
Transmission of Grapevine Red Blotch Virus by Spissistilus festinus [Say, 1830] (Hemiptera: Membracidae) between Free-Living Vines and Vitis vinifera ‘Cabernet Franc’

 , , , and
, , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials
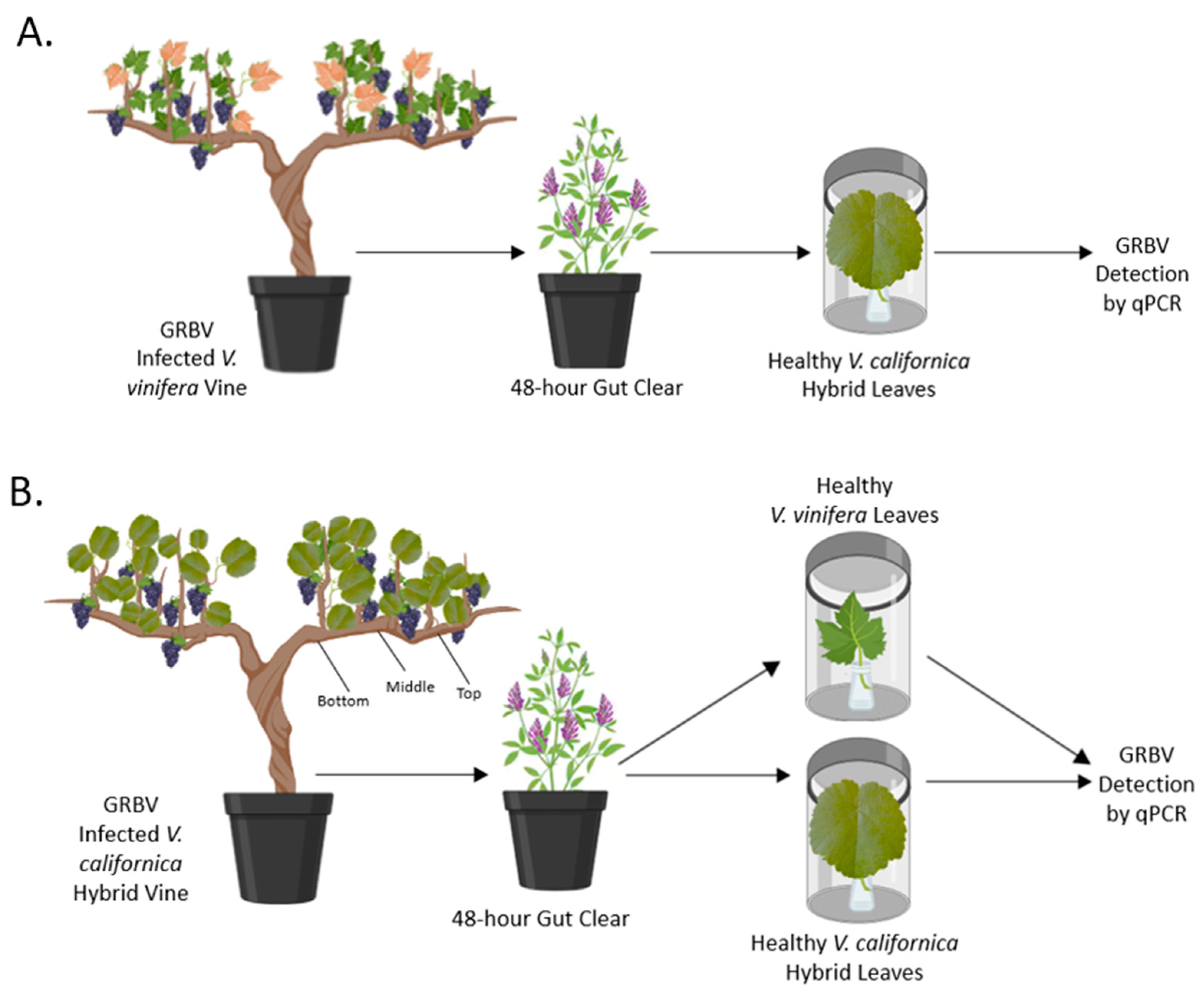
2.2. Transmission Assays of GRBV by S. festinus
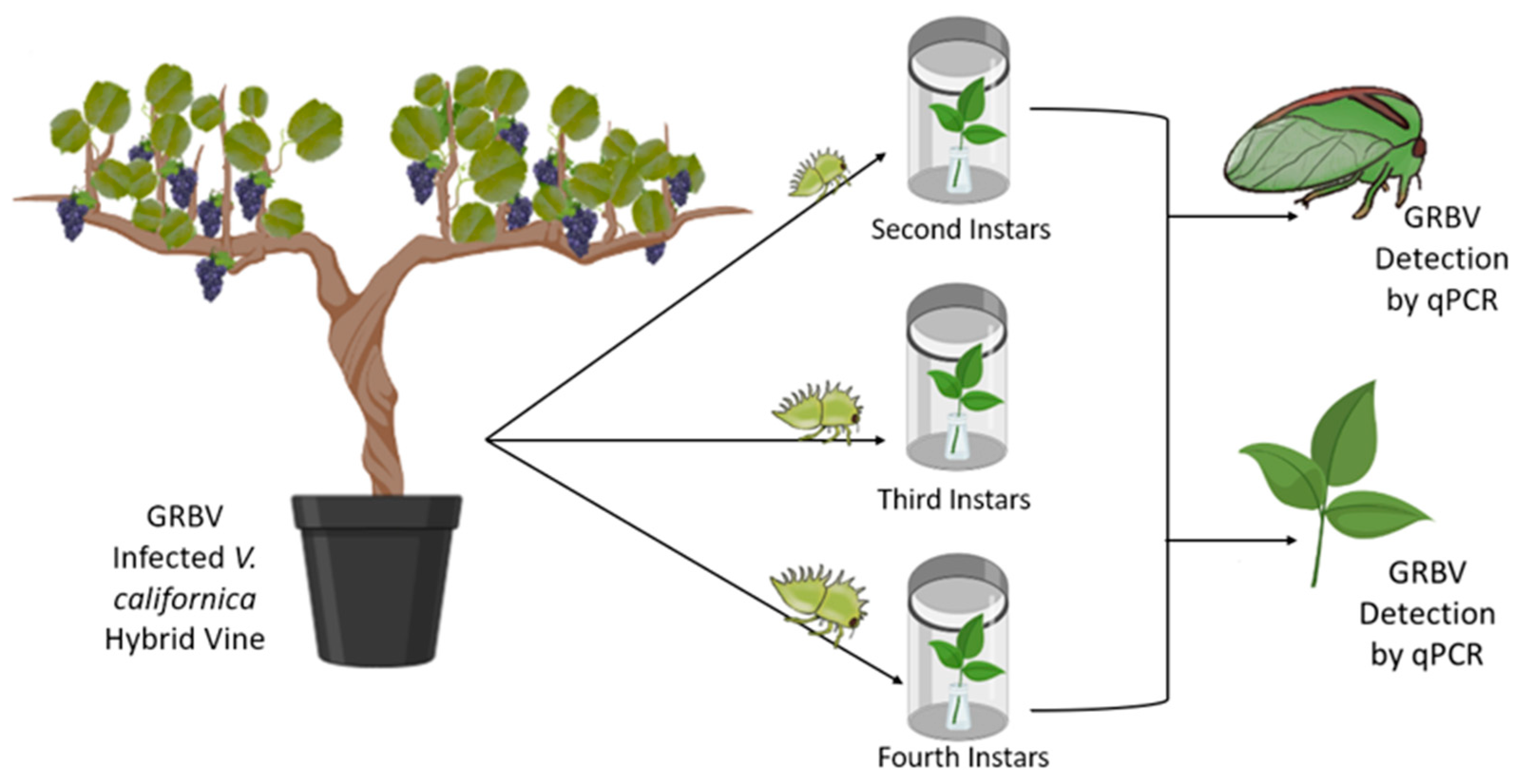
2.3. Trans-Stadial Transmission of GRBV by S. festinus
2.4. Assessing the Distribution of GRBV in Free-Living Vines
2.5. Nucleic Acid Extraction from Plant Tissue and S. festinus, and GRBV Detection by PCR and qPCR
2.6. Statistical Analyses
3. Results
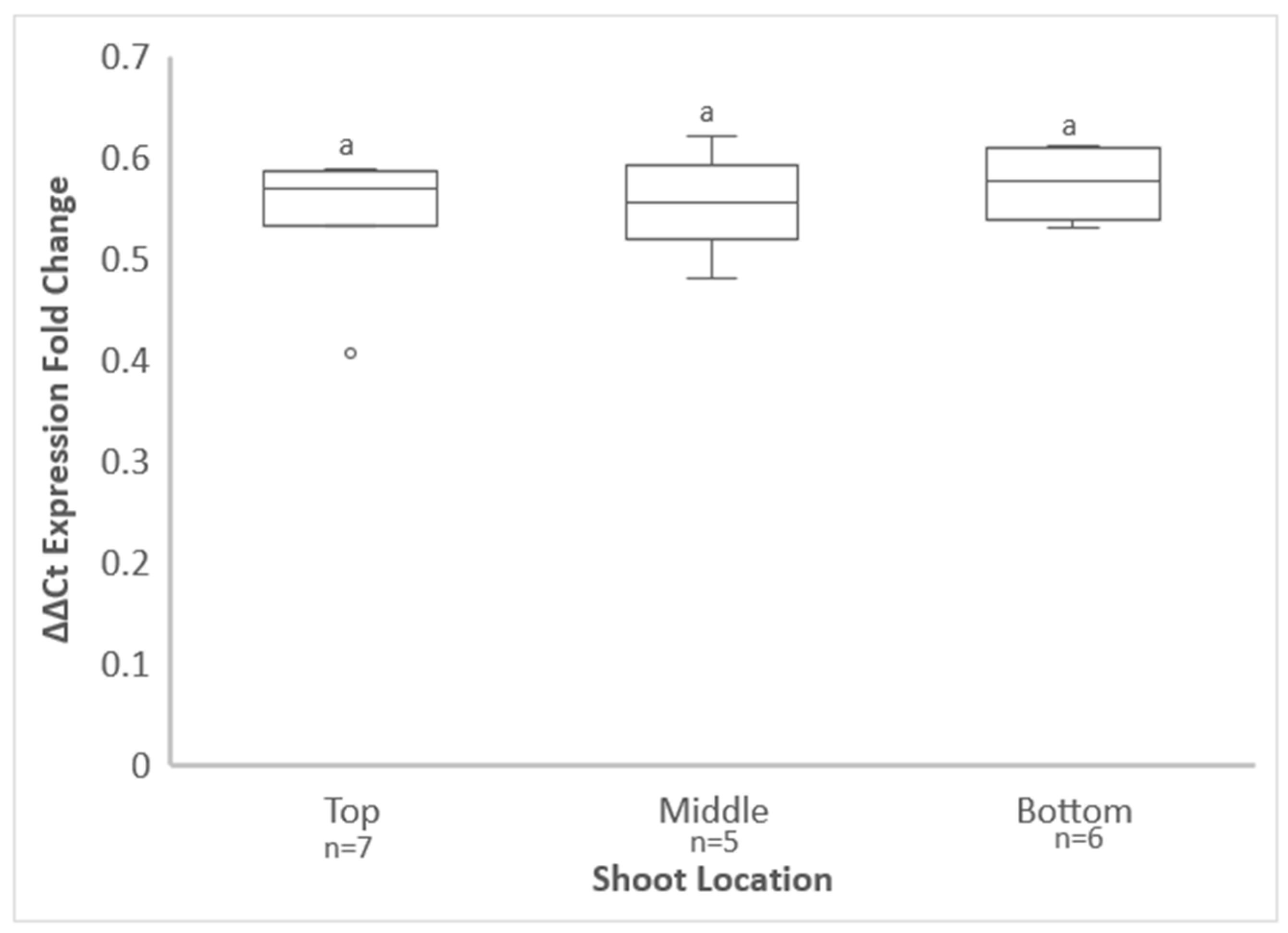
3.1. Identification of the Free-Living Vines Used in This Study and GRBV Distribution
3.2. GRBV Transmission by S. festinus from Infected Free-Living Vines
3.3. GRBV Transmission by S. festinus from Infected ‘Cabernet Franc’
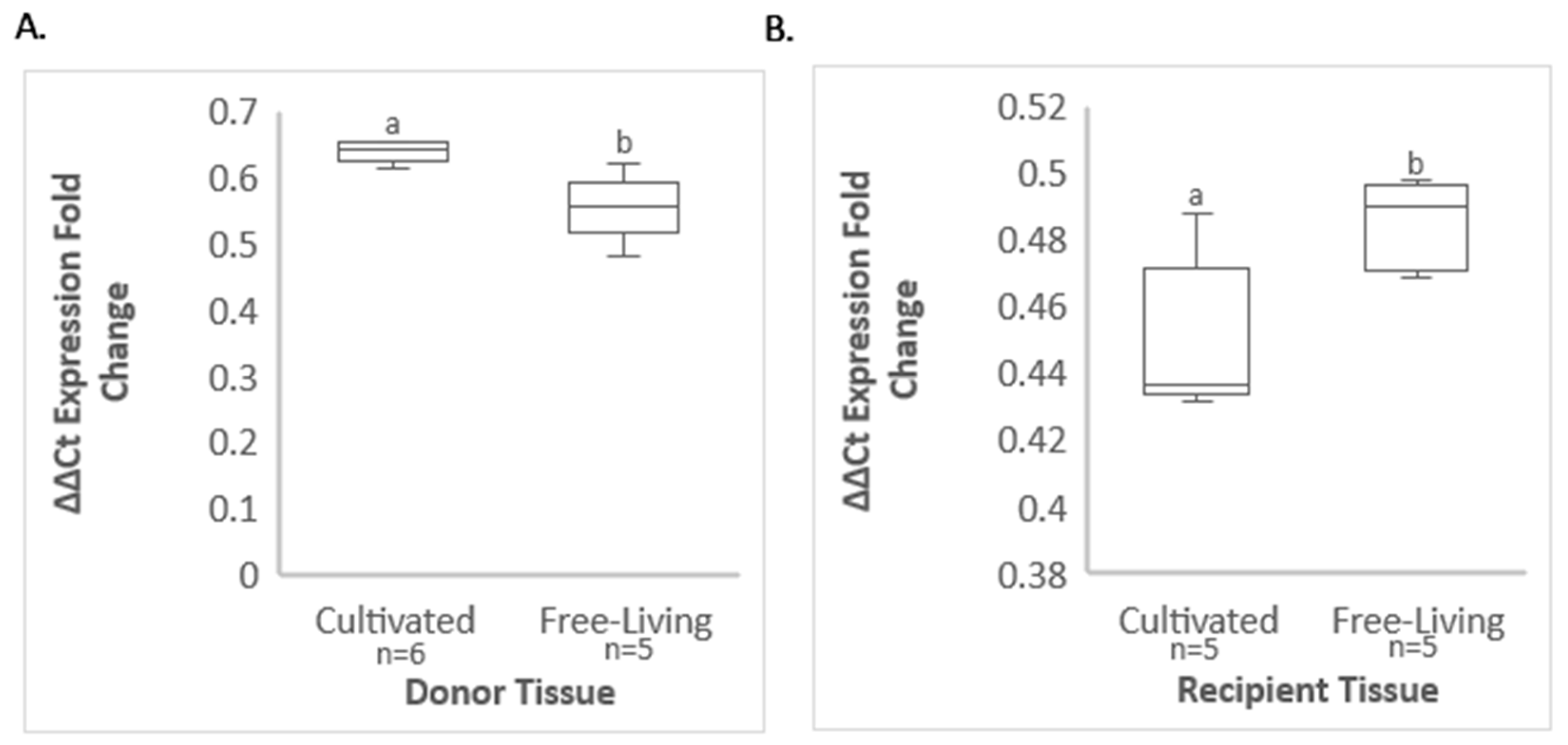
3.4. Comparative GRBV Titer in Infected Free-Living and ‘Cabernet Franc’ Vines
3.5. Trans-Stadial Transmission of GRBV by S. festinus Adults Derived from Instars That Emerged on Infected Free-Living Vines
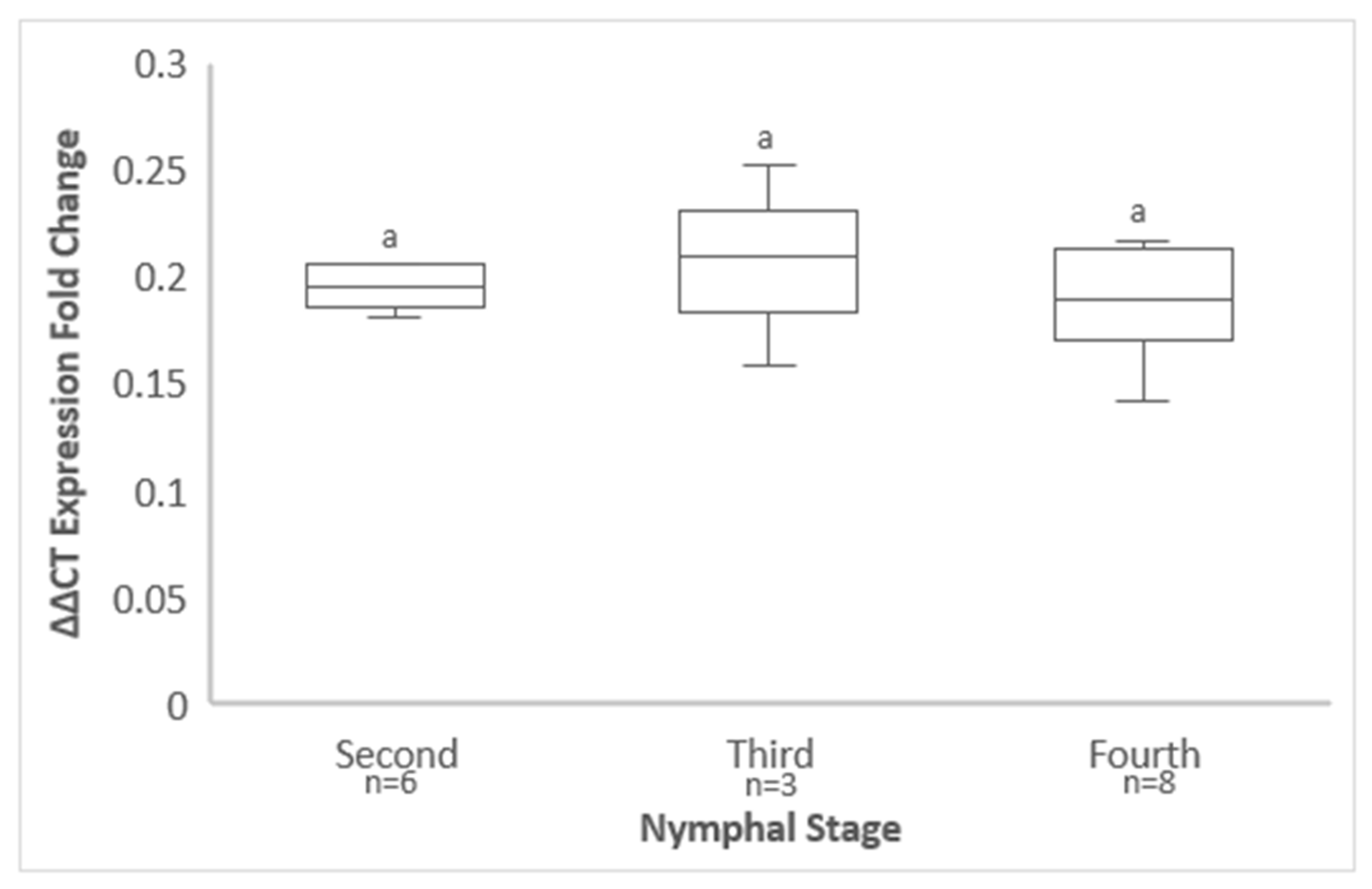
3.6. GRBV Titer in S. festinus Adults Derived from Instars That Emerged on Infected Free-Living Vines
3.7. Transmission of GRBV by S. festinus Adults Derived from Instars That Emerged on Infected Free-Living Vines
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Yepes, L.M.; Cieniewicz, E.; Krenz, B.; McLane, H.; Thompson, J.R.; Perry, K.; Fuchs, M. Causative Role of Grapevine Red Blotch Virus in Red Blotch Disease. Phytopathology 2018, 108, 902–909. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sudarshana, M.R.; Perry, K.L.; Fuchs, M.F. Grapevine red blotch-associated virus, an Emerging Threat to the Grapevine Industry. Phytopathology 2015, 105, 1026–1032. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cieniewicz, E.; Perry, K.; Fuchs, M. Grapevine Red Blotch: Molecular Biology of the Virus and Management of the Disease. In Grapevine Viruses: Molecular Biology, Diagnostics and Management; Meng, B., Martelli, G., Golino, D., Fuchs, M., Eds.; Springer: Cham, Switzerland, 2017; pp. 303–314. [Google Scholar]
- Cieniewicz, E.J.; Qui, W.; Saldarelli, P.; Fuchs, M. Believing is Seeing: Lessons from Emerging Viruses in Grapevine. J. Plant Pathol. 2020, 102, 619–632. [Google Scholar] [CrossRef]
- Rumbaugh, A.C.; Sudarshana, M.R.; Oberholster, A. Grapevine Red Blotch Disease Etiology and Its Impact on Grapevine Physiology and Berry and Wine Composition. Horticulturae 2021, 7, 552. [Google Scholar] [CrossRef]
- Blanco-Ulate, B.; Hopfer, H.; Figueroa-Balderas, R.; Ye, Z.; Rivero, R.M.; Albacete, A.; Pérez-Alfocea, F.; Koyama, R.; Anderson, M.M.; Smith, R.J.; et al. Red Blotch Disease Alters Grape Berry Development and Metabolism by Interfering with the Transcriptional and Hormonal Regulation of Ripening. J. Exp. Bot. 2017, 68, 1225–1238. [Google Scholar] [CrossRef] [Green Version]
- Martínez-Lüscher, J.; Plank, C.M.; Brillante, L.; Cooper, M.L.; Smith, R.J.; Al-Rwahnih, M.; Yu, R.; Oberholster, A.; Girardello, R.; Kurtural, S.K. Grapevine Red Blotch Virus May Reduce Carbon Translocation Leading to Impaired Grape Berry Ripening. J. Agric. Food Chem. 2019, 67, 2437–2448. [Google Scholar] [CrossRef]
- Ricketts, K.D.; Gómez, M.I.; Fuchs, M.F.; Martinson, T.E.; Smith, R.J.; Cooper, M.L.; Moyer, M.M.; Wise, A. Mitigating the Economic Impact of Grapevine Red Blotch: Optimizing Disease Management Strategies in U.S. Vineyards. Am. J. Enol. Vitic. 2017, 68, 127–135. [Google Scholar] [CrossRef]
- Krenz, B.; Thompson, J.R.; McLane, H.L.; Fuchs, M.; Perry, K.L. Grapevine red blotch-associated virus is Widespread in the United States. Phytopathology 2014, 104, 1232–1240. [Google Scholar] [CrossRef] [Green Version]
- Guo, T.W.; Vimalesvaran, D.; Thompson, J.R.; Perry, K.L.; Krenz, B. Subcellular Localization of Grapevine red blotch-associated virus ORFs V2 and V3. Virus Genes 2015, 51, 156–158. [Google Scholar] [CrossRef]
- Buchs, N.; Braga-Lagache, S.; Uldry, A.C.; Brodard, J.; Debonneville, C.; Reynard, J.S.; Heller, M. Absolute Quantification of Grapevine Red Blotch Virus in Grapevine Leaf and Petiole Tissues by Proteomics. Front. Plant Sci. 2018, 9, 1735. [Google Scholar] [CrossRef]
- Vargas-Asencio, J.; Liou, H.; Perry, K.L.; Thompson, J.R. Evidence for the Splicing of Grablovirus Transcripts Reveals a Putative Novel Open Reading Frame. J. Gen. Virol. 2019, 100, 709–720. [Google Scholar] [CrossRef] [PubMed]
- Adiputra, J.; Kesoju, S.R.; Naidu, R.A. The Relative Occurrence of Grapevine Leafroll-associated Virus 3 and Grapevine Red Blotch Virus in Washington State Vineyards. Plant Dis. 2018, 102, 2129–2135. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hoffmann, M.; Talton, W.; Nita, M.; Jones, T.; Al Rwahnih, M.; Sudarshana, R.; Almeyda, C. First Report of Grapevine red blotch virus, the Causal Agent of Grapevine Red Blotch Disease, in Vitis vinifera in North Carolina. Plant Dis. 2019, 104, 1266. [Google Scholar] [CrossRef]
- Soltani, N.; Hu, R.; Hensley, D.D.; Lockwood, D.L.; Perry, K.L.; Hajimorad, M.R. A Survey for Nine Major Viruses of Grapevines in Tennessee Vineyards. Plant Health Prog. 2021, 21, 157–161. [Google Scholar] [CrossRef]
- Poojari, S.; Lowery, D.T.; Rott, M.; Schmidt, A.M.; Úrbez-Torres, J.R. Incidence, Distribution and Genetic Diversity of Grapevine red blotch virus in British Columbia. Can. J. Plant Pathol. 2017, 39, 201–211. [Google Scholar] [CrossRef]
- Fall, M.L.; Xu, D.; Lemoyne, P.; Ben Moussa, I.E.; Beaulieu, C.; Carisse, O. A Diverse Virome of Leafroll-Infected Grapevine Unveiled by dsRNA Sequencing. Viruses 2020, 12, 1142. [Google Scholar] [CrossRef] [PubMed]
- Xiao, H.; Kim, W.S.; Meng, B. A Highly Effective and Versatile Technology for the Isolation of RNAs from Grapevines and Other Woody Perennials for use in Virus Diagnostics. Virol. J. 2015, 12, 171. [Google Scholar] [CrossRef] [Green Version]
- Reynard, J.-S.; Brodard, J.; Dubuis, N.; Zufferey, V.; Schumpp, O.; Schaerer, S.; Gugerli, P. Grapevine red blotch virus: Absence in Swiss Vineyards and Analysis of Potential Detrimental Effect on Viticultural Performance. Plant Dis. 2018, 102, 651–655. [Google Scholar] [CrossRef] [Green Version]
- Lim, S.; Igori, D.; Zhao, F.; Moon, J.S.; Cho, I.-S.; Choi, G.-S. First Report of Grapevine red blotch-associated virus on Grapevine in Korea. Plant Dis. 2016, 100, 1957. [Google Scholar] [CrossRef]
- Gasperin-Bulbarela, J.; Licea-Navarro, A.F.; Pino-Villar, C.; Hernández-Martínez, R.; Carillo-Tripp, J. First Report of Grapevine Red Blotch Virus in Mexico. Plant Dis. 2018, 103, 381. [Google Scholar] [CrossRef]
- Luna, F.; Debat, H.; Moyano, S.; Zavallo, D.; Asurmendi, S.; Gomez-Talquenca, S. First Report of Grapevine Red Blotch Virus Infecting Grapevine in Argentina. J. Plant Pathol. 2019, 101, 1239. [Google Scholar] [CrossRef] [Green Version]
- Marwal, A.; Kumar, R.; Paul Khurana, S.M.; Gaur, R.K. Complete Nucleotide Sequence of a New Geminivirus Isolated from Vitis vinifera in India: A Symptomless Host of Grapevine Red Blotch Virus. Virusdisease 2019, 30, 106–111. [Google Scholar] [CrossRef] [PubMed]
- Bertazzon, N.; Migliaro, D.; Rossa, A.; Filippin, L.; Casarin, S.; Giust, M.; Brancadoro, L.; Crespan, M.; Angelini, E. Grapevine Red Blotch Virus is Sporadically Present in a Germplasm Collection in Northern Italy. J. Plant Dis. Prot. 2021, 128, 1–5. [Google Scholar] [CrossRef]
- Reynard, J.-S.; Brodard, J.; Dubuis, N.; Kellenberger, I.; Spilmont, A.-S.; Roquis, D.; Maliogka, V.; Marchal, C.; Dedet, S.; Gning, O.; et al. Screening of Grapevine Red Blotch Virus in Two European Ampelographic Collections. J. Plant Pathol. 2022, 104, 9–15. [Google Scholar] [CrossRef]
- Cieniewicz, E.; Thompson, J.R.; McLane, H.; Perry, K.L.; Dangl, G.S.; Corbett, Q.; Martinson, T.; Wise, A.; Wallis, A.; O’Connell, J.; et al. Prevalence and Genetic Diversity of Grabloviruses in Free-Living Vitis spp. Plant Dis. 2018, 102, 2308–2316. [Google Scholar] [CrossRef] [Green Version]
- Cieniewicz, E.; Flasco, M.; Brunelli, M.; Onwumelu, A.; Wise, A.; Fuchs, M.F. Differential Spread of Grapevine Red Blotch Virus in California and New York Vineyards. Phytobiomes J. 2019, 3, 203–211. [Google Scholar] [CrossRef] [Green Version]
- Dalton, D.T.; Hilton, R.J.; Kaiser, C.; Daane, K.M.; Sudarshana, M.R.; Vo, J.; Zalom, F.G.; Buser, J.Z.; Walton, V.M. Spatial Associations of Vines Infected with Grapevine Red Blotch Virus in Oregon Vineyards. Plant Dis. 2019, 103, 1507–1514. [Google Scholar] [CrossRef]
- Achala, N.; Deshields, J.B.; Levin, A.D.; Hilton, R.; Rijal, J. Epidemiology of Grapevine Red Blotch Disease Progression in Southern Oregon Vineyards. Am. J. Enol. Vitic. 2022, 73, 116–124. [Google Scholar] [CrossRef]
- Bahder, B.W.; Zalom, F.G.; Sudarshana, M.R. An Evaluation of the Flora Adjacent to Wine Grape Vineyards for the Presence of Alternative Host Plants of Grapevine red blotch-associated virus. Plant Dis. 2016, 100, 1571–1574. [Google Scholar] [CrossRef] [Green Version]
- Flasco, M.; Hoyle, V.; Cieniewicz, E.J.; Roy, B.G.; McLane, H.L.; Perry, K.L.; Loeb, G.; Nault, B.; Heck, M.; Fuchs, M. Grapevine Red Blotch Virus Is Transmitted by the Three-Cornered Alfalfa Hopper in a Circulative, Nonpropagative Mode with Unique Attributes. Phytopathology 2021, 111, 1851–1861. [Google Scholar] [CrossRef]
- Cieniewicz, E.J.; Pethybridge, S.J.; Loeb, G.; Perry, K.; Fuchs, M. Insights into the Ecology of Grapevine red blotch virus in a Diseased Vineyard. Phytopathology 2018, 108, 94–102. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wilson, H.; Hogg, B.N.; Blaisdell, G.K.; Andersen, J.C.; Yazdani, A.S.; Billings, A.C.; Ooi, K.-I.M.; Soltani, N.; Almeida, R.P.P.; Cooper, M.L.; et al. Survey of Vineyard Insects and Plants to Identify Potential Insect Vectors and Noncrop Reservoirs of Grapevine Red Blotch Virus. PhytoFrontiers 2022, 2, 66–73. [Google Scholar] [CrossRef]
- Schoelz, J.; Volenberg, D.; Adhab, M.; Fang, Z.; Klassen, V.; Spinka, C.; Al Rwahnih, M. A Survey of Viruses Found in Grapevine Cultivars Grown in Missouri. Am. J. Enol. Vitic. 2021, 72, 73–84. [Google Scholar] [CrossRef]
- Perry, K.L.; McLane, H.; Hyder, M.Z.; Dangl, G.S.; Thompson, J.R.; Fuchs, M.F. Grapevine red blotch-associated virus is Present in Free-Living Vitis spp. Proximal to Cultivated Grapevine. Virology 2016, 106, 663–670. [Google Scholar] [CrossRef] [Green Version]
- Dangl, G.S.; Mendum, M.L.; Yang, J.; Walker, M.A.; Preece, J.E. Hybridization of Cultivated Vitis vinifera with Wild V. californica and V. girdiana in California. Ecol. Evol. 2015, 5, 5671–5684. [Google Scholar] [CrossRef]
- Setiono, F.J.; Chatterjee, D.; Fuchs, M.; Perry, K.L.; Thompson, J.R. The Distribution and Detection of Grapevine red blotch virus in its Host Depend on Time of Sampling and Tissue Type. Plant Dis. 2018, 102, 2187–2193. [Google Scholar] [CrossRef] [Green Version]
- Pereira, W.J.; Bassinello, P.Z.; Brondani, C.; Vianello, R.P. An Improved Method for RNA Extraction from Common Bean Seeds and Validation of Reference Genes for qPCR. Crop Breed. App. Biotechnol. 2017, 17, 150–158. [Google Scholar] [CrossRef]
- Klaassen, V.A.; Sim, S.T.; Dangl, G.S.; Osman, F.; AL Rwahnih, M.; Rowhani, A.; Golino, D.A. Vitis californica and Vitis californica × Vitis vinifera Hybrids are Hosts for Grapevine leafroll-associated virus-2 and -3 and Grapevine virus A and B. Plant Dis. 2011, 95, 657–665. [Google Scholar] [CrossRef] [Green Version]
- Diaz-Lara, A.; Dangle, G.S.; Yang, J.; Yang, J.; Golina, D.; Al Rwahnih, M. Identification of Grapevine Pinot gris virus in Free-Living Vitis spp. Located in Riparian Areas Adjacent to Commercial Vineyards. Plant Dis. 2021, 105, 2295–2298. [Google Scholar] [CrossRef]
- Reynard, J.S.; Brodard, J.; Remoliff, E.; Lefebvre, M.; Schumpp, O.; Candresse, T. A Novel Foveavirus Identified in Wild Grapevine (Vitis vinifera subsp. sylvestris). Arch. Virol. 2020, 165, 2999–3002. [Google Scholar] [CrossRef]
- Sabella, E.; Pierro, R.; Luvisi, A.; Panattoni, A.; D’Onofrio, C.; Scalabrelli, G.; Nutricati, E.; Aprile, A.; De Bellis, L.; Materazzi, A. Phylogenetic Analysis of Viruses in Tuscan Vitis vinifera sylvestris (Gmeli) Hegi. PLoS ONE 2018, 13, e0200875. [Google Scholar] [CrossRef] [PubMed]
- Hu, R.; Dias, N.P.; Soltani, N.; Vargas-Asencio, J.; Hensley, D.D.; Perry, K.L.; Domier, L.L.; Hajimorad, M.R. Cultivated and Wild Grapevines in Tennessee Possess Overlapping but Distinct Virus Populations. Plant Dis. 2021, 105, 2785–2791. [Google Scholar] [CrossRef] [PubMed]
- Nabeshima, T.; Abe, J. High-Throughput Sequencing Indicates Novel Varicosavirus, Emaravirus, and Deltapartitivirus Infections in Vitis coignetiae. Viruses 2021, 13, 827. [Google Scholar] [CrossRef] [PubMed]
- Petersen, S.M.; Keith, C.; Austin, K.; Howard, S.; Su, L.; Qui, W. A Natural Reservoir and Transmission Vector of Grapevine Vein Clearing Virus. Plant Dis. 2019, 103, 571–577. [Google Scholar] [CrossRef] [Green Version]
- Wege, C.; Saunders, K.; Stanley, J.; Jeske, H. Comparative Analysis of Tissue Tropism of Bipartite Geminiviruses. J. Phytopathol. 2001, 149, 359–368. [Google Scholar] [CrossRef]
- Heck, M.; Brault, V. Targeted Disruption of Aphid Transmission: A Vision for the Management of Crop Diseases Caused by Luteoviridae Members. Curr. Opin. Virol. 2018, 33, 24–32. [Google Scholar] [CrossRef] [Green Version]
- Preto, C.R.; Sudarshana, M.R.; Zalom, F.G. Feeding and Reproductive Hosts of Spissistilus festinus (Say) (Hemiptera: Membracidae) Found in Californian Vineyards. J. Econ. Entomol. 2018, 111, 2531–2535. [Google Scholar] [CrossRef]
- Moore, G.C.; Mueller, A.J. Biological Observations of the Threecornered Alfalfa Hopper on Soybean and Three Weed Species. J. Econ. Entomol. 1976, 69, 14–16. [Google Scholar] [CrossRef]
- Bick, E.N.; Kron, C.R.; Zalom, F.G. Timing the Implementation of Cultural Practices for Spissistilus festinus (Hemiptera: Membracidae) in California Vineyards Using a Stage-Structured Degree-Day Model. J. Econ. Entomol. 2020, 113, 2558–2562. [Google Scholar] [CrossRef]
- Bahder, B.W.; Zalom, F.G.; Jayanth, M.; Sudarshana, M.R. Phylogeny of the Geminivirus Coat Protein Sequence and Digital PCR Aid in Identifying Spissistilus festinus as a Vector of Grapevine red blotch-associated virus. Virology 2016, 106, 1223–1230. [Google Scholar] [CrossRef] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Leaves a | ||||||
|---|---|---|---|---|---|---|
| Free-Living Vine | Top A | Top B | Middle A | Middle B | Bottom A | Bottom B |
| Infected 1 | 2/3 | 2/3 | 2/3 | 2/3 | 2/3 | 1/3 |
| Infected 2 | 1/3 | 2/3 | 1/3 | 0/3 | 1/3 | 2/3 |
| Total b | 3/6 | 4/6 | 3/6 | 2/6 | 3/6 | 3/6 |
 |  |  | ||||
| Grand total c | 7/12 | 5/12 | 6/12 | |||
| Healthy | 0/3 | 0/3 | 0/3 | 0/3 | 0/3 | 0/3 |
| Virus Recipient Tissue | |||
|---|---|---|---|
| Virus Donor Vine | Cultivated a | Free-Living b | Total c |
| Cultivated | 5/27 c (19%) | 3/3 (100%) | 8/30 (27%) |
| Free-living | 2/13 (15%) | 10/13 (77%) | 12/26 (46%) |
| Total | 7/40 (18%) | 13/16 (81%) | 20/56 (36%) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hoyle, V.; Flasco, M.T.; Choi, J.; Cieniewicz, E.J.; McLane, H.; Perry, K.; Dangl, G.; Rwahnih, M.A.; Heck, M.; Loeb, G.; et al. Transmission of Grapevine Red Blotch Virus by Spissistilus festinus [Say, 1830] (Hemiptera: Membracidae) between Free-Living Vines and Vitis vinifera ‘Cabernet Franc’. Viruses 2022, 14, 1156. https://doi.org/10.3390/v14061156
Hoyle V, Flasco MT, Choi J, Cieniewicz EJ, McLane H, Perry K, Dangl G, Rwahnih MA, Heck M, Loeb G, et al. Transmission of Grapevine Red Blotch Virus by Spissistilus festinus [Say, 1830] (Hemiptera: Membracidae) between Free-Living Vines and Vitis vinifera ‘Cabernet Franc’. Viruses. 2022; 14(6):1156. https://doi.org/10.3390/v14061156
Chicago/Turabian StyleHoyle, Victoria, Madison T. Flasco, Jiyeong Choi, Elizabeth J. Cieniewicz, Heather McLane, Keith Perry, Gerald Dangl, Maher Al Rwahnih, Michelle Heck, Greg Loeb, and et al. 2022. "Transmission of Grapevine Red Blotch Virus by Spissistilus festinus [Say, 1830] (Hemiptera: Membracidae) between Free-Living Vines and Vitis vinifera ‘Cabernet Franc’" Viruses 14, no. 6: 1156. https://doi.org/10.3390/v14061156