
Occurrence, Distribution, and Transmission of Alfalfa Viruses in China

 , , and
, , and 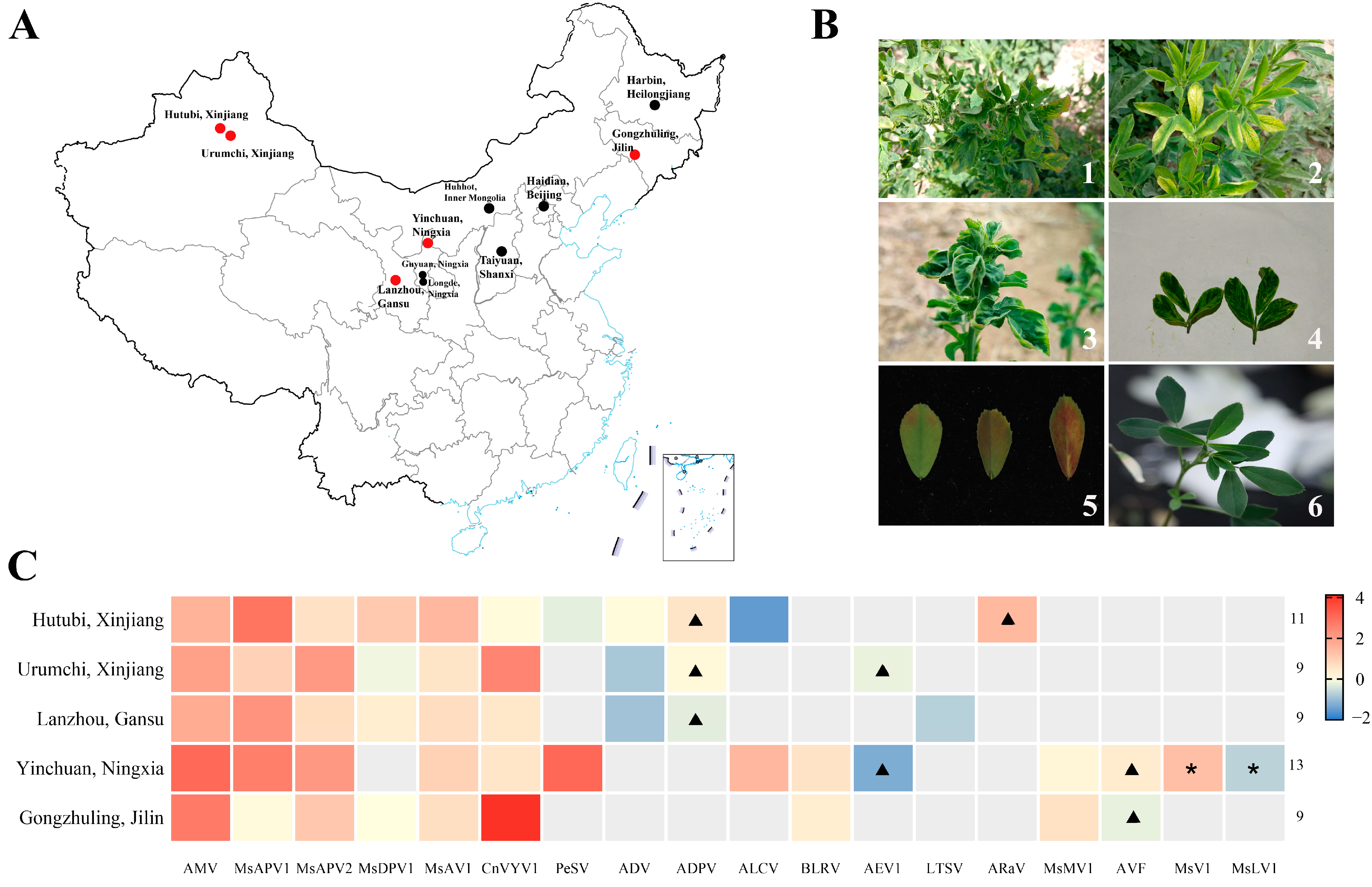{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant and Insect Cultures
2.2. Total RNA Extraction and Reverse Transcription
2.3. Sequencing and Analysis Data
2.4. Phylogenetic Analysis
2.5. Rearing of O. loti Nymph, Acquisition, and Transmission of AMV
2.6. Virus Detection
2.7. Statistical Analysis
3. Results
3.1. Identification and Verification of Alfalfa Virus
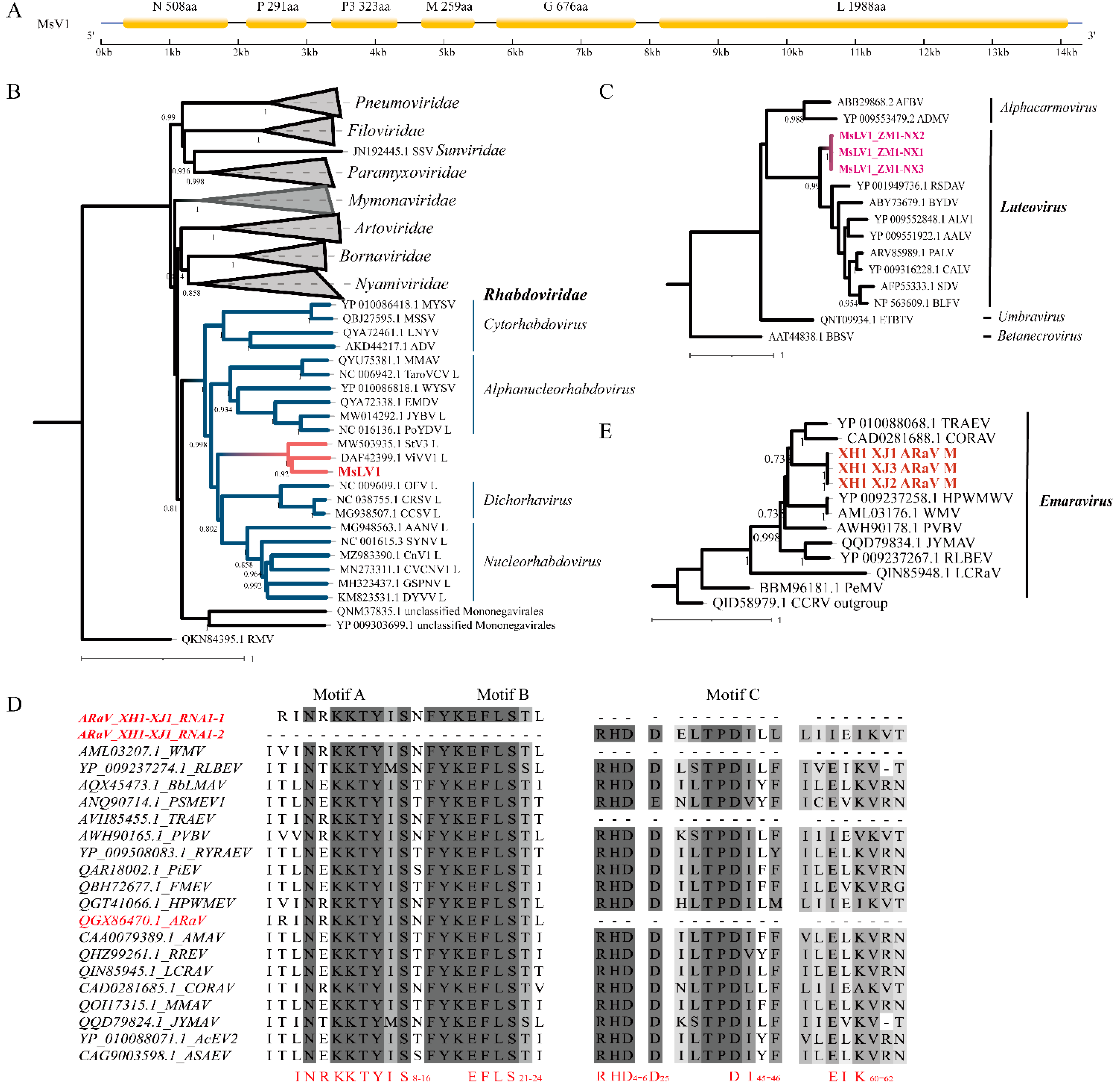
3.2. Genome Analysis Viruses
3.2.1. Medicago sativa Virus 1
3.2.2. Medicago sativa Luteovirus 1
3.2.3. Other Novel Viruses
3.3. Incidence and Distribution of the Main Viruses
3.4. Transmission of AMV by O. loti
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- McNeill, M.R.; Tu, X.; Altermann, E.; Beilei, W.; Shi, S. Sustainable management of Medicago sativa for future climates: Insect pests, endophytes and multitrophic interactions in a complex environment. Front. Agron. 2022, 4, 825087. [Google Scholar] [CrossRef]
- Ruckle, M.E.; Meier, M.A.; Frey, L.; Eicke, S.; Kölliker, R.; Zeeman, S.C.; Studer, B. Diurnal leaf starch content: An orphan trait in forage legumes. Agronomy 2017, 7, 16. [Google Scholar] [CrossRef] [Green Version]
- Wang, R.; Xu, W. Development characteristics and trend of alfalfa industry in China. J. Agric. Sci. Technol. 2021, 23, 7–12. [Google Scholar]
- Tao, Y.; Sun, Q.; Xu, L.; Li, F. Trends and challenges of alfalfa industry in China. Grassl. Prataculture 2022, 34, 1–10. [Google Scholar]
- National Animal Husbandry Service. China Prataculture Statistics (2016); China Agricultural Press: Beijing, China, 2017; pp. 19–21. [Google Scholar]
- Han, Y.; Hu, H.; Yu, Y.; Zhang, Z.; Fan, Z. Effects of alfalfa mosaic disease on photosynthetic performance, growth, and forage quality of Medicago sativa. Pratacultural Sci. 2019, 36, 2061–2068. [Google Scholar]
- Jaime, C.; Muchut, S.E.; Reutemann, A.G.; Gieco, J.O.; Dunger, G. Morphological changes, alteration of photosynthetic parameters and chlorophyll production induced by infection with alfalfa dwarf virus in Medicago sativa plants. Plant Pathol. 2020, 69, 393–402. [Google Scholar] [CrossRef]
- Davoodi, Z.; Heydarnejad, J.; Massumi, H.; Richet, C.; Galzi, S.; Filloux, D.; Roumagnac, P. First report of alfalfa leaf curl virus from alfalfa in Iran. Plant Dis. 2018, 102, 2385. [Google Scholar] [CrossRef] [PubMed]
- Samarfard, S.; McTaggart, A.R.; Sharman, M.; Bejerman, N.E.; Dietzgen, R.G. Viromes of ten alfalfa plants in Australia reveal diverse known viruses and a novel RNA virus. Pathogens 2020, 9, 214. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nan, Z. Alfalfa disease and its integrated control system in China. Anim. Sci. Vet. Med. 2001, 18, 81–84. [Google Scholar]
- Zhao, L.; Cao, M.; Huang, Q.; Wang, Y.; Sun, J.; Zhang, Y.; Hou, C.; Wu, Y. Occurrence and distribution of actinidia viruses in Shaanxi Province of China. Plant Dis. 2021, 105, 929–939. [Google Scholar] [CrossRef]
- Villamor, D.E.V.; Ho, T.; Al Rwahnih, M.; Martin, R.R.; Tzanetakis, I.E. High throughput sequencing for plant virus detection and discovery. Phytopathology 2019, 109, 716–725. [Google Scholar] [CrossRef]
- Wu, Z.; Yang, L.; Ren, X.; He, G.; Zhang, J.; Yang, J.; Qian, Z.; Dong, J.; Sun, L.; Zhu, Y.; et al. Deciphering the bat virome catalog to better understand the ecological diversity of bat viruses and the bat origin of emerging infectious diseases. ISME J. 2016, 10, 609–620. [Google Scholar] [CrossRef] [Green Version]
- Ng, T.F.F.; Duffy, S.; Polston, J.E.; Bixby, E.; Vallad, G.E.; Breitbart, M. Exploring the diversity of plant DNA viruses and their satellites using vector-enabled metagenomics on whiteflies. PLoS ONE 2011, 6, e19050. [Google Scholar] [CrossRef] [Green Version]
- Hodge, B.A.; Paul, P.A.; Stewart, L.R. Occurrence and high-throughput sequencing of viruses in ohio wheat. Plant Dis. 2020, 104, PDIS08191724RE. [Google Scholar] [CrossRef]
- Guo, Z.; Zhang, T.; Chen, Z.; Niu, J.; Cui, X.; Mao, Y.; Hassan, M.U.; Kareem, H.A.; Xu, N.; Sui, X.; et al. Occurrence, distribution, and genetic diversity of alfalfa (Medicago sativa L.) viruses in four major alfalfa-producing provinces of China. Front. Microbiol. 2022, 12, 771361. [Google Scholar] [CrossRef]
- Li, J.; Gu, H.; Liu, Y.; Wei, S.; Hu, G.; Wang, X.; McNeill, M.R.; Ban, L. RNA-seq reveals plant virus composition and diversity in alfalfa, thrips, and aphids in Beijing, China. Arch. Virol. 2021, 166, 1711–1722. [Google Scholar] [CrossRef]
- Dietzgen, R.G.; Mann, K.S.; Johnson, K.N. Plant virus–insect vector interactions: Current and potential future research directions. Viruses 2016, 8, 303. [Google Scholar] [CrossRef]
- Dixon, A.F.G. Aphid Ecology an Optimization Approach, 2nd ed.; Springer Science + Business Media: New York, NY, USA, 1998. [Google Scholar]
- Garran, J.; Gibbs, A. Studies on alfalfa mosaic virus and alfalfa aphids. Aust. J. Agric. Res. 1982, 33, 657–664. [Google Scholar] [CrossRef]
- Hill, J.H.; Alleman, R.; Hogg, D.B.; Grau, C.R. First report of transmission of Soybean mosaic virus and Alfalfa mosaic virus by Aphis glycines in the New World. Plant Dis. 2001, 85, 561. [Google Scholar] [CrossRef] [Green Version]
- Ryckebusch, F.; Sauvion, N.; Granier, M.; Roumagnac, P.; Peterschmitt, M. Alfalfa leaf curl virus is transmitted by Aphis craccivora in a highly specific circulative manner. Virology 2020, 546, 98–108. [Google Scholar] [CrossRef]
- Davis, T.S.; Wu, Y.; Eigenbrode, S.D. The effects of Bean leafroll virus on life history traits and host selection behavior of specialized pea aphid (Acyrthosiphon pisum, Hemiptera: Aphididae) genotypes. Environ. Entomol. 2017, 46, 68–74. [Google Scholar] [PubMed]
- McNeill, M.R.; Tu, X.; Ferguson, C.M.; Ban, L.; Hardwick, S.; Rong, Z.; Barratt, B.I.P.; Zehua, Z. Diversity and impacts of key grassland and forage arthropod pests in China and New Zealand: An overview of IPM and biosecurity opportunities. NeoBiota 2021, 65, 137–168. [Google Scholar] [CrossRef]
- Zhang, X.; Liu, C.; Yan, L.; Chen, Y.; Wei, L.; Qian, X. Study on the population dynamics of main arthropod groups in alfalfa fields. Acta Agrestia Sin. 2007, 15, 556–560. [Google Scholar]
- Wu, Y.; Li, X.; Zhao, X.; Wei, L. The endangerment of thrips to alfalfa. Chin. Grassl. 1988, 2, 25–27. [Google Scholar]
- Ma, J.; Chen, H.; Wang, Y.; Wang, J.; Du, Y. Species investigation and identification of alfalfa thrips in Ningxia. Jiangsu Agric. Sci. 2017, 45, 88–91. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.; Zhao, X.; Buhe, T. The thrips are main injurious insects of alfalfa production in China. Grassl. China 1990, 3, 65–66+64. [Google Scholar]
- Zhang, R.; Yang, F.; Xian, C.; Ma, J.; Zhang, S. A study on the yield loss and economic threshold of alfalfa damaged by thrip, Odontothrips loti. Plant Prot. 2005, 31, 47–49. [Google Scholar]
- Liu, Y.Q.; Li, J.; Ban, L.P. Morphology and distribution of antennal sensilla in three species of Thripidae (Thysanoptera) infesting alfalfa Medicago sativa. Insects 2021, 12, 81. [Google Scholar] [CrossRef]
- Zhang, X.; Pen, R.; Hu, G. Effect of Odontothrips loti nymphs sustainable damage on alfalfa growth. Grassl. Turf 2017, 37, 8–13. [Google Scholar]
- Jones, D.R. Plant viruses transmitted by thrips. Eur. J. Plant Pathol. 2005, 113, 119–157. [Google Scholar] [CrossRef]
- Rotenberg, D.; Jacobson, A.L.; Schneweis, D.J.; Whitfield, A.E. Thrips transmission of tospoviruses. Curr. Opin. Virol. 2015, 15, 80–89. [Google Scholar] [CrossRef]
- Mason, G.; Roggero, P.; Tavella, L. Detection of Tomato spotted wilt virus in its vector Frankliniella occidentalis by reverse transcription-polymerase chain reaction. J. Virol. Methods 2003, 109, 69–73. [Google Scholar] [CrossRef]
- Chen, H.; Zeng, Y.; Yang, Y.; Huang, L.; Tang, B.; Zhang, H.; Hao, F.; Liu, W.; Li, Y.; Liu, Y.; et al. Allele-aware chromosome-level genome assembly and efficient transgene-free genome editing for the autotetraploid cultivated alfalfa. Nat. Commun. 2020, 11, 2494. [Google Scholar] [CrossRef]
- Shen, C.; Du, H.; Chen, Z.; Lu, H.; Zhu, F.; Chen, H.; Meng, X.; Liu, Q.; Liu, P.; Zheng, L.; et al. The chromosome-level genome sequence of the autotetraploid alfalfa and resequencing of core germplasms provide genomic resources for alfalfa research. Mol. Plant 2020, 13, 1250–1261. [Google Scholar] [CrossRef]
- Li, H. Aligning sequence reads, clone sequences and assembly contigs with BWA-MEM. arXiv 2013, arXiv:1303.3997. [Google Scholar]
- Li, H.; Handsaker, B.; Wysoker, A.; Fennell, T.; Ruan, J.; Homer, N.; Marth, G.; Abecasis, G.; Durbin, R.; Subgroup, G.P.D.P. The sequence alignment/map format and samtools. Bioinformatics 2009, 25, 2078–2079. [Google Scholar] [CrossRef] [Green Version]
- Barnett, D.W.; Garrison, E.K.; Quinlan, A.R.; Strömberg, M.P.; Marth, G.T. BamTools: A C++ API and toolkit for analyzing and managing BAM files. Bioinformatics 2011, 27, 1691–1692. [Google Scholar] [CrossRef]
- Bankevich, A.; Nurk, S.; Antipov, D.; Gurevich, A.A.; Dvorkin, M.; Kulikov, A.S.; Lesin, V.M.; Nikolenko, S.I.; Pham, S.; Prjibelski, A.D.; et al. SPAdes: A new genome assembly algorithm and its applications to single-cell sequencing. J. Comput. Biol. 2012, 19, 455–477. [Google Scholar] [CrossRef] [Green Version]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular evolutionary genetics analysis across computing platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef]
- Letunic, I.; Bork, P. Interactive Tree Of Life (iTOL) v4: Recent updates and new developments. Nucleic Acids Res. 2019, 47, W256–W259. [Google Scholar] [CrossRef] [Green Version]
- Wan, Y.; Hussain, S.; Merchant, A.; Xu, B.; Xie, W.; Wang, S.; Zhang, Y.; Zhou, X.; Wu, Q. Tomato spotted wilt orthotospovirus influences the reproduction of its insect vector, western flower thrips, Frankliniella occidentalis, to facilitate transmission. Pest Manag. Sci. 2020, 76, 2406–2414. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T. Analysis of relative gene expression data using real-time quantitative PCR and the 2−DDCt method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Walker, P.J.; Blasdell, K.R.; Calisher, C.H.; Dietzgen, R.G.; Kondo, H.; Kurath, G.; Longdon, B.; Stone, D.M.; Tesh, R.B.; Tordo, N.; et al. ICTV Virus Taxonomy Profile: Rhabdoviridae. J. Gen. Virol. 2018, 99, 447–448. [Google Scholar] [CrossRef]
- Domier, L. Family-Luteoviridae. In Virus Taxonomy; King, A.M.Q., Adams, M.J., Carstens, E.B., Lefkowitz, E.J., Eds.; Elsevier: San Diego, CA, USA, 2012; pp. 1045–1053. [Google Scholar]
- Elbeaino, T.; Digiaro, M.; Mielke-Ehret, N.; Muehlbach, H.-P.; Martelli, G.P.; Consortium, I.R. ICTV Virus Taxonomy Profile: Fimoviridae. J. Gen. Virol. 2018, 99, 1478–1479. [Google Scholar] [CrossRef] [PubMed]
- Bejerman, N.; Nome, C.; Giolitti, F.; Kitajima, E.; de Breuil, S.; Pérez Fernández, J.; Basigalup, D.; Cornacchione, M.; Lenardon, S. First report of a Rhabdovirus infecting alfalfa in Argentina. Plant Dis. 2011, 95, 771. [Google Scholar] [CrossRef] [PubMed]
- Gaafar, Y.Z.A.; Richert-Poggeler, K.R.; Maass, C.; Vetten, H.J.; Ziebell, H. Characterisation of a novel nucleorhabdovirus infecting alfalfa (Medicago sativa). J. Virol. 2019, 16, 55. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gao, Q.; Xuehelati, R.; Wang, Y.; Li, K.; Wang, X.-B. Identification of Alfalfa dwarf virus in Xinjiang Province, China. Plant Dis. 2021, 105, 3768. [Google Scholar] [CrossRef]
- Refukati, X.; Fan, J.; Wang, L.; Ge, R.; Li, K. Identification and detection of Alfalfa dwarf virus (ADV) isolates in China. J. Xinjiang Agric. Univ. 2020, 43, 177–181. [Google Scholar]
- Li, K.; Muhanmaiti, A.; Ge, R.; LIiu, X.; Li, B.; Xuehelati, R. Identification of a new virus on alfalfa in Xinjiang. Pratacultural Sci. 2019, 36, 2319–2324. [Google Scholar]
- Trucco, V.; de Breuil, S.; Bejerman, N.; Lenardon, S.; Giolitti, F. Bean leafroll virus (BLRV) in Argentina: Molecular characterization and detection in alfalfa fields. Eur. J. Plant Pathol. 2016, 146, 207–212. [Google Scholar] [CrossRef]
- Kim, H.; Park, D.; Hahn, Y. Identification of novel RNA viruses in alfalfa (Medicago sativa): An Alphapartitivirus, a Deltapartitivirus, and a Marafivirus. Gene 2018, 638, 7–12. [Google Scholar] [CrossRef]
- Nemchinov, L.G.; François, S.; Roumagnac, P.; Ogliastro, M.; Hammond, R.W.; Mollov, D.S.; Filloux, D. Characterization of alfalfa virus F, a new member of the genus Marafivirus. PLoS ONE 2018, 13, e0203477. [Google Scholar] [CrossRef]
- Bejerman, N.; Giolitti, F.; Trucco, V.; de Breuil, S.; Dietzgen, R.G.; Lenardon, S. Complete genome sequence of a new enamovirus from Argentina infecting alfalfa plants showing dwarfism symptoms. Arch. Virol. 2016, 161, 2029–2032. [Google Scholar] [CrossRef]
- Nemchinov, L.G.; Grinstead, S.C.; Mollov, D.S. First report and complete genome sequence of Alfalfa enamovirus from Sudan. Genome Announc. 2017, 5, e00531-17. [Google Scholar] [CrossRef] [Green Version]
- Trucco, V.; Cabrera Mederos, D.; Lenardon, S.; Giolitti, F. Geographical distribution, genetic studies and vector transmission of alfalfa enamovirus-1 infecting alfalfa crop in Argentina. Virus Genes 2020, 56, 662–667. [Google Scholar] [CrossRef]
- Tao, S.; Wang, Y.-T.; Zhang, Q. Situation of domestic alfalfa market and the impact from Sino-US trade frictions and its countermeasures. Anim. Husb. Feed. Sci. 2019, 40, 46–50. [Google Scholar]
- Nibert, M.L.; Ghabrial, S.A.; Maiss, E.; Lesker, T.; Vainio, E.J.; Jiang, D.; Suzuki, N. Taxonomic reorganization of family Partitiviridae and other recent progress in partitivirus research. Virus Res. 2014, 188, 128–141. [Google Scholar] [CrossRef]
- Zhou, Q.; Liang, Q.; Han, L. Symptoms and pathogen detection of alfalfa virus disease. Pratacultural Sci. 2016, 33, 1297–1305. [Google Scholar]
- Bejerman, N.; Debat, H.; Nome, C.; Cabrera-Mederos, D.; Trucco, V.; de Breuil, S.; Lenardon, S.; Giolitti, F. Redefining the Medicago sativa alphapartitiviruses genome sequences. Virus Res. 2019, 265, 156–161. [Google Scholar] [CrossRef]
- Yoo, R.H.; Zhao, F.; Lim, S.; Igori, D.; Kim, S.-M.; An, T.-J.; Lee, S.-H.; Moon, J.S. The complete genome sequences of two isolates of cnidium vein yellowing virus, a tentative new member of the family Secoviridae. Arch. Virol. 2015, 160, 2911–2914. [Google Scholar] [CrossRef]
- Jiang, P.; Shao, J.; Nemchinov, L.G. Identification of emerging viral genomes in transcriptomic datasets of alfalfa (Medicago sativa L.). Virol. J. 2019, 16, 153. [Google Scholar] [CrossRef]
- vande Wetering, F.; Goldbach, R.; Peters, D. Tomato spotted wilt tospovirus ingestion by first instar larvae of Frankliniella occidentalis is a prerequisite for transmission. Phytopathology 1996, 86, 900–905. [Google Scholar] [CrossRef]
- De Assis Filho, F.M.; Naidu, R.A.; Deom, C.M.; Sherwood, J.L. Dynamics of Tomato spotted wilt virus replication in the alimentary canal of two thrips species. Phytopathology 2002, 92, 729–733. [Google Scholar] [CrossRef] [Green Version]
- De Assis Filho, F.M.; Deom, C.M.; Sherwood, J.L. Acquisition of Tomato spotted wilt virus by adults of two thrips species. Phytopathology 2004, 94, 333–336. [Google Scholar] [CrossRef] [Green Version]
- Wijkamp, I.; Peters, D. Determination of the median latent period of two Tospoviruses in Frankliniella occidentalis, using a novel leaf disk assay. Phytopathology 1993, 83, 986–991. [Google Scholar] [CrossRef]
- Wan, Y.; Zheng, X.; Xu, B.; Xie, W.; Wang, S.; Zhang, Y.; Zhou, X.; Wu, Q. Insecticide resistance increases the vector competence: A case study in Frankliniella occidentalis. J. Pest Sci. 2021, 94, 83–91. [Google Scholar] [CrossRef]
- Sakurai, T.; Murai, T.; Maeda, T.; Tsumuki, H. Sexual differences in transmission and accumulation of tomato spotted wilt virus in its insect vector Frankliniella occidentalis (Thysanoptera: Thripidae). Appl. Entomol. Zool. 1998, 33, 583–588. [Google Scholar] [CrossRef] [Green Version]
- Van de Wetering, F.; Hulshof, J.; Posthuma, K.; Harrewijn, P.; Goldbach, R.; Peters, D. Distinct feeding behavior between sexes of Frankliniella occidentalis results in higher scar production and lower tospovirus transmission by females. Entomol. Exp. Appl. 1998, 88, 9–15. [Google Scholar] [CrossRef]
- Ogada, P.A.; Poehling, H.-M. Sex-Specific Influences of Frankliniella occidentalis (Western Flower Thrips) in the Transmission of Tomato Spotted Wilt Virus (Tospovirus). J. Plant Dis. Prot. 2015, 122, 264–274. [Google Scholar] [CrossRef]
- Rotenberg, D.; Krishna Kumar, N.K.; Ullman, D.E.; Montero-Astúa, M.; Willis, D.K.; German, T.L.; Whitfield, A.E. Variation in Tomato spotted wilt virus titer in Frankliniella occidentalis and its association with frequency of transmission. Phytopathology 2009, 99, 404–410. [Google Scholar] [CrossRef] [Green Version]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, J.; Shang, Q.; Liu, Y.; Dai, W.; Li, X.; Wei, S.; Hu, G.; McNeill, M.R.; Ban, L. Occurrence, Distribution, and Transmission of Alfalfa Viruses in China. Viruses 2022, 14, 1519. https://doi.org/10.3390/v14071519
Li J, Shang Q, Liu Y, Dai W, Li X, Wei S, Hu G, McNeill MR, Ban L. Occurrence, Distribution, and Transmission of Alfalfa Viruses in China. Viruses. 2022; 14(7):1519. https://doi.org/10.3390/v14071519
Chicago/Turabian StyleLi, Jin, Qiaoxia Shang, Yanqi Liu, Wenting Dai, Xin Li, Shuhua Wei, Guixin Hu, Mark Richard McNeill, and Liping Ban. 2022. "Occurrence, Distribution, and Transmission of Alfalfa Viruses in China" Viruses 14, no. 7: 1519. https://doi.org/10.3390/v14071519