
A Reduced Starch Level in Plants at Early Stages of Infection by Viruses Can Be Considered a Broad-Range Indicator of Virus Presence

 , ,
, , {kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Plant and Virus Materials
2.2. Virus Inoculation and Agro-Infiltration
2.3. Sample Preparation for Starch Purification and Nucleic Acid Extraction
2.4. RNA Extraction and RT-PCR
2.5. DNA Preparation and PCR
2.6. Calibration of Starch Standard Curve
2.7. Starch Purification and Quantification
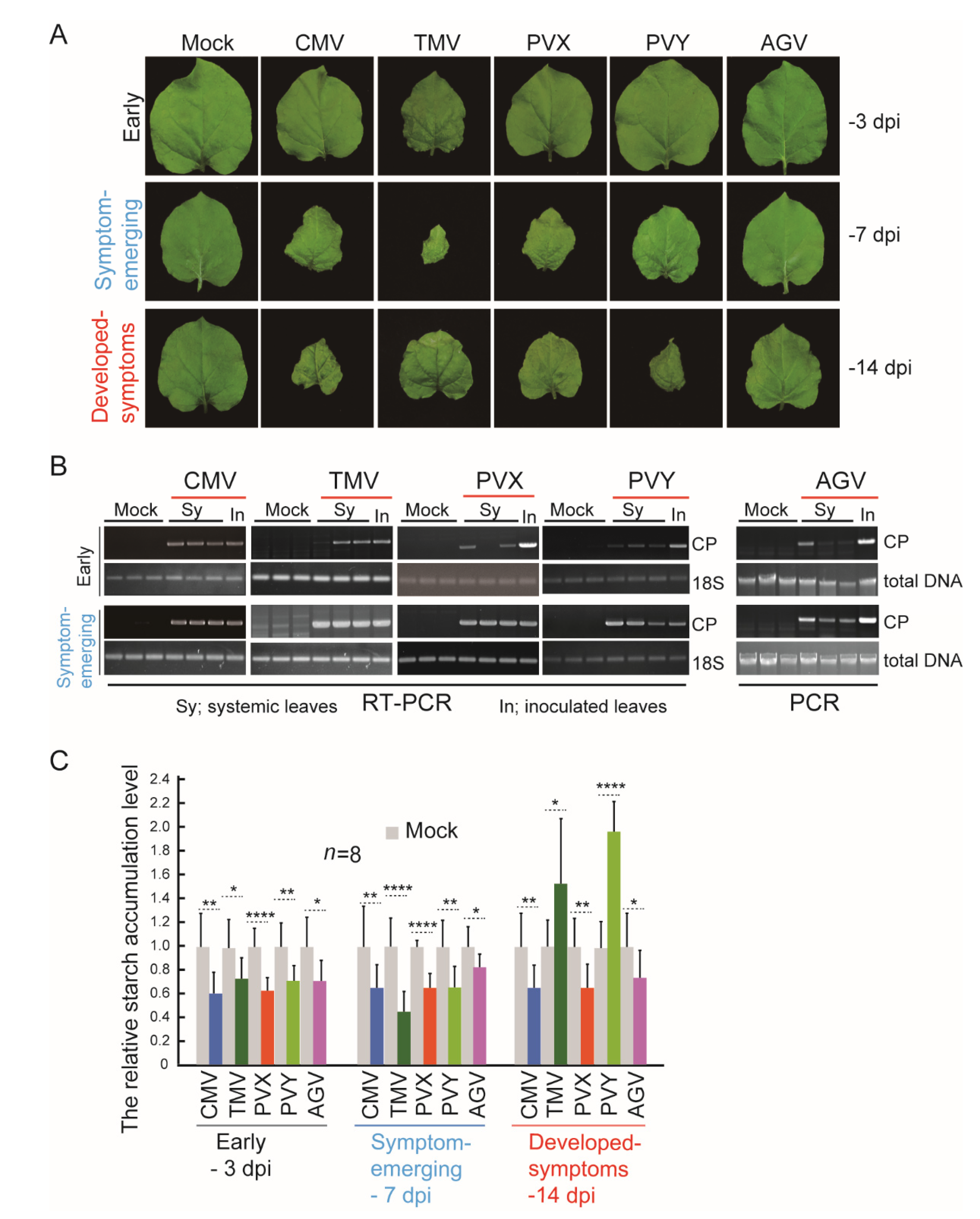
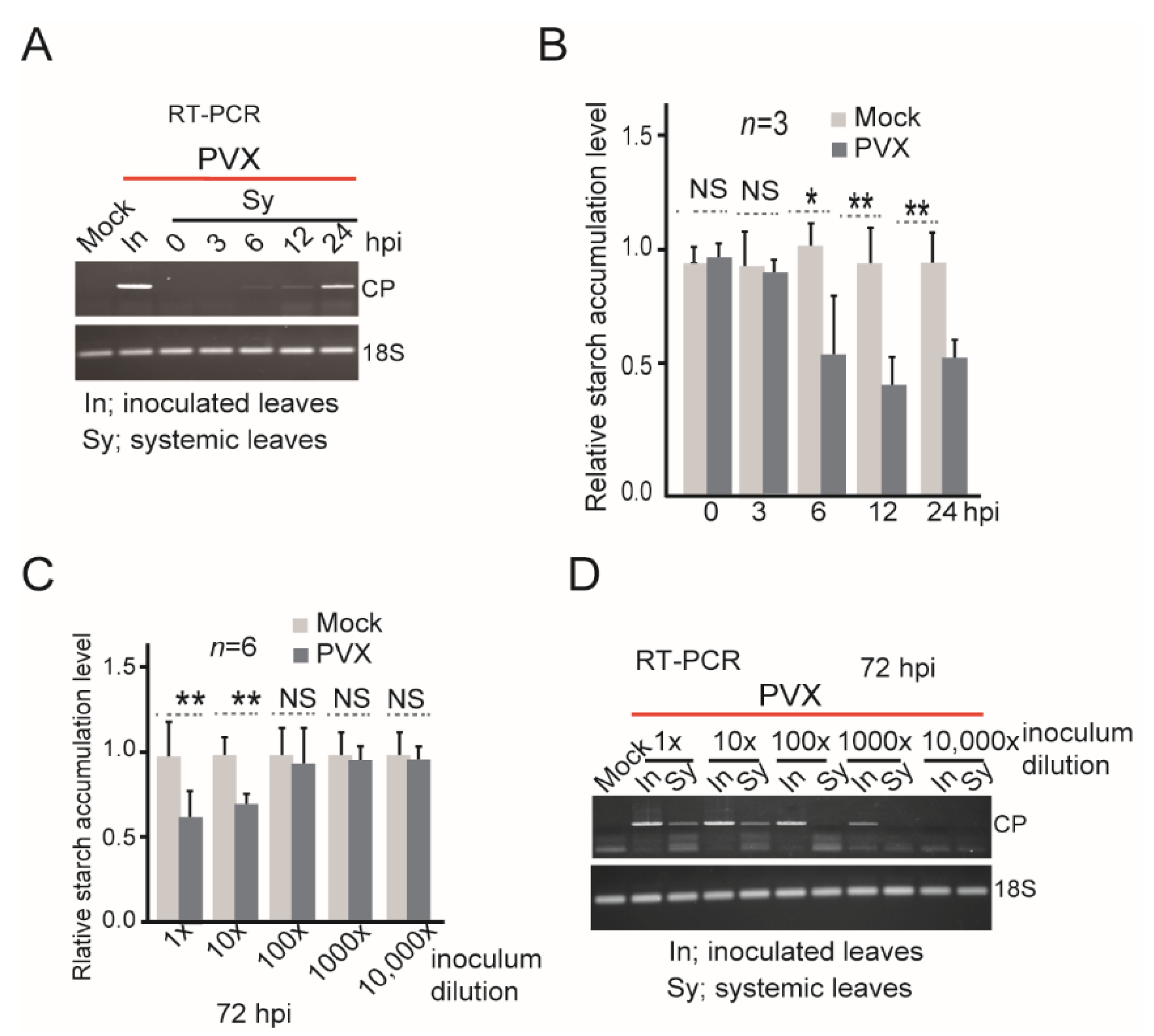
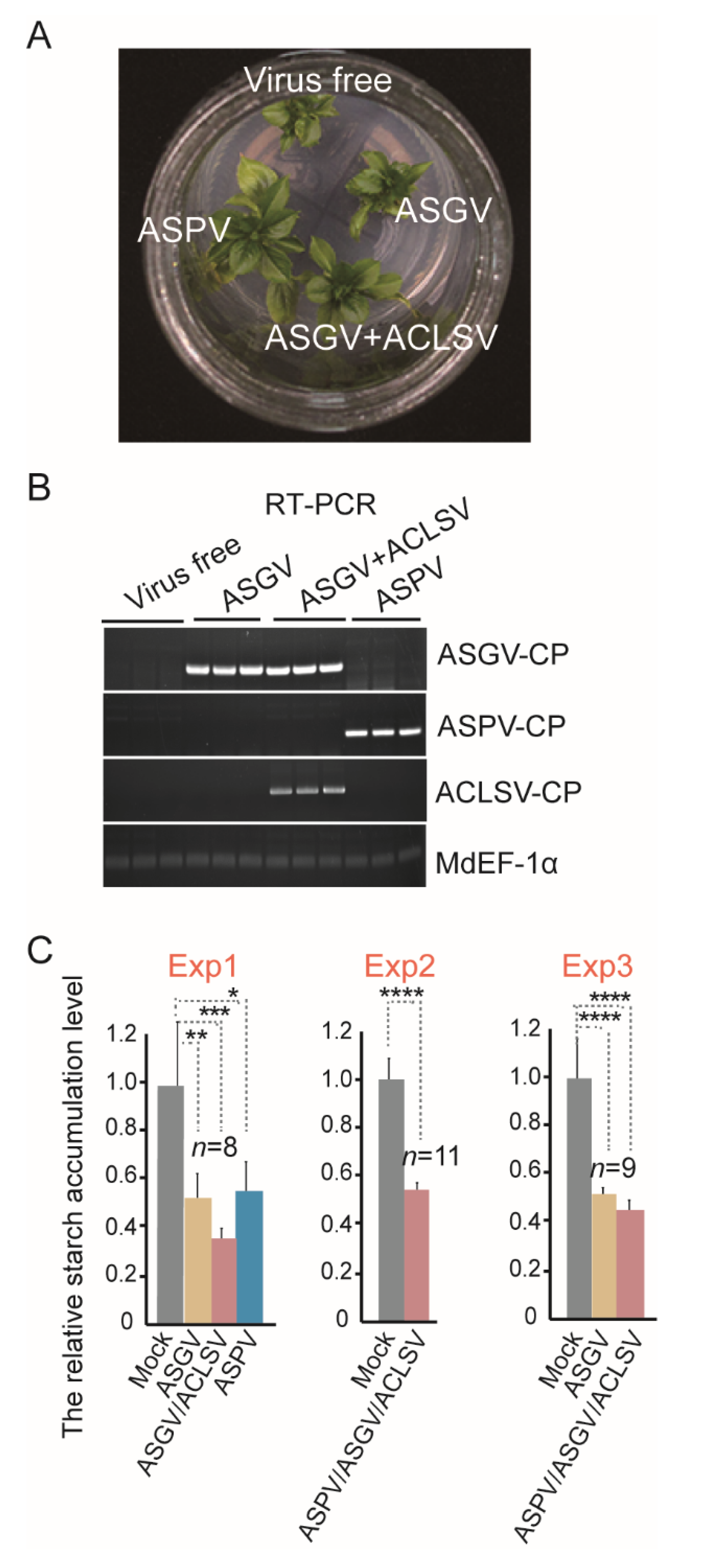
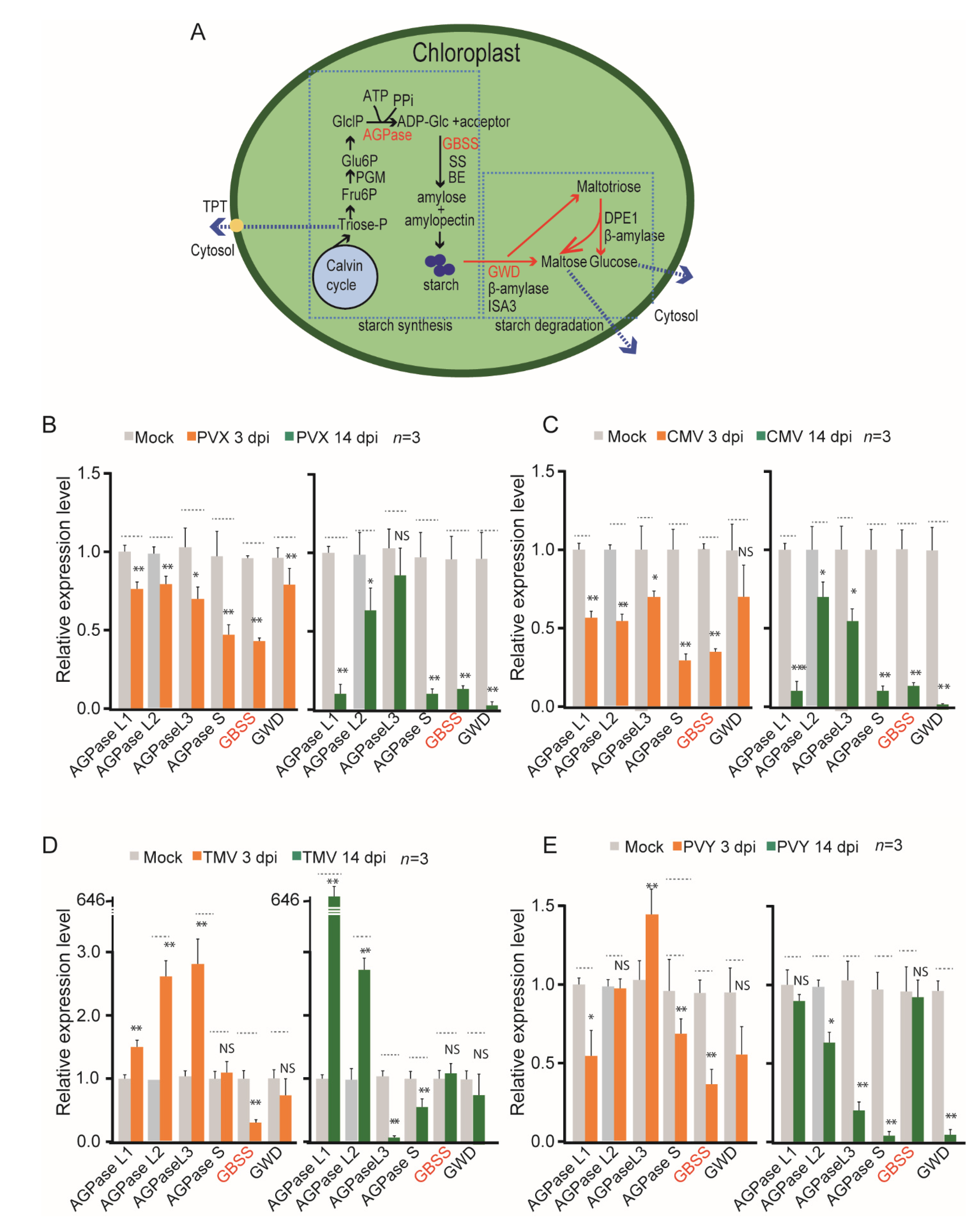
3. Results and Discussion
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Jones, R.A. Global plant virus disease pandemics and epidemics. Plants 2021, 10, 233. [Google Scholar] [CrossRef]
- Jeong, J.J.; Ju, H.J.; Noh, J. A Review of Detection Methods for the Plant Viruses. Res. Plant Dis. 2014, 20, 173–181. [Google Scholar] [CrossRef]
- Wang, M.-R.; Cui, Z.-H.; Li, J.-W.; Hao, X.-Y.; Zhao, L.; Wang, Q.-C. In vitro thermotherapy-based methods for plant virus eradication. Plant Methods 2018, 14, 87. [Google Scholar] [CrossRef] [Green Version]
- Davey, N.E.; Travé, G.; Gibson, T.J. How viruses hijack cell regulation. Trends Biochem. Sci. 2011, 36, 159–169. [Google Scholar] [CrossRef]
- Spriggs, C.C.; Harwood, M.C.; Tsai, B. How non-enveloped viruses hijack host machineries to cause infection. Adv. Virus Res. 2019, 104, 97–122. [Google Scholar]
- Mayer, K.A.; Stöckl, J.; Zlabinger, G.J.; Gualdoni, G.A. Hijacking the Supplies: Metabolism as a Novel Facet of Virus-Host Interaction. Front. Immunol. 2019, 10, 1533. [Google Scholar] [CrossRef]
- Trethewey, R.N.; Smith, A.M. Starch Metabolism in Leaves; Springer: Dordrecht, The Netherlands, 2006. [Google Scholar]
- Roberts, P.á.; Wood, K. Effects of a severe (P6) and a mild (W) strain of cucumber mosaic virus on tobacco leaf chlorophyll, starch and cell ultrastructure. Physiol. Plant Pathol. 1982, 21, 31–37. [Google Scholar] [CrossRef]
- Fraser, L.; Keeling, J.; Matthews, R. A reduction in starch accumulation in the apical dome of Chinese cabbage seedlings following inoculation with turnip yellow mosaic virus. Physiol. Plant Pathol. 1984, 24, 157–162. [Google Scholar] [CrossRef]
- Tecsi, L.; Wang, D.; Smith, A.; Leegood, R.C.; Maule, A. Red clover mottle virus infection affects sink-source relationships and starch accumulation in pea plants. J. Exp. Bot. 1992, 43, 1409–1412. [Google Scholar] [CrossRef]
- Zechmann, B.; Müller, M.; Zellnig, G. Cytological modifications in zucchini yellow mosaic virus (ZYMV)-infected Styrian pumpkin plants. Arch. Virol. 2003, 148, 1119–1133. [Google Scholar] [CrossRef]
- Técsi, L.I.; Maule, A.J.; Smith, A.M.; Leegood, R.C. Metabolic alterations in cotyledons of Cucurbita pepo infected by cucumber mosaic virus. J. Exp. Bot. 1994, 45, 1541–1551. [Google Scholar] [CrossRef]
- Li, K.; Wei, Y.-H.; Wang, R.-H.; Mao, J.-P.; Tian, H.-Y.; Chen, S.-Y.; Li, S.-H.; Tahir, M.-M.; Zhang, D. Mdm-MIR393b-mediated adventitious root formation by targeted regulation of MdTIR1A expression and weakened sensitivity to auxin in apple rootstock. Plant Sci. 2021, 308, 110909. [Google Scholar] [CrossRef]
- Li, B.; Feng, C.; Wang, M.; Hu, L.; Chen, L.; Wang, Q. Cryopreservation of shoot tips of apple (Malus) by encapsulation-dehydration using adventitious shoots derived from leaf segments. Vitr. Cell. Dev. Biol. Plant 2014, 50, 357–368. [Google Scholar] [CrossRef]
- Liang, P.; Navarro, B.; Zhang, Z.; Wang, H.; Lu, M.; Xiao, H.; Wu, Q.; Zhou, X.; Di Serio, F.; Li, S. Identification and characterization of a novel geminivirus with a monopartite genome infecting apple trees. J. Gen. Virol. 2015, 96, 2411–2420. [Google Scholar] [CrossRef]
- Sun, L.; Andika, I.B.; Shen, J.; Yang, D.; Chen, J. The P 2 of W heat yellow mosaic virus rearranges the endoplasmic reticulum and recruits other viral proteins into replication-associated inclusion bodies. Mol. Plant Pathol. 2014, 15, 466–478. [Google Scholar] [CrossRef]
- Liu, D.; Shi, L.; Han, C.; Yu, J.; Li, D.; Zhang, Y. Validation of reference genes for gene expression studies in virus-infected Nicotiana benthamiana using quantitative real-time PCR. PLoS ONE 2012, 7, e46451. [Google Scholar] [CrossRef]
- Murray, M.; Thompson, W. Rapid isolation of high molecular weight plant DNA. Nucleic Acids Res. 1980, 8, 4321–4326. [Google Scholar] [CrossRef] [Green Version]
- Nakamura, S.; Satoh, H.; Ohtsubo, K. Development of formulae for estimating amylose content, amylopectin chain length distribution, and resistant starch content based on the iodine absorption curve of rice starch. Biosci. Biotechnol. Biochem. 2015, 79, 443–455. [Google Scholar] [CrossRef] [Green Version]
- McGrance, S.J.; Cornell, H.J.; Rix, C.J. A simple and rapid colorimetric method for the determination of amylose in starch products. Starch-Stärke 1998, 50, 158–163. [Google Scholar] [CrossRef]
- Zhao, J.; Zhang, X.; Hong, Y.; Liu, Y. Chloroplast in Plant-Virus Interaction. Front. Microbiol. 2016, 7, 1565. [Google Scholar] [CrossRef] [Green Version]
- Fernandez, O.; Ishihara, H.; George, G.M.; Mengin, V.; Flis, A.; Sumner, D.; Arrivault, S.; Feil, R.; Lunn, J.E.; Zeeman, S.C.; et al. Leaf Starch Turnover Occurs in Long Days and in Falling Light at the End of the Day. Plant Physiol. 2017, 174, 2199–2212. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, S.; Li, Y. Current understanding of the interplays between host hormones and plant viral infections. PLoS Pathog. 2021, 17, e1009242. [Google Scholar] [CrossRef] [PubMed]
- Thalmann, M.; Pazmino, D.; Seung, D.; Horrer, D.; Nigro, A.; Meier, T.; Kölling, K.; Pfeifhofer, H.W.; Zeeman, S.C.; Santelia, D. Regulation of leaf starch degradation by abscisic acid is important for osmotic stress tolerance in plants. Plant Cell 2016, 28, 1860–1878. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xing, F.; Robe, B.L.; Zhang, Z.; Wang, H.; Li, S. Genomic Analysis, Sequence Diversity, and Occurrence of Apple necrotic mosaic virus, a Novel Ilarvirus Associated with Mosaic Disease of Apple Trees in China. Plant Dis. 2018, 102, 1841–1847. [Google Scholar] [CrossRef] [Green Version]
- Orzechowski, S. Starch metabolism in leaves. Acta Biochim. Pol. 2008, 55, 435–445. [Google Scholar] [CrossRef]
- Pfister, B.; Zeeman, S.C. Formation of starch in plant cells. Cell. Mol. Life Sci. 2016, 73, 2781–2807. [Google Scholar] [CrossRef] [Green Version]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhao, W.; Wang, L.; Liu, M.; Zhang, D.; Andika, I.B.; Zhu, Y.; Sun, L. A Reduced Starch Level in Plants at Early Stages of Infection by Viruses Can Be Considered a Broad-Range Indicator of Virus Presence. Viruses 2022, 14, 1176. https://doi.org/10.3390/v14061176
Zhao W, Wang L, Liu M, Zhang D, Andika IB, Zhu Y, Sun L. A Reduced Starch Level in Plants at Early Stages of Infection by Viruses Can Be Considered a Broad-Range Indicator of Virus Presence. Viruses. 2022; 14(6):1176. https://doi.org/10.3390/v14061176
Chicago/Turabian StyleZhao, Wanying, Li Wang, Meizi Liu, Dong Zhang, Ida Bagus Andika, Ying Zhu, and Liying Sun. 2022. "A Reduced Starch Level in Plants at Early Stages of Infection by Viruses Can Be Considered a Broad-Range Indicator of Virus Presence" Viruses 14, no. 6: 1176. https://doi.org/10.3390/v14061176