
Dynamic Interactions of Post Cleaved NS2B Cofactor and NS3 Protease Identified by Integrative Structural Approaches

 , , ,
, , ,
Abstract
:1. Introduction
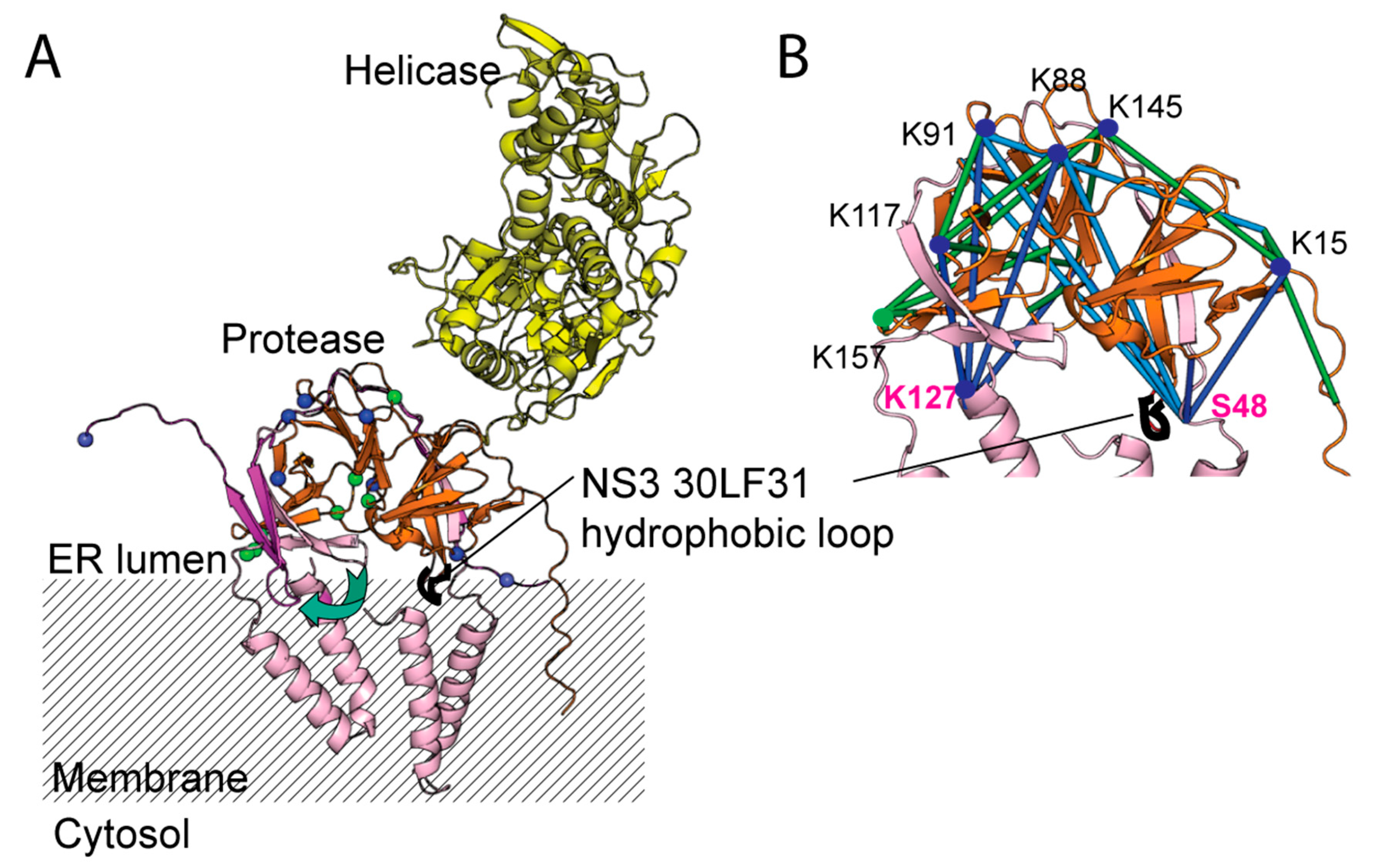
2. Results
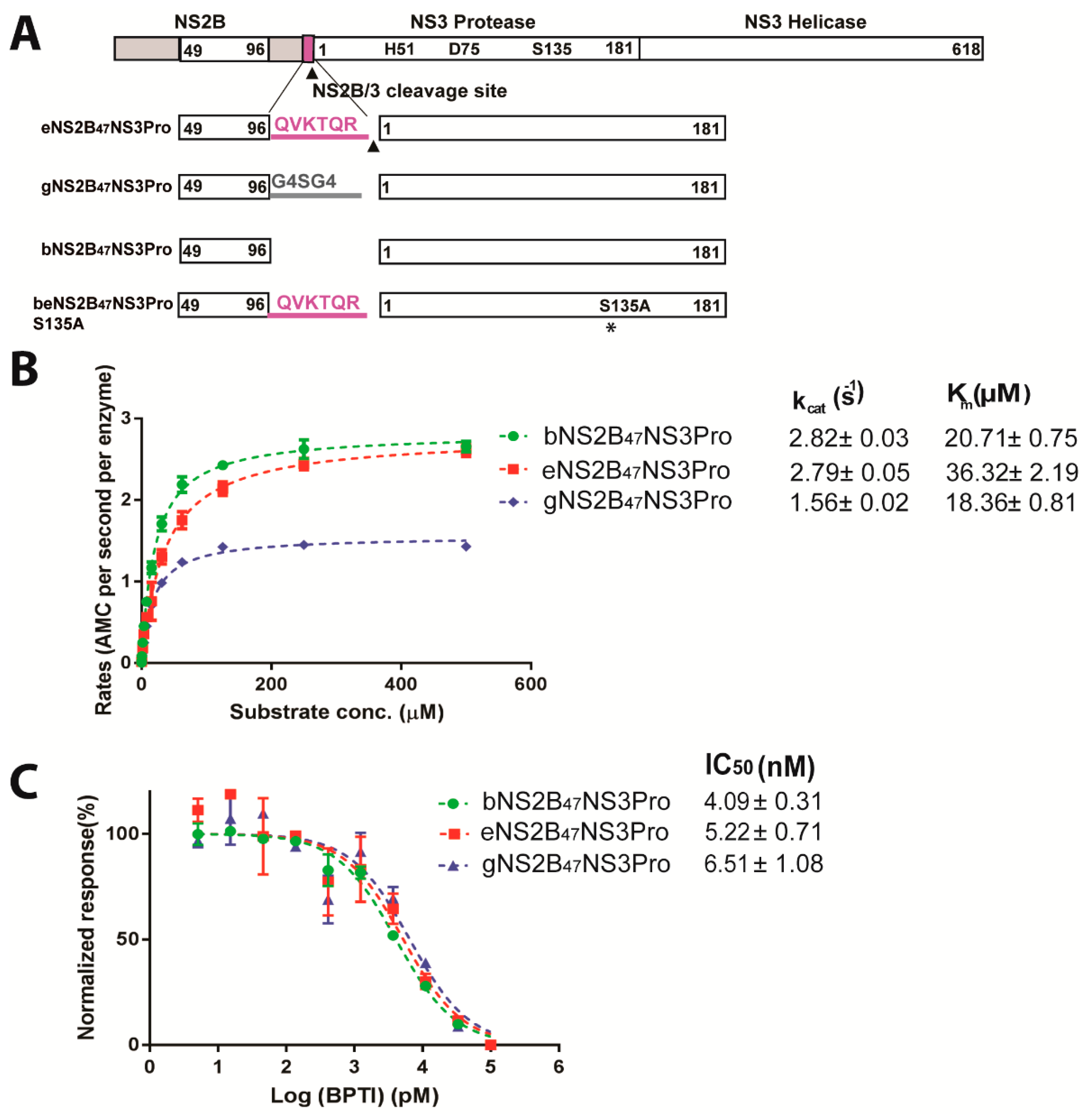
2.1. DENV4 NS2B/3 Cleavage Site at NS2B C-Terminus Does Not Interfere with Protease Activity
2.2. Molecular Weight Determination of DENV4 b-, e-, g-NS2B47NS3Pro Constructs by LC-MS
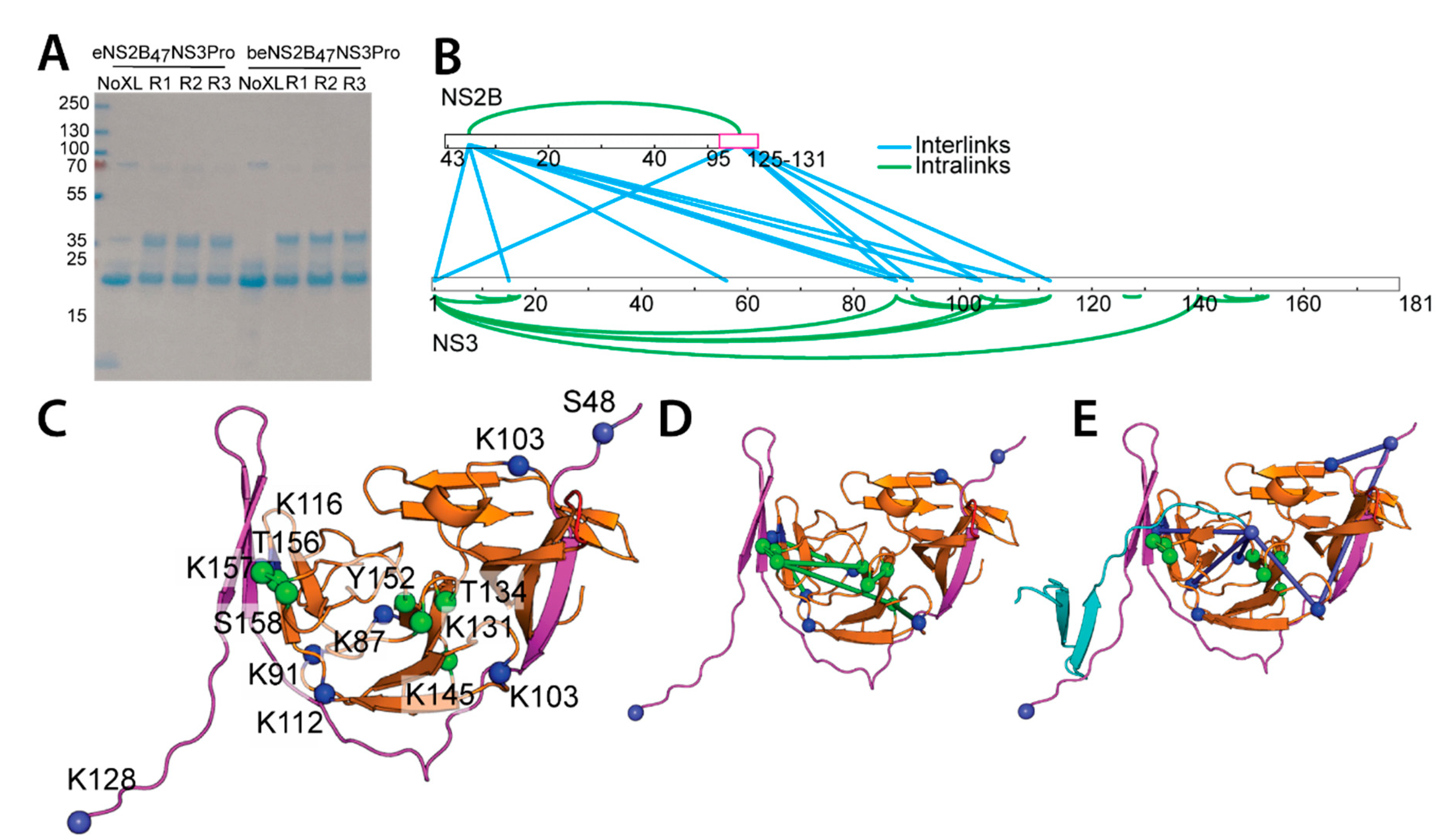
2.3. Mapping the Long-Range Interactions between DENV4 NS2B and NS3 Protease via Cross-Linking Mass Spectrometry
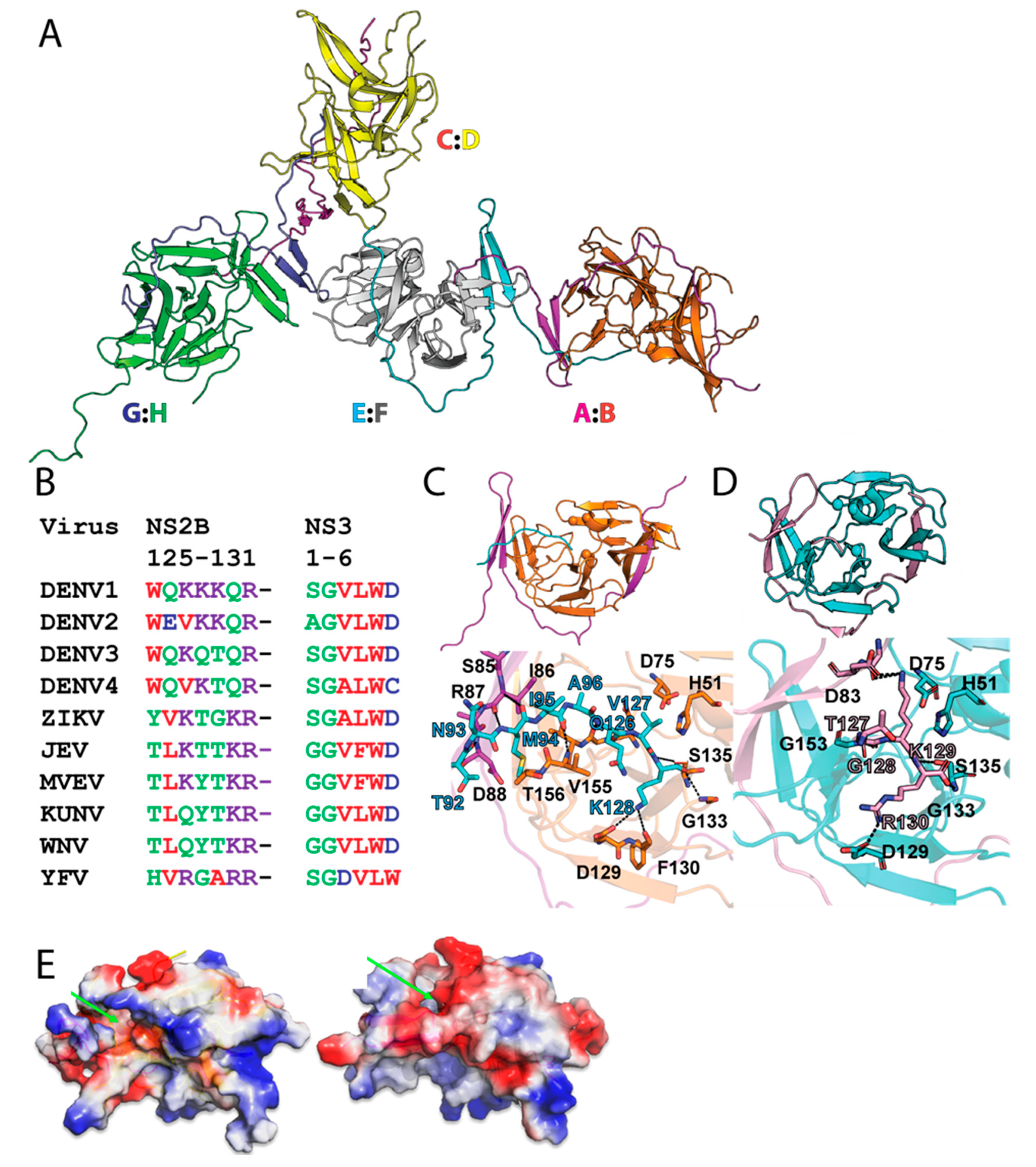
3. Discussion
4. Materials and Method
4.1. Protein Purification
4.2. Protease Activity Assay
4.3. Protease Inhibition Assay
4.4. Crystallization
4.5. eNS2B47NS3Pro S135A Mutagenesis
4.6. beNS2B47NS3Pro Mutagenesis
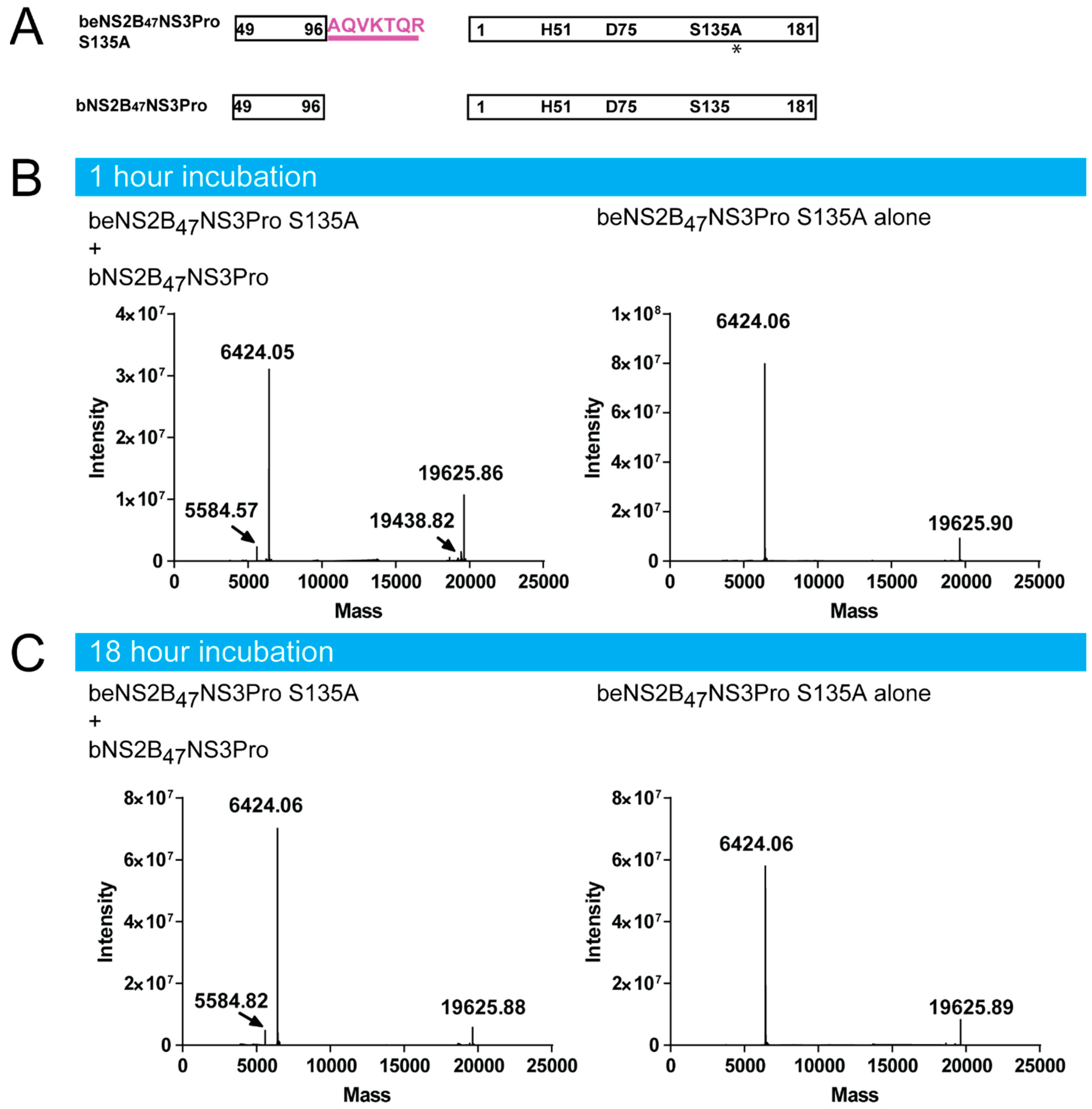
4.7. beNS2B47NS3Pro S135a Digestion by bNS2B47NS3Pro
4.8. Intact Protein LC-MS Analysis and Data Processing
4.9. Cross-Linking Mass Spectrometry and Data Analysis
4.10. 3D Structure Prediction by AlphaFold
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kyle, J.L.; Harris, E. Global Spread and Persistence of Dengue. Annu. Rev. Microbiol. 2008, 62, 71–92. [Google Scholar] [CrossRef] [Green Version]
- Chambers, T.J.; Hahn, C.S.; Galler, R.; Rice, C.M. Flavivirus Genome Organization, Expression, and Replication. Annu. Rev. Microbiol. 1990, 44, 649–688. [Google Scholar] [CrossRef]
- Falgout, B.; Pethel, M.; Zhang, Y.M.; Lai, C.J. Both nonstructural proteins NS2B and NS3 are required for the proteolytic processing of dengue virus nonstructural proteins. J. Virol. 1991, 65, 2467–2475. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cahour, A.; Falgout, B.; Lai, C.J. Cleavage of the dengue virus polyprotein at the NS3/NS4A and NS4B/NS5 junctions is mediated by viral protease NS2B-NS3, whereas NS4A/NS4B may be processed by a cellular protease. J. Virol. 1992, 66, 1535–1542. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Amberg, S.M.; Nestorowicz, A.; McCourt, D.W.; Rice, C.M. NS2B-3 proteinase-mediated processing in the yellow fever virus structural region: In vitro and in vivo studies. J. Virol. 1994, 68, 3794–3802. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, W.-N.; Loscha, K.V.; Nitsche, C.; Graham, B.; Otting, G. The dengue virus NS2B-NS3 protease retains the closed conformation in the complex with BPTI. FEBS Lett. 2014, 588, 2206–2211. [Google Scholar] [CrossRef] [Green Version]
- Assenberg, R.; Mastrangelo, E.; Walter, T.S.; Verma, A.; Milani, M.; Owens, R.J.; Stuart, D.I.; Grimes, J.M.; Mancini, E.J. Crystal Structure of a Novel Conformational State of the Flavivirus NS3 Protein: Implications for Polyprotein Processing and Viral Replication. J. Virol. 2009, 83, 12895–12906. [Google Scholar] [CrossRef] [Green Version]
- Erbel, P.; Schiering, N.; D’Arcy, A.; Renatus, M.; Kroemer, M.; Lim, S.P.; Yin, Z.; Keller, T.; Vasudevan, S.G.; Hommel, U. Structural basis for the activation of flaviviral NS3 proteases from dengue and West Nile virus. Nat. Struct. Mol. Biol. 2006, 13, 372–373. [Google Scholar] [CrossRef] [PubMed]
- D’Arcy, A.; Chaillet, M.; Schiering, N.; Villard, F.; Lim, S.P.; Lefeuvre, P.; Erbel, P. Purification and crystallization of dengue and West Nile virus NS2B–NS3 complexes. Acta Crystallogr. Sect. F Struct. Biol. Cryst. Commun. 2006, 62, 157–162. [Google Scholar] [CrossRef] [Green Version]
- Aleshin, A.E.; Shiryaev, S.A.; Strongin, A.Y.; Liddington, R.C. Structural evidence for regulation and specificity of flaviviral proteases and evolution of the Flaviviridae fold. Protein Sci. 2007, 16, 795–806. [Google Scholar] [CrossRef] [Green Version]
- de la Cruz, L.; Nguyen, T.H.D.; Ozawa, K.; Shin, J.; Graham, B.; Huber, T.; Otting, G. Binding of low molecular weight inhibitors promotes large conformational changes in the dengue virus NS2B-NS3 protease: Fold analysis by pseudocontact shifts. J. Am. Chem. Soc. 2011, 133, 19205–19215. [Google Scholar] [CrossRef] [PubMed]
- Su, X.-C.; Ozawa, K.; Qi, R.; Vasudevan, S.; Lim, S.P.; Otting, G. NMR Analysis of the Dynamic Exchange of the NS2B Cofactor between Open and Closed Conformations of the West Nile Virus NS2B-NS3 Protease. PLoS Negl. Trop. Dis. 2009, 3, e561. [Google Scholar] [CrossRef] [PubMed]
- Su, X.-C.; Ozawa, K.; Yagi, H.; Lim, S.P.; Wen, D.; Ekonomiuk, D.; Huang, D.; Keller, T.H.; Sonntag, S.; Caflisch, A.; et al. NMR study of complexes between low molecular mass inhibitors and the West Nile virus NS2B-NS3 protease. FEBS J. 2009, 276, 4244–4255. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, Y.M.; Gayen, S.; Kang, C.; Joy, J.K.; Huang, Q.; Chen, A.S.; Wee, J.L.K.; Ang, M.J.Y.; Lim, H.A.; Hung, A.W.; et al. NMR Analysis of a Novel Enzymatically Active Unlinked Dengue NS2B-NS3 Protease Complex. J. Biol. Chem. 2013, 288, 12891–12900. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Phoo, W.W.; Li, Y.; Zhang, Z.; Lee, M.Y.; Loh, Y.R.; Tan, Y.B.; Ng, E.Y.; Lescar, J.; Kang, C.; Luo, D. Structure of the NS2B-NS3 protease from Zika virus after self-cleavage. Nat. Commun. 2016, 7, 13410. [Google Scholar] [CrossRef]
- Phoo, W.W.; El Sahili, A.; Zhang, Z.; Chen, M.W.; Liew, C.W.; Lescar, J.; Vasudevan, S.G.; Luo, D. Crystal structures of full length DENV4 NS2B-NS3 reveal the dynamic interaction between NS2B and NS3. Antivir. Res. 2020, 182, 104900. [Google Scholar] [CrossRef]
- Trnka, M.J.; Pellarin, R.; Robinson, P.J. Role of integrative structural biology in understanding transcriptional initiation. Methods 2019, 159–160, 4–22. [Google Scholar] [CrossRef] [PubMed]
- Alber, F.; Förster, F.; Korkin, D.; Topf, M.; Sali, A. Integrating Diverse Data for Structure Determination of Macromolecular Assemblies. Annu. Rev. Biochem. 2008, 77, 443–477. [Google Scholar] [CrossRef] [Green Version]
- Lasker, K.; Förster, F.; Bohn, S.; Walzthoeni, T.; Villa, E.; Unverdorben, P.; Beck, F.; Aebersold, R.; Sali, A.; Baumeister, W. Molecular architecture of the 26S proteasome holocomplex determined by an integrative approach. Proc. Natl. Acad. Sci. USA 2012, 109, 1380–1387. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Z.; Li, Y.; Loh, Y.R.; Phoo, W.W.; Hung, A.W.; Kang, C.; Luo, D. Crystal structure of unlinked NS2B-NS3 protease from Zika virus. Science 2016, 354, 1597–1600. [Google Scholar] [CrossRef]
- Maus, H.; Barthels, F.; Hammerschmidt, S.J.; Kopp, K.; Millies, B.; Gellert, A.; Ruggieri, A.; Schirmeister, T. SAR of novel benzothiazoles targeting an allosteric pocket of DENV and ZIKV NS2B/NS3 proteases. Bioorganic Med. Chem. 2021, 47, 116392. [Google Scholar] [CrossRef] [PubMed]
- Luo, D.; Xu, T.; Hunke, C.; Grüber, G.; Vasudevan, S.; Lescar, J. Crystal Structure of the NS3 Protease-Helicase from Dengue Virus. J. Virol. 2008, 82, 173–183. [Google Scholar] [CrossRef] [Green Version]
- Bera, A.K.; Kuhn, R.J.; Smith, J.L. Functional Characterization of cis and trans Activity of the Flavivirus NS2B-NS3 Protease. J. Biol. Chem. 2007, 282, 12883–12892. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Constant, D.A.; Mateo, R.; Nagamine, C.M.; Kirkegaard, K. Targeting intramolecular proteinase NS2B/3 cleavages for trans-dominant inhibition of dengue virus. Proc. Natl. Acad. Sci. USA 2018, 115, 10136–10141. [Google Scholar] [CrossRef] [Green Version]
- Kronenberger, T.; Serafim, M.S.M.; Tonduru, A.K.; Maltarollo, V.G.; Poso, A. Ligand Accessibility Insights to the Dengue Virus NS3-NS2B Protease Assessed by Long-Timescale Molecular Dynamics Simulations. ChemMedChem 2021, 16, 2524–2534. [Google Scholar] [CrossRef] [PubMed]
- Evans, P.R.; Murshudov, G.N. How good are my data and what is the resolution? Acta Crystallogr. D Biol. Crystallogr. 2013, 69, 1204–1214. [Google Scholar] [CrossRef] [PubMed]
- Adams, P.D.; Afonine, P.V.; Bunkóczi, G.; Chen, V.B.; Davis, I.W.; Echols, N.; Headd, J.J.; Hung, L.; Kapral, G.J.; Grosse-Kunstleve, R.W.; et al. PHENIX: A comprehensive Python-based system for macromolecular structure solution. Acta Crystallogr. D Biol. Crystallogr. 2010, 66, 213–221. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Afonine, P.V.; Grosse-Kunstleve, R.W.; Echols, N.; Headd, J.J.; Moriarty, N.W.; Mustyakimov, M.; Terwilliger, T.C.; Urzhumtsev, A.; Zwart, P.H.; Adams, P.D. Towards automated crystallographic structure refinement with phenix.refine. Acta Crystallogr. D Biol. Crystallogr. 2012, 68, 352–367. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Emsley, P.; Cowtan, K. Coot: Model-building tools for molecular graphics. Acta Crystallogr. Sect. D Struct. Biol. Crystallogr. 2004, 60, 2126–2132. [Google Scholar] [CrossRef] [Green Version]
- Liu, F.; Lössl, P.; Scheltema, R.; Viner, R.; Heck, A.J.R. Optimized fragmentation schemes and data analysis strategies for proteome-wide cross-link identification. Nat. Commun. 2017, 8, 15473. [Google Scholar] [CrossRef]
- Ser, Z.; Cifani, P.; Kentsis, A. Optimized Cross-Linking Mass Spectrometry for in Situ Interaction Proteomics. J. Proteome Res. 2019, 18, 2545–2558. [Google Scholar] [CrossRef] [PubMed]
- Lu, L.; Millikin, R.J.; Solntsev, S.K.; Rolfs, Z.; Scalf, M.; Shortreed, M.R.; Smith, L.M. Identification of MS-Cleavable and Noncleavable Chemically Cross-Linked Peptides with MetaMorpheus. J. Proteome Res. 2018, 17, 2370–2376. [Google Scholar] [CrossRef] [PubMed]
- Oliveros, J.C. (2007–2015). Chap. Available online: https://bioinfogp.cnb.csic.es/tools/venny/index.html (accessed on 20 April 2022).
- Kosinski, J.; von Appen, A.; Ori, A.; Karius, K.; Müller, C.; Beck, M. Xlink Analyzer: Software for analysis and visualization of cross-linking data in the context of three-dimensional structures. J. Struct. Biol. 2015, 189, 177–183. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pettersen, E.F.; Goddard, T.D.; Huang, C.C.; Couch, G.S.; Greenblatt, D.M.; Meng, E.C.; Ferrin, T.E. UCSF Chimera--a visualization system for exploratory research and analysis. J. Comput. Chem. 2004, 25, 1605–1612. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Okuda, S.; Watanabe, Y.; Moriya, Y.; Kawano, S.; Yamamoto, T.; Matsumoto, M.; Takami, T.; Kobayashi, D.; Araki, N.; Yoshizawa, A.C.; et al. jPOSTrepo: An international standard data repository for proteomes. Nucleic Acids Res. 2017, 45, D1107–D1111. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jumper, J.; Evans, R.; Pritzel, A.; Green, T.; Figurnov, M.; Ronneberger, O.; Tunyasuvunakool, K.; Bates, R.; Žídek, A.; Potapenko, A.; et al. Highly accurate protein structure prediction with AlphaFold. Nature 2021, 596, 583–589. [Google Scholar] [CrossRef] [PubMed]
- Mirdita, M.; Ovchinnikov, S.; Steinegger, M. ColabFold-Making protein folding accessible to all. Nat. Methods 2022, 19, 679–682. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Data Collection Statistics | PDB ID: 7VMV |
|---|---|
| Wavelength (Å) | 1.0 |
| Resolution range (Å) | 43.330–3.351 (3.471–3.351) |
| Space group | F 2 3 |
| Unit cell a, b, c α, β, γ, (Å) (o) | 259.984 259.984 259.984 90 90 90 |
| Total reflections | 36,553 (3535) |
| Unique reflections | 20,533 (2084) |
| Multiplicity | 1.8 (1.7) |
| Completeness (%) | 97.65 (99.43) |
| Mean I/sigma (I) | 10.00 (1.36) |
| Wilson B-factor (Å2) | 123.70 |
| a Rmerge | 0.04594 (0.5728) |
| R-meas | 0.06497 (0.8101) |
| R-pim | 0.04594 (0.5728) |
| CC1/2 | 0.998 (0.512) |
| CC * | 1 (0.823) |
| Refinement statistics | |
| Reflections used in refinement | 20,504 (2079) |
| Reflections used for R-free | 988 (121) |
| b Rwork | 0.2258 (0.3606) |
| c Rfree | 0.2628 (0.3576) |
| Number of non-hydrogen atoms | 6046 |
| macromolecules | 6046 |
| Protein residues | 852 |
| d RMSD (bonds) (Å) | 0.004 |
| RMSD (angles) (o) | 0.98 |
| Ramachandran favored (%) | 92.70 |
| Ramachandran allowed (%) | 7.18 |
| Ramachandran outliers (%) | 0.12 |
| Rotamer outliers (%) | 3.63 |
| Clashscore | 9.22 |
| Average B-factor | 138.53 |
| macromolecules | 138.53 |
| Number of TLS groups | 8 |
| Target | Purpose | Orientation | Sequence |
|---|---|---|---|
| eNS2B47NS3Pro | S135A Mutation | Forward | ACCCGGAACGGCGGGCTCTCCTA |
| Reverse | TTGAAATCCAGAGTTACTGCTCCAATTTCTC | ||
| eNS2B47NS3Pro | Linearisation | Forward | TCAGGAGCTCTGTGGGACG |
| Reverse | TCTTTGTGTTTTCACTTGTGC | ||
| eNS2B47NS3Pro | Fragment Insertion | Forward | GTGAAAACACAAAGATAAGAATTCTTGTACACGGCCGCATAATCGAAATTAATACGACTCACTATAGGGGAATTGTGAGCGGATAACAATTCCCCATCTTAGTATATTAGTTAAGTATAAGAAGGAGATATACATATGTCAGGAGCTCTGTGG |
| Reverse | CCACAGAGCTCCTGACATATGTATATCTCCTTCTTATACTTAACTAATATACTAAGATGGGGAATTGTTATCCGCTCACAATTCCCCTATAGTGAGTCGTATTAATTTCGATTATGCGGCCGTGTACAAGAATTCTTATCTTTGTGTTTTCAC |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Quek, J.-P.; Ser, Z.; Chew, B.L.A.; Li, X.; Wang, L.; Sobota, R.M.; Luo, D.; Phoo, W.W. Dynamic Interactions of Post Cleaved NS2B Cofactor and NS3 Protease Identified by Integrative Structural Approaches. Viruses 2022, 14, 1440. https://doi.org/10.3390/v14071440
Quek J-P, Ser Z, Chew BLA, Li X, Wang L, Sobota RM, Luo D, Phoo WW. Dynamic Interactions of Post Cleaved NS2B Cofactor and NS3 Protease Identified by Integrative Structural Approaches. Viruses. 2022; 14(7):1440. https://doi.org/10.3390/v14071440
Chicago/Turabian StyleQuek, Jun-Ping, Zheng Ser, Bing Liang Alvin Chew, Xin Li, Lili Wang, Radoslaw M. Sobota, Dahai Luo, and Wint Wint Phoo. 2022. "Dynamic Interactions of Post Cleaved NS2B Cofactor and NS3 Protease Identified by Integrative Structural Approaches" Viruses 14, no. 7: 1440. https://doi.org/10.3390/v14071440