012 in Mexico acquired an extended cleavage site through recombination with host 28S rRNA. Virol. J. 2013, 10, 139. [Google Scholar] [CrossRef] [Green Version]
- Kapczynski, D.R.; Pantin-Jackwood, M.; Guzman, S.G.; Ricardez, Y.; Spackman, E.; Bertran, K.; Suarez, D.L.; Swayne, D.E. Characterization of the 2012 Highly Pathogenic Avian Influenza H7N3 Virus Isolated from Poultry in an Outbreak in Mexico: Pathobiology and Vaccine Protection. J. Virol. 2013, 87, 9086–9096. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bonfanti, L.; Monne, I.; Tamba, M.; Santucci, U.; Massi, P.; Patregnani, T.; Loli Piccolomini, L.; Natalini, S.; Ferri, G.; Cattoli, G.; et al. Highly pathogenic H7N7 avian influenza in Italy. Vet. Rec. 2014, 174, 382. [Google Scholar] [CrossRef] [PubMed]
- Fusaro, A.; Tassoni, L.; Milani, A.; Hughes, J.; Salviato, A.; Murcia, P.R.; Massi, P.; Zamperin, G.; Bonfanti, L.; Marangon, S.; et al. Unexpected Interfarm Transmission Dynamics during a Highly Pathogenic Avian Influenza Epidemic. J. Virol. 2016, 90, 6401–6411. [Google Scholar] [CrossRef] [Green Version]
- Office International des Épizooties. Rerport Id. FUR_14811, Australia. 2013. Available online: https://wahis.oie.int/#/report-info?reportId=5343 (accessed on 27 March 2022).
- Byrne, A.M.P.; Reid, S.M.; Seekings, A.H.; Núñez, A.; Prieto, A.B.O.; Ridout, S.; Warren, C.J.; Puranik, A.; Ceeraz, V.; Essen, S.; et al. H7N7 avian influenza virus mutation from low to high pathogenicity on a layer chicken farm in the uk. Viruses 2021, 13, 259. [Google Scholar] [CrossRef]
- Dietze, K.; Graaf, A.; Homeier-Bachmann, T.; Grund, C.; Forth, L.; Pohlmann, A.; Jeske, C.; Wintermann, M.; Beer, M.; Conraths, F.J.; et al. From low to high pathogenicity—Characterization of H7N7 avian influenza viruses in two epidemiologically linked outbreaks. Transbound. Emerg. Dis. 2018, 65, 1576–1587. [Google Scholar] [CrossRef] [PubMed]
- Briand, F.X.; Niqueux, E.; Schmitz, A.; Hirchaud, E.; Quenault, H.; Allée, C.; Le Prioux, A.; Guillou-Cloarec, C.; Ogor, K.; Le Bras, M.O.; et al. Emergence and multiple reassortments of French 2015-2016 highly pathogenic H5 avian influenza viruses. Infect. Genet. Evol. 2018, 61, 208–214. [Google Scholar] [CrossRef] [PubMed]
- Briand, F.X.; Schmitz, A.; Ogor, K.; Le Prioux, A.; Guillou-Cloarec, C.; Guillemoto, C.; Allée, C.; Le Bras, M.; Hirchaud, E.; Quenault, H.; et al. Emerging highly pathogenic H5 avian influenza viruses in France during winter 2015/16: Phylogenetic analyses and markers for zoonotic potential. Euro Surveill. 2017, 22, 30473. [Google Scholar] [CrossRef] [Green Version]
- Office International des Épizooties. Report Id. FUR_22420, Algeria. 2016. Available online: https://wahis.oie.int/#/report-info?reportId=8691 (accessed on 27 March 2022).
- Pantin-Jackwood, M.J.; Stephens, C.B.; Bertran, K.; Swayne, D.E.; Spackman, E. The pathogenesis of H7N8 low and highly pathogenic avian influenza viruses from the United States 2016 outbreak in chickens, turkeys and mallards. PLoS ONE 2017, 12, e0177265. [Google Scholar] [CrossRef]
- Killian, M.L.; Kim-Torchetti, M.; Hines, N.; Yingst, S.; DeLiberto, T.; Lee, D.H. Outbreak of H7N8 Low Pathogenic Avian Influenza in Commercial Turkeys with Spontaneous Mutation to Highly Pathogenic Avian Influenza. Genome Announc. 2016, 4, e00457-16. [Google Scholar] [CrossRef] [Green Version]
- Mulatti, P.; Zecchin, B.; Monne, I.; Vieira, J.T.; Dorotea, T.; Terregino, C.; Lorenzetto, M.; Piccolomini, L.L.; Santi, A.; Massi, P.; et al. H7N7 Highly Pathogenic Avian Influenza in Poultry Farms in Italy in 2016. Avian Dis. 2017, 61, 261–266. [Google Scholar] [CrossRef]
- Lee, D.H.; Torchetti, M.K.; Killian, M.L.; Berhane, Y.; Swayne, D.E. Highly pathogenic avian influenza A(H7N9) virus, Tennessee, USA, March 2017. Emerg. Infect. Dis. 2017, 23, 1860–1863. [Google Scholar] [CrossRef] [PubMed]
- Youk, S.; Lee, D.H.; Killian, M.L.; Pantin-Jackwood, M.J.; Swayne, D.E.; Torchetti, M.K. Highly Pathogenic Avian Influenza A(H7N3) Virus in Poultry, United States, 2020. Emerg. Infect. Dis. 2020, 26, 2966–2969. [Google Scholar] [CrossRef] [PubMed]
- Office International des Épizooties. Report Id. FUR_37191, Australia. 2020. Available online: https://wahis.oie.int/#/report-info?reportId=16548 (accessed on 27 March 2022).
- Office International des Épizooties. Report Id. FUR_150999, Lithuania. 2021. Available online: https://wahis.oie.int/#/report-info?reportId=37130 (accessed on 27 March 2022).
- Morgan, I.R.; Kelly, A.P. Epidemiology of an avian influenza outbreak in Victoria in 1985. Aust. Vet. J. 1990, 67, 125–128. [Google Scholar] [CrossRef] [PubMed]
- Philippa, J.D.W.; Munster, V.J.; Bolhuis, H.V.; Bestebroer, T.M.; Schaftenaar, W.; Beyer, W.E.P.; Fouchier, R.A.M.; Kuiken, T.; Osterhaus, A.D.M.E. Highly pathogenic avian influenza (H7N7): Vaccination of zoo birds and transmission to non-poultry species. Vaccine 2005, 23, 5743–5750. [Google Scholar] [CrossRef] [PubMed]
- Verhagen, J.H.; Fouchier, R.A.M.; Lewis, N. Highly pathogenic avian influenza viruses at the wild–domestic bird interface in europe: Future directions for research and surveillance. Viruses 2021, 13, 212. [Google Scholar] [CrossRef]
- Chen, H.; Smith, G.J.D.; Zhang, S.Y.; Qin, K.; Wang, J.; Li, K.S.; Webster, R.G.; Peiris, J.S.M.; Guan, Y. H5N1 virus outbreak in migratory waterfowl. Nature 2005, 436, 191–192. [Google Scholar] [CrossRef]
- Office International des Épizooties. Report Id. CAN-2021-HPAI-001, Canada. 2021. Available online: https://wahis.oie.int/#/report-info?reportId=45054 (accessed on 21 May 2022).
- Wiley, D.C.; Skehel, J.J. The structure and function of the hemagglutinin membrane glycoprotein of influenza virus. Annu. Rev. Biochem. 1987, 56, 365–394. [Google Scholar] [CrossRef]
- Chen, J.; Lee, K.H.; Steinhauer, D.A.; Stevens, D.J.; Skehel, J.J.; Wiley, D.C. Structure of the hemagglutinin precursor cleavage site, a determinant of influenza pathogenicity and the origin of the labile conformation. Cell 1998, 95, 409–417. [Google Scholar] [CrossRef] [Green Version]
- Klenk, H.-D.; Rott, R.; Orlich, M.; Blödorn, J. Activation of influenza A viruses by trypsin treatment. Virology 1975, 68, 426–439. [Google Scholar] [CrossRef]
- Maeda, T.; Ohnishi, S. Activation of influenza virus by acidic media causes hemolysis and fusion of erythrocytes. FEBS Lett. 1980, 122, 283–287. [Google Scholar] [CrossRef] [Green Version]
- Skehel, J.J.; Bayley, P.M.; Brown, E.B.; Martin, S.R.; Waterfield, M.D.; White, J.M.; Wilson, I.A.; Wiley, D.C. Changes in the conformation of influenza virus hemagglutinin at the pH optimum of virus-mediated membrane fusion. Proc. Natl. Acad. Sci. USA 1982, 79, 968–972. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stieneke-Gröber, A.; Vey, M.; Angliker, H.; Shaw, E.; Thomas, G.; Roberts, C.; Klenk, H.D.; Garten, W. Influenza virus hemagglutinin with multibasic cleavage site is activated by furin, a subtilisin-like endoprotease. EMBO J. 1992, 11, 2407–2414. [Google Scholar] [CrossRef] [PubMed]
- Bosch, F.X.; Garten, W.; Klenk, H.D.; Rott, R. Proteolytic cleavage of influenza virus hemagglutinins: Primary structure of the connecting peptide between HA1 and HA2 determines proteolytic cleavability and pathogenicity of Avian influenza viruses. Virology 1981, 113, 725–735. [Google Scholar] [CrossRef]
- Horimoto, T.; Kawaoka, Y. Reverse genetics provides direct evidence for a correlation of hemagglutinin cleavability and virulence of an avian influenza A virus. J. Virol. 1994, 68, 3120–3128. [Google Scholar] [CrossRef] [Green Version]
- Bogs, J.; Veits, J.; Gohrbandt, S.; Hundt, J.; Stech, O.; Breithaupt, A.; Teifke, J.P.; Mettenleiter, T.C.; Stech, J. Highly pathogenic H5N1 influenza viruses carry virulence determinants beyond the polybasic hemagglutinin cleavage site. PLoS ONE 2010, 5, 11826. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abdelwhab, E.S.M.; Veits, J.; Mettenleiter, T.C. Genetic changes that accompanied shifts of low pathogenic avian influenza viruses toward higher pathogenicity in poultry. Virulence 2013, 4, 441–452. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Richard, M.; Fouchier, R.; Monne, I.; Kuiken, T. Mechanisms and risk factors for mutation from low to highly pathogenic avian influenza virus. EFSA Support. Publ. 2017, 14, 1287E. [Google Scholar] [CrossRef] [Green Version]
- Escalera-Zamudio, M.; Golden, M.; Gutiérrez, B.; Thézé, J.; Keown, J.R.; Carrique, L.; Bowden, T.A.; Pybus, O.G. Parallel evolution in the emergence of highly pathogenic avian influenza A viruses. Nat. Commun. 2020, 11, 5511. [Google Scholar] [CrossRef]
- Li, J.; Cardona, C.J. Adaptation and Transmission of a Wild Duck Avian Influenza Isolate in Chickens. Avian Dis. 2010, 54, 586–590. [Google Scholar] [CrossRef]
- Matrosovich, M.; Zhou, N.; Kawaoka, Y.; Webster, R. The Surface Glycoproteins of H5 Influenza Viruses Isolated from Humans, Chickens, and Wild Aquatic Birds Have Distinguishable Properties. J. Virol. 1999, 73, 1146. [Google Scholar] [CrossRef] [Green Version]
- Hoffmann, T.W.; Munier, S.; Larcher, T.; Soubieux, D.; Ledevin, M.; Esnault, E.; Tourdes, A.; Croville, G.; Guérin, J.-L.; Quéré, P.; et al. Length Variations in the NA Stalk of an H7N1 Influenza Virus Have Opposite Effects on Viral Excretion in Chickens and Ducks. J. Virol. 2012, 86, 584–588. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sorrell, E.M.; Song, H.; Pena, L.; Perez, D.R. A 27-Amino-Acid Deletion in the Neuraminidase Stalk Supports Replication of an Avian H2N2 Influenza A Virus in the Respiratory Tract of Chickens. J. Virol. 2010, 84, 11831–11840. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, H.; Yu, Z.; Hu, Y.; Tu, J.; Zou, W.; Peng, Y.; Zhu, J.; Li, Y.; Zhang, A.; Yu, Z.; et al. The special neuraminidase stalk-motif responsible for increased virulence and pathogenesis of H5N1 influenza A virus. PLoS ONE 2009, 4, e6277. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Dohna, H.Z.; Cardona, C.J.; Miller, J.; Carpenter, T.E. Emergence and Genetic Variation of Neuraminidase Stalk Deletions in Avian Influenza Viruses. PLoS ONE 2011, 6, e14722. [Google Scholar] [CrossRef]
- Dundon, W.G.; Milani, A.; Cattoli, G.; Capua, I. Progressive truncation of the Non-Structural 1 gene of H7N1 avian influenza viruses following extensive circulation in poultry. Virus Res. 2006, 119, 171–176. [Google Scholar] [CrossRef]
- Abdelwhab, E.S.M.; Veits, J.; Breithaupt, A.; Gohrbandt, S.; Ziller, M.; Teifke, J.P.; Stech, J.; Mettenleiter, T.C. Prevalence of the C-terminal truncations of NS1 in avian influenza A viruses and effect on virulence and replication of a highly pathogenic H7N1 virus in chickens. Virulence 2016, 7, 546–557. [Google Scholar] [CrossRef] [Green Version]
- Abdelwhab, E.S.M.; Veits, J.; Tauscher, K.; Ziller, M.; Grund, C.; Hassan, M.K.; Shaheen, M.; Harder, T.C.; Teifke, J.; Stech, J.; et al. Progressive glycosylation of the haemagglutinin of avian influenza H5N1 modulates virus replication, virulence and chicken-to-chicken transmission without significant impact on antigenic drift. J. Gen. Virol. 2016, 97, 3193–3204. [Google Scholar] [CrossRef]
- Sealy, J.E.; Howard, W.A.; Molesti, E.; Iqbal, M.; Temperton, N.J.; Banks, J.; Slomka, M.J.; Barclay, W.S.; Long, J.S. Amino acid substitutions in the H5N1 avian influenza haemagglutinin alter pH of fusion and receptor binding to promote a highly pathogenic phenotype in chickens. J. Gen. Virol. 2021, 102, 001672. [Google Scholar] [CrossRef]
- DuBois, R.M.; Zaraket, H.; Reddivari, M.; Heath, R.J.; White, S.W.; Russell, C.J. Acid Stability of the Hemagglutinin Protein Regulates H5N1 Influenza Virus Pathogenicity. PLOS Pathog. 2011, 7, e1002398. [Google Scholar] [CrossRef] [Green Version]
- Reed, M.L.; Bridges, O.A.; Seiler, P.; Kim, J.-K.; Yen, H.-L.; Salomon, R.; Govorkova, E.A.; Webster, R.G.; Russell, C.J. The pH of Activation of the Hemagglutinin Protein Regulates H5N1 Influenza Virus Pathogenicity and Transmissibility in Ducks. J. Virol. 2010, 84, 1527–1535. [Google Scholar] [CrossRef] [Green Version]
- Garten, W.; Braden, C.; Arendt, A.; Peitsch, C.; Baron, J.; Lu, Y.; Pawletko, K.; Hardes, K.; Steinmetzer, T.; Böttcher-Friebertshäuser, E. Influenza virus activating host proteases: Identification, localization and inhibitors as potential therapeutics. Eur. J. Cell Biol. 2015, 94, 375–383. [Google Scholar] [CrossRef] [PubMed]
- Galloway, S.E.; Reed, M.L.; Russell, C.J.; Steinhauer, D.A. Influenza HA Subtypes Demonstrate Divergent Phenotypes for Cleavage Activation and pH of Fusion: Implications for Host Range and Adaptation. PLoS Pathog. 2013, 9, e1003151. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hatesuer, B.; Bertram, S.; Mehnert, N.; Bahgat, M.M.; Nelson, P.S.; Pöhlman, S.; Schughart, K. Tmprss2 Is Essential for Influenza H1N1 Virus Pathogenesis in Mice. PLoS Pathog. 2013, 9, e1003774. [Google Scholar] [CrossRef] [Green Version]
- Tarnow, C.; Engels, G.; Arendt, A.; Schwalm, F.; Sediri, H.; Preuss, A.; Nelson, P.S.; Garten, W.; Klenk, H.D.; Gabriel, G.; et al. TMPRSS2 Is a Host Factor That Is Essential for Pneumotropism and Pathogenicity of H7N9 Influenza A Virus in Mice. J. Virol. 2014, 88, 4744–4751. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sakai, K.; Ami, Y.; Tahara, M.; Kubota, T.; Anraku, M.; Abe, M.; Nakajima, N.; Sekizuka, T.; Shirato, K.; Suzaki, Y.; et al. The Host Protease TMPRSS2 Plays a Major Role in In Vivo Replication of Emerging H7N9 and Seasonal Influenza Viruses. J. Virol. 2014, 88, 5608–5616. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lambertz, R.L.O.; Gerhauser, I.; Nehlmeier, I.; Leist, S.R.; Kollmus, H.; Pöhlmann, S.; Schughart, K. Tmprss2 knock-out mice are resistant to H10 influenza a virus pathogenesis. J. Gen. Virol. 2019, 100, 1073–1078. [Google Scholar] [CrossRef]
- Lambertz, R.L.O.; Gerhauser, I.; Nehlmeier, I.; Gärtner, S.; Winkler, M.; Leist, S.R.; Kollmus, H.; Pöhlmann, S.; Schughart, K. H2 influenza A virus is not pathogenic in Tmprss2 knock-out mice. Virol. J. 2020, 17, 56. [Google Scholar] [CrossRef]
- Limburg, H.; Harbig, A.; Bestle, D.; Stein, D.A.; Moulton, H.M.; Jaeger, J.; Janga, H.; Hardes, K.; Koepke, J.; Schulte, L.; et al. TMPRSS2 Is the Major Activating Protease of Influenza A Virus in Primary Human Airway Cells and Influenza B Virus in Human Type II Pneumocytes. J. Virol. 2019, 93, e00649-19. [Google Scholar] [CrossRef] [Green Version]
- Bestle, D.; Limburg, H.; Kruhl, D.; Harbig, A.; Stein, D.A.; Moulton, H.; Matrosovich, M.; Abdelwhab, E.M.; Stech, J.; Böttcher-Friebertshäuser, E. Hemagglutinins of Avian Influenza Viruses Are Proteolytically Activated by TMPRSS2 in Human and Murine Airway Cells. J. Virol. 2021, 95, e0090621. [Google Scholar] [CrossRef]
- Böttcher-Friebertshäuser, E.; Freuer, C.; Sielaff, F.; Schmidt, S.; Eickmann, M.; Uhlendorff, J.; Steinmetzer, T.; Klenk, H.-D.; Garten, W. Cleavage of Influenza Virus Hemagglutinin by Airway Proteases TMPRSS2 and HAT Differs in Subcellular Localization and Susceptibility to Protease Inhibitors. J. Virol. 2010, 84, 5605–5614. [Google Scholar] [CrossRef] [Green Version]
- Wysocka, M.; Spichalska, B.; Lesner, A.; Jaros, M.; Brzozowski, K.; Łgowska, A.; Rolka, K. Substrate specificity and inhibitory study of human airway trypsin-like protease. Bioorg. Med. Chem. 2010, 18, 5504–5509. [Google Scholar] [CrossRef] [PubMed]
- Baron, J.; Tarnow, C.; Mayoli-Nüssle, D.; Schilling, E.; Meyer, D.; Hammami, M.; Schwalm, F.; Steinmetzer, T.; Guan, Y.; Garten, W.; et al. Matriptase, HAT, and TMPRSS2 Activate the Hemagglutinin of H9N2 Influenza A Viruses. J. Virol. 2013, 87, 1811–1820. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Beaulieu, A.; Gravel, É.; Cloutier, A.; Marois, I.; Colombo, É.; Désilets, A.; Verreault, C.; Leduc, R.; Marsault, É.; Richter, M.V. Matriptase Proteolytically Activates Influenza Virus and Promotes Multicycle Replication in the Human Airway Epithelium. J. Virol. 2013, 87, 4237–4251. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hamilton, B.S.; Gludish, D.W.J.; Whittaker, G.R. Cleavage Activation of the Human-Adapted Influenza Virus Subtypes by Matriptase Reveals both Subtype and Strain Specificities. J. Virol. 2012, 86, 10579–10586. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bertram, S.; Glowacka, I.; Blazejewska, P.; Soilleux, E.; Allen, P.; Danisch, S.; Steffen, I.; Choi, S.-Y.; Park, Y.; Schneider, H.; et al. TMPRSS2 and TMPRSS4 Facilitate Trypsin-Independent Spread of Influenza Virus in Caco-2 Cells. J. Virol. 2010, 84, 10016–10025. [Google Scholar] [CrossRef] [Green Version]
- Kido, H.; Yokogoshi, Y.; Sakai, K.; Tashiro, M.; Kishino, Y.; Fukutomi, A.; Katunuma, N. Isolation and characterization of a novel trypsin-like protease found in rat bronchiolar epithelial Clara cells. A possible activator of the viral fusion glycoprotein. J. Biol. Chem. 1992, 267, 13573–13579. [Google Scholar] [CrossRef]
- Murakami, M.; Towatari, T.; Ohuchi, M.; Shiota, M.; Akao, M.; Okumura, Y.; Parry, M.A.A.; Kido, H. Mini-plasmin found in the epithelial cells of bronchioles triggers infection by broad-spectrum influenza A viruses and Sendai virus. Eur. J. Biochem. 2001, 268, 2847–2855. [Google Scholar] [CrossRef] [PubMed]
- Gotoh, B.; Ogasawara, T.; Toyoda, T.; Inocencio, N.M.; Hamaguchi, M.; Nagai, Y. An endoprotease homologous to the blood clotting factor X as a determinant of viral tropism in chick embryo. EMBO J. 1990, 9, 4189–4195. [Google Scholar] [CrossRef]
- Lazarowitz, S.G.; Goldberg, A.R.; Choppin, P.W. Proteolytic cleavage by plasmin of the HA polypeptide of influenza virus: Host cell activation of serum plasminogen. Virology 1973, 56, 172–180. [Google Scholar] [CrossRef]
- Goto, H.; Kawaoka, Y. A novel mechanism for the acquisition of virulence by a human influenza A virus. Proc. Natl. Acad. Sci. USA 1998, 95, 10224–10228. [Google Scholar] [CrossRef] [Green Version]
- Kwon, H.I.; Kim, Y.I.; Park, S.J.; Kim, E.H.; Kim, S.; Si, Y.J.; Song, M.S.; Pascua, P.N.Q.; Govorkova, E.A.; Webster, R.G.; et al. A novel neuraminidase-dependent hemagglutinin cleavage mechanism enables the systemic spread of an H7N6 avian influenza virus. MBio 2019, 10, e02369-19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Garten, W. Characterization of Proprotein Convertases and Their Involvement in Virus Propagation. In Activation of Viruses by Host Proteases; Böttcher-Friebertshäuser, E., Garten, W., Klenk, H., Eds.; Springer: Cham, Switzerland, 2018; pp. 205–248. [Google Scholar]
- Shang, J.; Wan, Y.; Luo, C.; Ye, G.; Geng, Q.; Auerbach, A.; Li, F. Cell entry mechanisms of SARS-CoV-2. Proc. Natl. Acad. Sci. USA 2020, 117, 11727–11734. [Google Scholar] [CrossRef]
- Horimoto, T.; Nakayama, K.; Smeekens, S.P.; Kawaoka, Y. Proprotein-processing endoproteases PC6 and furin both activate hemagglutinin of virulent avian influenza viruses. J. Virol. 1994, 68, 6074–6078. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Basak, A.; Zhong, M.; Munzer, J.S.; Chrétien, M.; Seidah, N.G. Implication of the proprotein convertases furin, PC5 and PC7 in the cleavage of surface glycoproteins of Hong Kong, Ebola and respiratory syncytial viruses: A comparative analysis with fluorogenic peptides. Biochem. J. 2001, 353, 537–545. [Google Scholar] [CrossRef]
- Klenk, H.D.; Wöllert, W.; Rott, R.; Scholtissek, C. Association of influenza virus proteins with cytoplasmic fractions. Virology 1974, 57, 28–41. [Google Scholar] [CrossRef]
- Vey, M.; Orlich, M.; Adler, S.; Klenk, H.D.; Rott, R.; Garten, W. Hemagglutinin activation of pathogenic avian influenza viruses of serotype H7 requires the protease recognition motif R-X-K/R-R. Virology 1992, 188, 408–413. [Google Scholar] [CrossRef]
- Okumura, Y.; Takahashi, E.; Yano, M.; Ohuchi, M.; Daidoji, T.; Nakaya, T.; Böttcher, E.; Garten, W.; Klenk, H.-D.; Kido, H. Novel Type II Transmembrane Serine Proteases, MSPL and TMPRSS13, Proteolytically Activate Membrane Fusion Activity of the Hemagglutinin of Highly Pathogenic Avian Influenza Viruses and Induce Their Multicycle Replication. J. Virol. 2010, 84, 5089–5096. [Google Scholar] [CrossRef] [Green Version]
- Feldmann, A.; Schäfer, M.K.-H.; Garten, W.; Klenk, H.-D. Targeted Infection of Endothelial Cells by Avian Influenza Virus A/FPV/Rostock/34 (H7N1) in Chicken Embryos. J. Virol. 2000, 74, 8018–8027. [Google Scholar] [CrossRef] [Green Version]
- Luczo, J.M.; Stambas, J.; Durr, P.A.; Michalski, W.P.; Bingham, J. Molecular pathogenesis of H5 highly pathogenic avian influenza: The role of the haemagglutinin cleavage site motif. Rev. Med. Virol. 2015, 25, 406–430. [Google Scholar] [CrossRef] [Green Version]
- Duckert, P.; Brunak, S.; Blom, N. Prediction of proprotein convertase cleavage sites. Protein Eng. Des. Sel. 2004, 17, 107–112. [Google Scholar] [CrossRef] [Green Version]
- Tian, S. A 20 Residues Motif Delineates the Furin Cleavage Site and its Physical Properties May Influence Viral Fusion. Biochem. Insights 2009, 2, 9–20. [Google Scholar] [CrossRef] [Green Version]
- Kawaoka, Y.; Webster, R.G. Sequence requirements for cleavage activation of influenza virus hemagglutinin expressed in mammalian cells. Proc. Natl. Acad. Sci. USA 1988, 85, 324–328. [Google Scholar] [CrossRef] [Green Version]
- Orlich, M.; Gottwald, H.; Rott, R. Nonhomologous recombination between the hemagglutinin gene and the nucleoprotein gene of an influenza virus. Virology 1994, 204, 462–465. [Google Scholar] [CrossRef] [PubMed]
- Luczo, J.M.; Tachedjian, M.; Harper, J.A.; Payne, J.S.; Butler, J.M.; Sapats, S.I.; Lowther, S.L.; Michalski, W.P.; Stambas, J.; Bingham, J. Evolution of high pathogenicity of H5 avian influenza virus: Haemagglutinin cleavage site selection of reverse-genetics mutants during passage in chickens. Sci. Rep. 2018, 8, 11518. [Google Scholar] [CrossRef]
- Lee, C.W.; Lee, Y.J.; Senne, D.A.; Suarez, D.L. Pathogenic potential of North American H7N2 avian influenza virus: A mutagenesis study using reverse genetics. Virology 2006, 353, 388–395. [Google Scholar] [CrossRef] [Green Version]
- Morsy, J.; Garten, W.; Rott, R. Activation of an influenza virus A/turkey/Oregon/71 HA insertion variant by the subtilisin-like endoprotease furin. Virology 1994, 202, 988–991. [Google Scholar] [CrossRef] [PubMed]
- Hamilton, B.S.; Sun, X.; Chung, C.; Whittaker, G.R. Acquisition of a novel eleven amino acid insertion directly N-terminal to a tetrabasic cleavage site confers intracellular cleavage of an H7N7 influenza virus hemagglutinin. Virology 2012, 434, 88–95. [Google Scholar] [CrossRef] [Green Version]
- Londt, B.Z.; Banks, J.; Alexander, D.J. Highly pathogenic avian influenza viruses with low virulence for chickens in in vivo tests. Avian Pathol. 2007, 36, 347–350. [Google Scholar] [CrossRef] [Green Version]
- Kawaoka, Y.; Naeve, C.W.; Webster, R.G. Is virulence of H5N2 influenza viruses in chickens associated with loss of carbohydrate from the hemagglutinin? Virology 1984, 139, 303–316. [Google Scholar] [CrossRef]
- Kawaoka, Y.; Webster, R.G. Interplay between carbohydrate in the stalk and the length of the connecting peptide determines the cleavability of influenza virus hemagglutinin. J. Virol. 1989, 63, 3296–3300. [Google Scholar] [CrossRef] [Green Version]
- Ohuchi, M.; Orlich, M.; Ohuchi, R.; Simpson, B.E.J.; Garten, W.; Klenk, H.D.; Rott, R. Mutations at the cleavage site of the hemagglutinin alter the pathogenicity of influenza virus a/chick/penn/83 (H5N2). Virology 1989, 168, 274–280. [Google Scholar] [CrossRef]
- Soda, K.; Asakura, S.; Okamatsu, M.; Sakoda, Y.; Kida, H. H9N2 influenza virus acquires intravenous pathogenicity on the introduction of a pair of di-basic amino acid residues at the cleavage site of the hemagglutinin and consecutive passages in chickens. Virol. J. 2011, 8, 64. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cheng, M.-C.; Soda, K.; Lee, M.-S.; Lee, S.-H.; Sakoda, Y.; Kida, H.; Wang, C.-H. Isolation and Characterization of Potentially Pathogenic H5N2 Influenza Virus from a Chicken in Taiwan in 2008. Source Avian Dis. 2010, 54, 885–893. [Google Scholar] [CrossRef] [PubMed]
- Soda, K.; Cheng, M.C.; Yoshida, H.; Endo, M.; Lee, S.H.; Okamatsu, M.; Sakoda, Y.; Wang, C.H.; Kida, H. A low pathogenic H5N2 influenza virus isolated in Taiwan acquired high Pathogenicity by consecutive passages in chickens. J. Vet. Med. Sci. 2011, 73, 767–772. [Google Scholar] [CrossRef] [Green Version]
- Brown, I.; Abolnik, C.; Garcia-Garcia, J.; McCullough, S.; Swayne, D.E.; Cattoli, G. High-pathogenicity avian influenza outbreaks since 2008, excluding multi-continental panzootic of H5 Goose/Guangdong-lineage viruses. In Animal Influenza; Swayne, D.E., Ed.; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 2016; pp. 248–270. [Google Scholar]
- Gischke, M.; Ulrich, R.; Fatola, O.I.; Scheibner, D.; Salaheldin, A.H.; Crossley, B.; Böttcher-Friebertshäuser, E.; Veits, J.; Mettenleiter, T.C.; Abdelwhab, E.M. Insertion of basic amino acids in the hemagglutinin cleavage site of H4N2 avian influenza virus (AIV)—reduced virus fitness in chickens is restored by reassortment with highly pathogenic H5N1 AIV. Int. J. Mol. Sci. 2020, 21, 2353. [Google Scholar] [CrossRef] [Green Version]
- Sun, Y.; Sun, S.; Ma, J.; Tan, Y.; Du, L.; Shen, Y.; Mu, Q.; Pu, J.; Lin, D.; Liu, J. Identification and characterization of avian-origin H3N2 canine influenza viruses in northern China during 2009–2010. Virology 2013, 435, 301–307. [Google Scholar] [CrossRef] [Green Version]
- Tse, L.V.; Hamilton, A.M.; Friling, T.; Whittaker, G.R. A Novel Activation Mechanism of Avian Influenza Virus H9N2 by Furin. J. Virol. 2014, 88, 1673–1683. [Google Scholar] [CrossRef] [Green Version]
- Zhang, J.; Ma, K.; Li, B.; Chen, Y.; Qiu, Z.; Xing, J.; Huang, J.; Hu, C.; Huang, Y.; Li, H.; et al. A risk marker of tribasic hemagglutinin cleavage site in influenza A (H9N2) virus. Commun. Biol. 2021, 4, 71. [Google Scholar] [CrossRef] [PubMed]
- Parvin, R.; Schinkoethe, J.; Grund, C.; Ulrich, R.; Bönte, F.; Behr, K.P.; Voss, M.; Samad, M.A.; Hassan, K.E.; Luttermann, C.; et al. Comparison of pathogenicity of subtype H9 avian influenza wild-type viruses from a wide geographic origin expressing mono-, di-, or tri-basic hemagglutinin cleavage sites. Vet. Res. 2020, 51, 48. [Google Scholar] [CrossRef] [Green Version]
- Gischke, M.; Bagato, O.; Breithaupt, A.; Scheibner, D.; Blaurock, C.; Vallbracht, M.; Karger, A.; Crossley, B.; Veits, J.; Böttcher-Friebertshäuser, E.; et al. The role of glycosylation in the N-terminus of the hemagglutinin of a unique H4N2 with a natural polybasic cleavage site in virus fitness in vitro and in vivo. Virulence 2021, 12, 666–678. [Google Scholar] [CrossRef]
- Funk, M.; de Bruin, A.C.M.; Spronken, M.I.; Gultyaev, A.P.; Richard, M. In silico analyses of the role of codon usage in H5 and H7 hemagglutinins in highly pathogenic avian influenza genesis. Viruses 2022, 14, 1352. [Google Scholar] [CrossRef]
- Gu, M.; Xu, L.; Wang, X.; Liu, X. Current situation of H9N2 subtype avian influenza in China. Vet. Res. 2017, 48, 1–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Veits, J.; Weber, S.; Stech, O.; Breithaupt, A.; Gräber, M.; Gohrbandt, S.; Bogs, J.; Hundt, J.; Teifke, J.P.; Mettenleiter, T.C.; et al. Avian influenza virus hemagglutinins H2, H4, H8, and H14 support a highly pathogenic phenotype. Proc. Natl. Acad. Sci. USA 2012, 109, 2579–2584. [Google Scholar] [CrossRef] [Green Version]
- Gohrbandt, S.; Veits, J.; Breithaupt, A.; Hundt, J.; Teifke, J.P.; Stech, O.; Mettenleiter, T.C.; Stech, J. H9 avian influenza reassortant with engineered polybasic cleavage site displays a highly pathogenic phenotype in chicken. J. Gen. Virol. 2011, 92, 1843–1853. [Google Scholar] [CrossRef]
- Munster, V.J.; Schrauwen, E.J.A.; de Wit, E.; van den Brand, J.M.A.; Bestebroer, T.M.; Herfst, S.; Rimmelzwaan, G.F.; Osterhaus, A.D.M.E.; Fouchier, R.A.M. Insertion of a Multibasic Cleavage Motif into the Hemagglutinin of a Low-Pathogenic Avian Influenza H6N1 Virus Induces a Highly Pathogenic Phenotype. J. Virol. 2010, 84, 7953–7960. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ohuchi, R.; Ohuchi, M.; Garten, W.; Klenk, H.D. Human influenza virus hemagglutinin with high sensitivity to proteolytic activation. J. Virol. 1991, 65, 3530–3537. [Google Scholar] [CrossRef] [Green Version]
- Kawaoka, Y. Structural features influencing hemagglutinin cleavability in a human influenza A virus. J. Virol. 1991, 65, 1195–1201. [Google Scholar] [CrossRef] [Green Version]
- Stech, O.; Veits, J.; Weber, S.; Deckers, D.; Schröer, D.; Vahlenkamp, T.W.; Breithaupt, A.; Teifke, J.; Mettenleiter, T.C.; Stech, J. Acquisition of a Polybasic Hemagglutinin Cleavage Site by a Low-Pathogenic Avian Influenza Virus Is Not Sufficient for Immediate Transformation into a Highly Pathogenic Strain. J. Virol. 2009, 83, 5864–5868. [Google Scholar] [CrossRef] [Green Version]
- Schrauwen, E.J.A.; Bestebroer, T.M.; Munster, V.J.; de Wit, E.; Herfst, S.; Rimmelzwaan, G.F.; Osterhaus, A.D.M.E.; Fouchier, R.A.M. Insertion of a multibasic cleavage site in the haemagglutinin of human influenza H3N2 virus does not increase pathogenicity in ferrets. J. Gen. Virol. 2011, 92, 1410–1415. [Google Scholar] [CrossRef]
- Suarez, D.L. Influenza A virus. In Animal Influenza; Swayne, D.E., Ed.; John Wiley and Sons, Inc.: Hoboken, NJ, USA, 2017; p. 17. [Google Scholar]
- Perdue, M.; Crawford, J.; Garcia, M.; Latimer, J.; Swayne, D. Occurrence and Possible Mechanisms of Cleavage-Site Insertions in the Avian Influenza Hemagglutinin Gene. Avian Dis. 2003, 47, 182–193. [Google Scholar]
- Gultyaev, A.P.; Richard, M.; Spronken, M.I.; Olsthoorn, R.C.L.; Fouchier, R.A.M. Conserved structural RNA domains in regions coding for cleavage site motifs in hemagglutinin genes of influenza viruses. Virus Evol. 2019, 5, vez034. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gultyaev, A.P.; Spronken, M.I.; Funk, M.; Fouchier, R.A.M.; Richard, M. Insertions of codons encoding basic amino acids in H7 hemagglutinins of influenza A viruses occur by recombination with RNA at hotspots near snoRNA binding sites. RNA 2021, 27, 123–132. [Google Scholar] [CrossRef] [PubMed]
- Ito, T.; Goto, H.; Yamamoto, E.; Tanaka, H.; Takeuchi, M.; Kuwayama, M.; Kawaoka, Y.; Otsuki, K. Generation of a Highly Pathogenic Avian Influenza A Virus from an Avirulent Field Isolate by Passaging in Chickens. J. Virol. 2001, 75, 4439. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abolnik, C. Evolution of H5 highly pathogenic avian influenza: Sequence data indicate stepwise changes in the cleavage site. Arch. Virol. 2017, 162, 2219–2230. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sanjuán, R.; Domingo-Calap, P. Mechanisms of viral mutation. Cell. Mol. Life Sci. 2016, 73, 4433–4448. [Google Scholar] [CrossRef] [Green Version]
- Metreveli, G.; Zohari, S.; Ejdersund, A.; Berg, M. Phylogenetic Analysis of the Hemagglutinin Gene of Low Pathogenic Avian Influenza Virus H7N7 Strains in Mallards in Northern Europe. Avian Dis. 2010, 54, 453–456. [Google Scholar] [CrossRef]
- Perdue, M.L.; García, M.; Senne, D.; Fraire, M. Virulence-associated sequence duplication at the hemagglutinin cleavage site of avian influenza viruses. Virus Res. 1997, 49, 173–186. [Google Scholar] [CrossRef]
- Wandzik, J.M.; Kouba, T.; Karuppasamy, M.; Pflug, A.; Drncova, P.; Provaznik, J.; Azevedo, N.; Cusack, S. A Structure-Based Model for the Complete Transcription Cycle of Influenza Polymerase. Cell 2020, 181, 877–893.e21. [Google Scholar] [CrossRef]
- Poon, L.L.M.; Pritlove, D.C.; Fodor, E.; Brownlee, G.G. Direct evidence that the poly(A) tail of influenza A virus mRNA is synthesized by reiterative copying of a U track in the virion RNA template. J. Virol. 1999, 73, 3473–3476. [Google Scholar] [CrossRef] [Green Version]
- Nao, N.; Yamagishi, J.; Miyamoto, H.; Igarashi, M.; Manzoor, R.; Ohnuma, A.; Tsuda, Y.; Furuyama, W.; Shigeno, A.; Kajihara, M.; et al. Genetic predisposition to acquire a polybasic cleavage site for highly pathogenic avian influenza virus hemagglutinin. MBio 2017, 8, e02298-16. [Google Scholar] [CrossRef] [Green Version]
- Brugh, M.; Perdue, M.L. Emergence of Highly Pathogenic Virus during Selective Chicken Passage of the Prototype Mildly Pathogenic Chicken/Pennsylvania/83 (H5N2) Influenza Virus. Avian Dis. 1991, 35, 824–833. [Google Scholar] [CrossRef] [PubMed]
- Horimoto, T.; Kawaoka, Y. Molecular Changes in virulent mutants arising from avirulent avian influenza viruses during Replication in 14-day-old embryonated eggs. Virology 1995, 206, 755–759. [Google Scholar] [CrossRef] [Green Version]
- Perdue, M.L.; Garcia, M.; Beck, J.; Brugh, M.; Swayne, D.E. An arg-lys insertion at the hemagglutinin cleavage site of an H5N2 avian influenza isolate. Virus Genes 1996, 12, 77–84. [Google Scholar] [CrossRef]
- Silvano, F.D.; Yoshikawa, M.; Shimada, A.; Otsuki, K.; Umemura, T. Enhanced Neuropathogenicity of Avian Influenza A Virus by Passages through Air Sac and Brain of Chicks. J. Vet. Med. Sci 1997, 59, 143–148. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shinya, K.; Awakura, T.; Shimada, A.; Silvano, F.D.; Umemura, T.; Otsuki, K. Pathogenesis of pancreatic atrophy by avian influenza a virus infection. Avian Pathol. 1995, 24, 623–632. [Google Scholar] [CrossRef]
- Brugh, M.; Beck, J.R. Recovery of Minority Subpopulations of Highly Pathogenic Avian Influenza Virus. Avian Dis. 2003, 47, 166–174. [Google Scholar]
- Laleye, A.T.; Abolnik, C. Emergence of highly pathogenic H5N2 and H7N1 influenza A viruses from low pathogenic precursors by serial passage in ovo. PLoS ONE 2020, 15, e0240290. [Google Scholar] [CrossRef]
- Seekings, A.H.; Howard, W.A.; Nuñéz, A.; Slomka, M.J.; Banyard, A.C.; Hicks, D.; Ellis, R.J.; Nuñéz-García, J.; Hartgroves, L.C.; Barclay, W.S.; et al. The emergence of H7N7 highly pathogenic avian influenza virus from low pathogenicity avian influenza virus using an in ovo embryo culture model. Viruses 2020, 12, 920. [Google Scholar] [CrossRef]
- Kida, Y.; Okuya, K.; Saito, T.; Yamagishi, J.; Ohnuma, A.; Hattori, T.; Miyamoto, H.; Manzoor, R.; Yoshida, R.; Nao, N.; et al. Structural Requirements in the Hemagglutinin Cleavage Site-Coding RNA Region for the Generation of Highly Pathogenic Avian Influenza Virus. Pathogens 2021, 10, 1597. [Google Scholar] [CrossRef]
- Brugh, M. Highly pathogenic virus recovered from chickens infected with mildly pathogenic 1986 isolates of H5N2 avian influenza virus. Avian Dis. 1988, 32, 695–703. [Google Scholar] [CrossRef]
- Khatchikian, D.; Orlich, M.; Rott, R. Increased viral pathogenicity after insertion of a 28S ribosomal RNA sequence into the haemagglutinin gene of an influenza virus. Nature 1989, 340, 156–157. [Google Scholar] [CrossRef] [PubMed]
- Orlich, M.; Khatchikian, D.; Teigler, A.; Rott, R. Structural variation occurring in the hemagglutinin of influenza virus A/turkey/Oregon/71 during adaptation to different cell types. Virology 1990, 176, 531–538. [Google Scholar] [CrossRef]
- Li, S.Q.; Orlich, M.; Rott, R. Generation of seal influenza virus variants pathogenic for chickens, because of hemagglutinin cleavage site changes. J. Virol. 1990, 64, 3297–3303. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Williams, G.D.; Townsend, D.; Wylie, K.M.; Kim, P.J.; Amarasinghe, G.K.; Kutluay, S.B.; Boon, A.C.M. Nucleotide resolution mapping of influenza A virus nucleoprotein-RNA interactions reveals RNA features required for replication. Nat. Commun. 2018, 9, 465. [Google Scholar] [CrossRef] [Green Version]
- Le Sage, V.; Nanni, A.V.; Bhagwat, A.R.; Snyder, D.J.; Cooper, V.S.; Lakdawala, S.S.; Lee, N. Non-uniform and non-random binding of nucleoprotein to influenza A and B viral RNA. Viruses 2018, 10, 522. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gultyaev, A.P.; Spronken, M.I.; Richard, M.; Schrauwen, E.J.A.; Olsthoorn, R.C.L.; Fouchier, R.A.M. Subtype-specific structural constraints in the evolution of influenza A virus hemagglutinin genes. Sci. Rep. 2016, 6, 38892. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dupré, G.; Hoede, C.; Figueroa, T.; Bessière, P.; Bertagnoli, S.; Ducatez, M.; Gaspin, C.; Volmer, R. Phylodynamic study of the conserved RNA structure encompassing the hemagglutinin cleavage site encoding region of H5 and H7 low pathogenic avian influenza viruses. Virus Evol. 2021, 7, veab093. [Google Scholar] [CrossRef]
- Kouba, T.; Drncová, P.; Cusack, S. Structural snapshots of actively transcribing influenza polymerase. Nat. Struct. Mol. Biol. 2019, 26, 460–470. [Google Scholar] [CrossRef] [PubMed]
- Banks, J.; Speidel, E.S.; Moore, E.; Plowright, L.; Piccirillo, A.; Capua, I.; Cordioli, P.; Fioretti, A.; Alexander, D.J. Changes in the haemagglutinin and the neuraminidase genes prior to the emergence of highly pathogenic H7N1 avian influenza viruses in Italy. Arch Virol 2001, 146, 963–973. [Google Scholar] [CrossRef] [PubMed]
- Bratkovič, T.; Bozič, J.; Rogelj, B. Functional diversity of small nucleolar RNAs. Nucleic Acids Res. 2020, 48, 1627–1651. [Google Scholar] [CrossRef] [Green Version]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
de Bruin, A.C.M.; Funk, M.; Spronken, M.I.; Gultyaev, A.P.; Fouchier, R.A.M.; Richard, M. Hemagglutinin Subtype Specificity and Mechanisms of Highly Pathogenic Avian Influenza Virus Genesis. Viruses 2022, 14, 1566. https://doi.org/10.3390/v14071566
de Bruin ACM, Funk M, Spronken MI, Gultyaev AP, Fouchier RAM, Richard M. Hemagglutinin Subtype Specificity and Mechanisms of Highly Pathogenic Avian Influenza Virus Genesis. Viruses. 2022; 14(7):1566. https://doi.org/10.3390/v14071566
Chicago/Turabian Stylede Bruin, Anja C. M., Mathis Funk, Monique I. Spronken, Alexander P. Gultyaev, Ron A. M. Fouchier, and Mathilde Richard. 2022. "Hemagglutinin Subtype Specificity and Mechanisms of Highly Pathogenic Avian Influenza Virus Genesis" Viruses 14, no. 7: 1566. https://doi.org/10.3390/v14071566