
Can Virus-like Particles Be Used as Synergistic Agent in Pest Management?

 , and
, and
Abstract
:1. Introduction
2. Synergistic Agents as Innovative Strategies to Improve Insecticides Efficacy
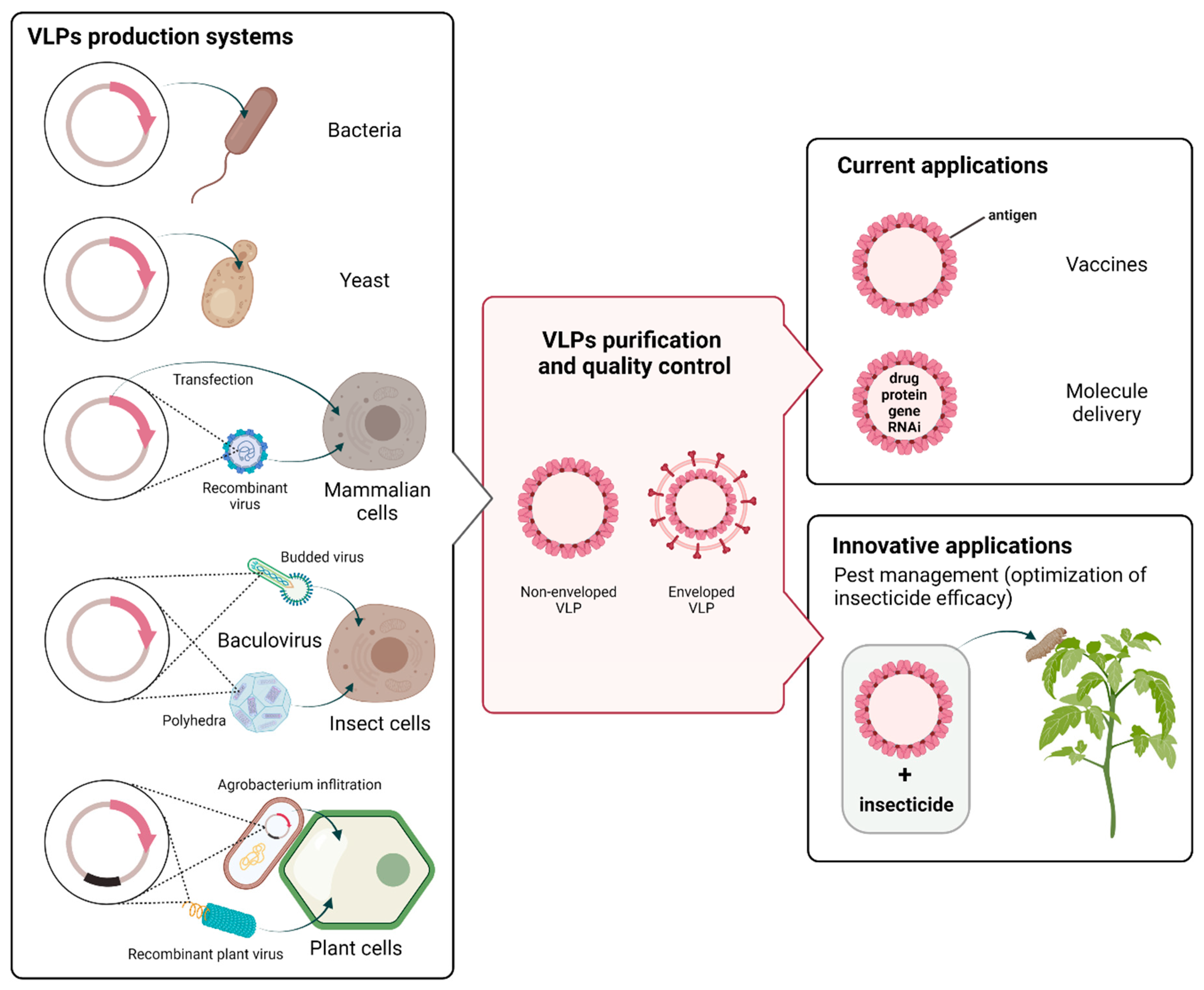
3. Various Systems Used to Produce VLPs
3.1. Bacterial Cells
3.2. Yeast Cells
3.3. Mammalian Cells
3.4. Baculovirus/Insect Cells
3.5. Plant Cells
4. Current Applications of VLPs
4.1. VLP-Based Vaccines
4.2. Molecule Delivery
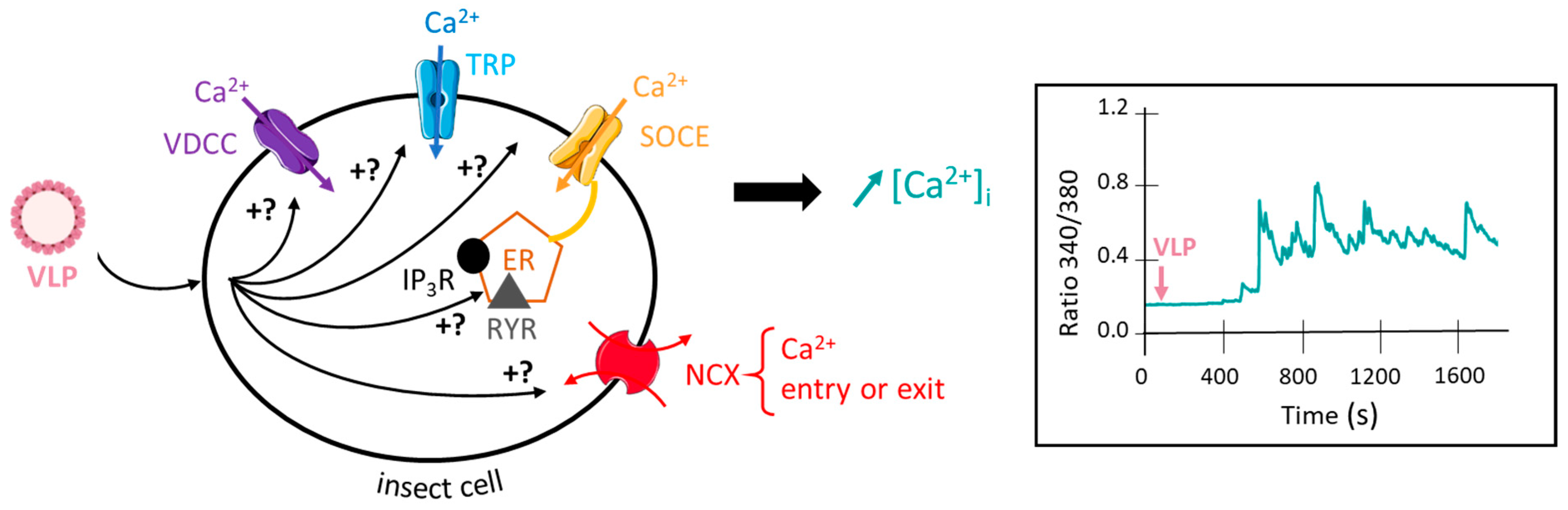
5. Future Direction in Pest Management: VLPs as Synergistic Agent of Insecticides
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- OECD. OECD Environmental Outlook to 2050: The Consequences of Inaction. Available online: https://www.oecd.org/g20/topics/energy-environment-green-growth/oecdenvironmentaloutlookto2050theconsequencesofinaction.htm (accessed on 1 December 2021).
- Mostafalou, S.; Abdollahi, M. Pesticides: An update of human exposure and toxicity. Arch. Toxicol. 2017, 91, 549–599. [Google Scholar] [CrossRef] [PubMed]
- Sanchez-Bayo, F. Indirect Effect of Pesticides on Insects and Other Arthropods. Toxics 2021, 9, 177. [Google Scholar] [CrossRef] [PubMed]
- Sparks, T.C.; Crossthwaite, A.J.; Nauen, R.; Banba, S.; Cordova, D.; Earley, F.; Ebbinghaus-Kintscher, U.; Fujioka, S.; Hirao, A.; Karmon, D.; et al. Insecticides, biologics and nematicides: Updates to IRAC’s mode of action classification—A tool for resistance management. Pestic. Biochem. Physiol. 2020, 167, 104587. [Google Scholar] [CrossRef] [PubMed]
- Balabanidou, V.; Grigoraki, L.; Vontas, J. Insect cuticle: A critical determinant of insecticide resistance. Curr. Opin. Insect Sci. 2018, 27, 68–74. [Google Scholar] [CrossRef] [PubMed]
- Casida, J.E.; Durkin, K.A. Neuroactive insecticides: Targets, selectivity, resistance, and secondary effects. Annu. Rev. Entomol. 2013, 58, 99–117. [Google Scholar] [CrossRef] [PubMed]
- Sparks, T.C.; Storer, N.; Porter, A.; Slater, R.; Nauen, R. Insecticide resistance management and industry: The origins and evolution of the Insecticide Resistance Action Committee (IRAC) and the mode of action classification scheme. Pest Manag. Sci. 2021, 77, 2609–2619. [Google Scholar] [CrossRef]
- United States Environmental Protection Agency. Available online: https://www.epa.gov/safepestcontrol/integrated-pest-management-ipm-principles (accessed on 3 November 2021).
- Deshayes, C.; Siegwart, M.; Pauron, D.; Froger, J.A.; Lapied, B.; Apaire-Marchais, V. Microbial Pest Control Agents: Are they a Specific and Safe Tool for Insect Pest Management? Curr. Med. Chem. 2017, 24, 2959–2973. [Google Scholar] [CrossRef]
- Murali-Baskaran, R.K.; Sharma, K.C.; Kaushal, P.; Kumar, J.; Parthiban, P.; Senthil-Nathan, S.; Mankin, R.W. Role of kairomone in biological control of crop pests—A review. Physiol. Mol. Plant Pathol. 2018, 101, 3–15. [Google Scholar] [CrossRef]
- Ons, L.; Bylemans, D.; Thevissen, K.; Cammue, B.P.A. Combining Biocontrol Agents with Chemical Fungicides for Integrated Plant Fungal Disease Control. Microorganisms 2020, 8, 1930. [Google Scholar] [CrossRef]
- Fletcher, S.J.; Reeves, P.T.; Hoang, B.T.; Mitter, N. A Perspective on RNAi-Based Biopesticides. Front. Plant Sci. 2020, 11, 51. [Google Scholar] [CrossRef] [Green Version]
- Kolliopoulou, A.; Taning, C.N.T.; Smagghe, G.; Swevers, L. Viral Delivery of dsRNA for Control of Insect Agricultural Pests and Vectors of Human Disease: Prospects and Challenges. Front. Physiol. 2017, 8, 399. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumar, A.; Choudhary, A.; Kaur, H.; Mehta, S.; Husen, A. Smart nanomaterial and nanocomposite with advanced agrochemical activities. Nanoscale Res. Lett. 2021, 16, 156. [Google Scholar] [CrossRef] [PubMed]
- Stejskal, V.; Vendl, T.; Aulicky, R.; Athanassiou, C. Synthetic and Natural Insecticides: Gas, Liquid, Gel and Solid Formulations for Stored-Product and Food-Industry Pest Control. Insects 2021, 12, 590. [Google Scholar] [CrossRef] [PubMed]
- Grillo, R.; Fraceto, L.F.; Amorim, M.J.B.; Scott-Fordsmand, J.J.; Schoonjans, R.; Chaudhry, Q. Ecotoxicological and regulatory aspects of environmental sustainability of nanopesticides. J. Hazard. Mater. 2021, 404, 124148. [Google Scholar] [CrossRef] [PubMed]
- Apaire-Marchais, V.; Ogliastro, M.; Chandre, F.; Pennetier, C.; Raymond, V.; Lapied, B. Virus and calcium: An unexpected tandem to optimize insecticide efficacy. Environ. Microbiol. Rep. 2016, 8, 168–178. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bonnet, J.; Pennetier, C.; Duchon, S.; Lapied, B.; Corbel, V. Multi-function oxidases are responsible for the synergistic interactions occurring between repellents and insecticides in mosquitoes. Parasites Vectors 2009, 2, 17. [Google Scholar] [CrossRef] [Green Version]
- Deshayes, C.; Moreau, E.; Pitti-Caballero, J.; Froger, J.A.; Apaire-Marchais, V.; Lapied, B. Synergistic agent and intracellular calcium, a successful partnership in the optimization of insecticide efficacy. Curr. Opin. Insect Sci. 2018, 30, 52–58. [Google Scholar] [CrossRef]
- El-Adouzi, M.; Apaire-Marchais, V.; Raymond, V.; Lapied, B. New Insecticide Composition. Patent 16,674,692, 4 June 2020. [Google Scholar]
- Moreau, E.; Mikulska-Ruminska, K.; Goulu, M.; Perrier, S.; Deshayes, C.; Stankiewicz, M.; Apaire-Marchais, V.; Nowak, W.; Lapied, B. Orthosteric muscarinic receptor activation by the insect repellent IR3535 opens new prospects in insecticide-based vector control. Sci. Rep. 2020, 10, 6842. [Google Scholar] [CrossRef] [Green Version]
- Pennetier, C.; Corbel, V.; Boko, P.; Odjo, A.; N’Guessan, R.; Lapied, B.; Hougard, J.M. Synergy between repellents and non-pyrethroid insecticides strongly extends the efficacy of treated nets against Anopheles Gambiae. Malar. J. 2007, 6, 38. [Google Scholar] [CrossRef] [Green Version]
- Pennetier, C.; Costantini, C.; Corbel, V.; Licciardi, S.; Dabire, R.K.; Lapied, B.; Chandre, F.; Hougard, J.M. Synergy between repellents and organophosphates on bed nets: Efficacy and behavioural response of natural free-flying An. gambiae mosquitoes. PLoS ONE 2009, 4, e7896. [Google Scholar] [CrossRef] [Green Version]
- Pitti Caballero, J.; Murillo, L.; List, O.; Bastiat, G.; Flochlay-Sigognault, A.; Guerino, F.; Lefrançois, C.; Lautram, N.; Lapied, B.; Apaire-Marchais, V. Nanoencapsulated deltamethrin as synergistic agent potentiates insecticide effect of indoxacarb through an unusual neuronal calcium-dependent mechanism. Pestic. Biochem. Physiol. 2019, 157, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Raymond, V.; Goven, D.; Benzidane, Y.; List, O.; Lapied, B. Influence of Cellular and Molecular Factors on Membrane Target Sensitivity to Insecticides. Curr. Med. Chem. 2017, 24, 2974–2987. [Google Scholar] [CrossRef] [PubMed]
- Snoeck, S.; Greenhalgh, R.; Tirry, L.; Clark, R.M.; Van Leeuwen, T.; Dermauw, W. The effect of insecticide synergist treatment on genome-wide gene expression in a polyphagous pest. Sci. Rep. 2017, 7, 13440. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bakowski, D.; Murray, F.; Parekh, A.B. Store-Operated Ca(2+) Channels: Mechanism, Function, Pharmacology, and Therapeutic Targets. Annu. Rev. Pharmacol. Toxicol. 2021, 61, 629–654. [Google Scholar] [CrossRef] [PubMed]
- Abd-Ella, A.; Stankiewicz, M.; Mikulska, K.; Nowak, W.; Pennetier, C.; Goulu, M.; Fruchart-Gaillard, C.; Licznar, P.; Apaire-Marchais, V.; List, O.; et al. The Repellent DEET Potentiates Carbamate Effects via Insect Muscarinic Receptor Interactions: An Alternative Strategy to Control Insect Vector-Borne Diseases. PLoS ONE 2015, 10, e0126406. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bodereau-Dubois, B.; List, O.; Calas-List, D.; Marques, O.; Communal, P.Y.; Thany, S.H.; Lapied, B. Transmembrane potential polarization, calcium influx, and receptor conformational state modulate the sensitivity of the imidacloprid-insensitive neuronal insect nicotinic acetylcholine receptor to neonicotinoid insecticides. J. Pharmacol. Exp. Ther. 2012, 341, 326–339. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Houchat, J.N.; Taillebois, E.; Thany, S.H. Effects of the DAG analogue 1,2-dioctanoyl-sn-glycerol (DiC8) on nicotine- and clothianidin-evoked currents through alpha-bungarotoxin-insensitive nicotinic acetylcholine receptors expressed on cockroach neurosecretory cells. Neurotoxicology 2020, 78, 143–151. [Google Scholar] [CrossRef]
- Lavialle-Defaix, C.; Moignot, B.; Legros, C.; Lapied, B. How does calcium-dependent intracellular regulation of voltage-dependent sodium current increase the sensitivity to the oxadiazine insecticide indoxacarb metabolite decarbomethoxylated JW062 (DCJW) in insect pacemaker neurons? J. Pharmacol. Exp. Ther. 2010, 333, 264–272. [Google Scholar] [CrossRef] [Green Version]
- List, O.; Calas-List, D.; Taillebois, E.; Juchaux, M.; Heuland, E.; Thany, S.H. Inhibition of PaCaMKII-E isoform in the dorsal unpaired median neurosecretory cells of cockroach reduces nicotine- and clothianidin-induced currents. J. Neurochem. 2014, 130, 507–513. [Google Scholar] [CrossRef]
- Murillo, L.; Hamon, A.; Es-Salah-Lamoureux, Z.; Itier, V.; Quinchard, S.; Lapied, B. Inhibition of protein kinase C decreases sensitivity of GABA receptor subtype to fipronil insecticide in insect neurosecretory cells. Neurotoxicology 2011, 32, 828–835. [Google Scholar] [CrossRef]
- Pennetier, C.; Costantini, C.; Corbel, V.; Licciardi, S.; Dabire, R.K.; Lapied, B.; Chandre, F.; Hougard, J.M. Mixture for controlling insecticide-resistant malaria vectors. Emerg. Infect. Dis. 2008, 14, 1707–1714. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lapied, B.; Pennetier, C.; Apaire-Marchais, V.; Licznar, P.; Corbel, V. Innovative applications for insect viruses: Towards insecticide sensitization. Trends Biotechnol. 2009, 27, 190–198. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Licznar, P.; List, O.; Goven, D.; Nna, R.N.; Lapied, B.; Apaire-Marchais, V. A novel method using Autographa californica multiple nucleopolyhedrovirus for increasing the sensitivity of insecticide through calcium influx in insect cell line. J. Virol. Methods 2014, 195, 72–75. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chung, Y.H.; Cai, H.; Steinmetz, N.F. Viral nanoparticles for drug delivery, imaging, immunotherapy, and theranostic applications. Adv. Drug Deliv. Rev. 2020, 156, 214–235. [Google Scholar] [CrossRef] [PubMed]
- Steinmetz, N.F. Viral nanoparticles as platforms for next-generation therapeutics and imaging devices. Nanomed. Nanotechnol. Biol. Med. 2010, 6, 634–641. [Google Scholar] [CrossRef] [Green Version]
- Nooraei, S.; Bahrulolum, H.; Hoseini, Z.S.; Katalani, C.; Hajizade, A.; Easton, A.J.; Ahmadian, G. Virus-like particles: Preparation, immunogenicity and their roles as nanovaccines and drug nanocarriers. J. Nanobiotechnology 2021, 19, 59. [Google Scholar] [CrossRef]
- Huang, X.; Wang, X.; Zhang, J.; Xia, N.; Zhao, Q. Escherichia coli-derived virus-like particles in vaccine development. Npj Vaccines 2017, 2, 3. [Google Scholar] [CrossRef] [Green Version]
- Kim, H.J.; Kim, H.J. Yeast as an expression system for producing virus-like particles: What factors do we need to consider? Lett. Appl. Microbiol. 2017, 64, 111–123. [Google Scholar] [CrossRef]
- Kost, T.A.; Condreay, J.P.; Jarvis, D.L. Baculovirus as versatile vectors for protein expression in insect and mammalian cells. Nat. Biotechnol. 2005, 23, 567–575. [Google Scholar] [CrossRef]
- Marsian, J.; Lomonossoff, G.P. Molecular pharming—VLPs made in plants. Curr. Opin. Biotechnol. 2016, 37, 201–206. [Google Scholar] [CrossRef] [Green Version]
- Glass, P.J.; White, L.J.; Ball, J.M.; Leparc-Goffart, I.; Hardy, M.E.; Estes, M.K. Norwalk virus open reading frame 3 encodes a minor structural protein. J. Virol. 2000, 74, 6581–6591. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fuenmayor, J.; Gòdia, F.; Cervera, L. Production of virus-like particles for vaccines. New Biotechnol. 2017, 39, 174–180. [Google Scholar] [CrossRef] [PubMed]
- Le, D.T.; Muller, K.M. In Vitro Assembly of Virus-Like Particles and Their Applications. Life 2021, 11, 334. [Google Scholar] [CrossRef] [PubMed]
- Zeltins, A. Construction and characterization of virus-like particles: A review. Mol. Biotechnol. 2013, 53, 92–107. [Google Scholar] [CrossRef]
- Roldao, A.; Mellado, M.C.; Castilho, L.R.; Carrondo, M.J.; Alves, P.M. Virus-like particles in vaccine development. Expert Rev. Vaccines 2010, 9, 1149–1176. [Google Scholar] [CrossRef]
- Roldão, A.; Silva, A.C.; Mellado, M.C.M.; Alves, P.M.; Carrondo, M.J.T. Viruses and Virus-Like Particles in Biotechnology: Fundamentals and Applications. Compr. Biotechnol. 2017, 1, 633–656. [Google Scholar] [CrossRef]
- Grataitong, K.; Huault, S.; Chotwiwatthanakun, C.; Jariyapong, P.; Thongsum, O.; Chawiwithaya, C.; Chakrabandhu, K.; Hueber, A.-O.; Weerachatyanukul, W. Chimeric virus-like particles (VLPs) designed from shrimp nodavirus (MrNV) capsid protein specifically target EGFR-positive human colorectal cancer cells. Sci. Rep. 2021, 11, 16579. [Google Scholar] [CrossRef]
- Lai, W.B.; Middelberg, A.P. The production of human papillomavirus type 16 L1 vaccine product from Escherichia coli inclusion bodies. Bioprocess Biosyst. Eng. 2002, 25, 121–128. [Google Scholar] [CrossRef]
- Selas Castiñeiras, T.; Williams, S.G.; Hitchcock, A.G.; Smith, D.C. E. coli strain engineering for the production of advanced biopharmaceutical products. FEMS Microbiol. Lett. 2018, 365, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Zhang, W.; Carmichael, J.; Ferguson, J.; Inglis, S.; Ashrafian, H.; Stanley, M. Expression of human papillomavirus type 16 L1 protein in Escherichia coli: Denaturation, renaturation, and self-assembly of virus-like particles in vitro. Virology 1998, 243, 423–431. [Google Scholar] [CrossRef] [Green Version]
- Dai, S.; Wang, H.; Deng, F. Advances and challenges in enveloped virus-like particle (VLP)-based vaccines. J. Immunol. Sci. 2018, 2, 36–41. [Google Scholar]
- Liu, W.; Jiang, H.; Zhou, J.; Yang, X.; Tang, Y.; Fang, D.; Jiang, L. Recombinant dengue virus-like particles from Pichia pastoris: Efficient production and immunological properties. Virus Genes 2010, 40, 53–59. [Google Scholar] [CrossRef] [PubMed]
- Sakuragi, S.; Goto, T.; Sano, K.; Morikawa, Y. HIV type 1 Gag virus-like particle budding from spheroplasts of Saccharomyces cerevisiae. Proc. Natl. Acad. Sci. USA 2002, 99, 7956–7961. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Berger, I.; Fitzgerald, D.J.; Richmond, T.J. Baculovirus expression system for heterologous multiprotein complexes. Nat. Biotechnol. 2004, 22, 1583–1587. [Google Scholar] [CrossRef] [PubMed]
- Harrison, R.L.; Jarvis, D.L. Transforming lepidopteran insect cells for improved protein processing. Methods Mol. Biol. 2007, 388, 341–356. [Google Scholar] [CrossRef]
- Jarvis, D.L.; Kawar, Z.S.; Hollister, J.R. Engineering N-glycosylation pathways in the baculovirus-insect cell system. Curr. Opin. Biotechnol. 1998, 9, 528–533. [Google Scholar] [CrossRef]
- Legastelois, I.; Buffin, S.; Peubez, I.; Mignon, C.; Sodoyer, R.; Werle, B. Non-conventional expression systems for the production of vaccine proteins and immunotherapeutic molecules. Hum. Vaccines Immunother. 2017, 13, 947–961. [Google Scholar] [CrossRef] [Green Version]
- Liu, F.; Wu, X.; Li, L.; Liu, Z.; Wang, Z. Use of baculovirus expression system for generation of virus-like particles: Successes and challenges. Protein Expr. Purif. 2013, 90, 104–116. [Google Scholar] [CrossRef]
- Walski, T.; De Schutter, K.; Van Damme, E.J.M.; Smagghe, G. Diversity and functions of protein glycosylation in insects. Insect Biochem. Mol. Biol. 2017, 83, 21–34. [Google Scholar] [CrossRef]
- Wang, P.; Granados, R.R.; Shuler, M.L. Studies on serum-free culture of insect cells for virus propagation and recombinant protein production. J. Invertebr. Pathol. 1992, 59, 46–53. [Google Scholar] [CrossRef]
- Yamaji, H.; Manabe, T.; Kitaura, A.; Izumoto, E.; Fukuda, H. Efficient production of recombinant protein in immobilized insect cell culture using serum-free basal media after baculovirus infection. Biochem. Eng. J. 2006, 28, 67–72. [Google Scholar] [CrossRef] [Green Version]
- Zhu, F.; Li, D.; Chen, K. Structures and functions of invertebrate glycosylation. Open Biol. 2019, 9, 180232. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cervera, L.; Gutierrez-Granados, S.; Martinez, M.; Blanco, J.; Godia, F.; Segura, M.M. Generation of HIV-1 Gag VLPs by transient transfection of HEK 293 suspension cell cultures using an optimized animal-derived component free medium. J. Biotechnol. 2013, 166, 152–165. [Google Scholar] [CrossRef]
- Fontana, D.; Kratje, R.; Etcheverrigaray, M.; Prieto, C. Immunogenic virus-like particles continuously expressed in mammalian cells as a veterinary rabies vaccine candidate. Vaccine 2015, 33, 4238–4246. [Google Scholar] [CrossRef] [PubMed]
- Ren, J.; Bell, G.; Coy, D.H.; Brunicardi, F.C. Activation of human somatostatin receptor type 2 causes inhibition of cell growth in transfected HEK293 but not in transfected CHO cells. J. Surg. Res. 1997, 71, 13–18. [Google Scholar] [CrossRef]
- Cid, R.; Bolívar, J. Platforms for Production of Protein-Based Vaccines: From Classical to Next-Generation Strategies. Biomolecules 2021, 11, 1072. [Google Scholar] [CrossRef]
- Huang, Z.; Elkin, G.; Maloney, B.J.; Beuhner, N.; Arntzen, C.J.; Thanavala, Y.; Mason, H.S. Virus-like particle expression and assembly in plants: Hepatitis B and Norwalk viruses. Vaccine 2005, 23, 1851–1858. [Google Scholar] [CrossRef]
- Komarova, T.V.; Baschieri, S.; Donini, M.; Marusic, C.; Benvenuto, E.; Dorokhov, Y.L. Transient expression systems for plant-derived biopharmaceuticals. Expert Rev. Vaccines 2010, 9, 859–876. [Google Scholar] [CrossRef]
- Lossl, A.G.; Waheed, M.T. Chloroplast-derived vaccines against human diseases: Achievements, challenges and scopes. Plant Biotechnol. J. 2011, 9, 527–539. [Google Scholar] [CrossRef]
- Mason, H.S.; Warzecha, H.; Mor, T.; Arntzen, C.J. Edible plant vaccines: Applications for prophylactic and therapeutic molecular medicine. Trends Mol. Med. 2002, 8, 324–329. [Google Scholar] [CrossRef]
- Oey, M.; Lohse, M.; Kreikemeyer, B.; Bock, R. Exhaustion of the chloroplast protein synthesis capacity by massive expression of a highly stable protein antibiotic. Plant J. Cell Mol. Biol. 2009, 57, 436–445. [Google Scholar] [CrossRef] [PubMed]
- Scotti, N.; Rybicki, E.P. Virus-like particles produced in plants as potential vaccines. Expert Rev. Vaccines 2013, 12, 211–224. [Google Scholar] [CrossRef] [PubMed]
- Xu, J.; Towler, M.; Weathers, P.J. Platforms for Plant-Based Protein Production. Bioprocess. Plant Vitr. Syst. 2018, 509–548. [Google Scholar] [CrossRef]
- Aires, K.A.; Cianciarullo, A.M.; Carneiro, S.M.; Villa, L.L.; Boccardo, E.; Perez-Martinez, G.; Perez-Arellano, I.; Oliveira, M.L.; Ho, P.L. Production of human papillomavirus type 16 L1 virus-like particles by recombinant Lactobacillus casei cells. Appl. Environ. Microbiol. 2006, 72, 745–752. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Phelps, J.P.; Dao, P.; Jin, H.; Rasochova, L. Expression and self-assembly of cowpea chlorotic mottle virus-like particles in Pseudomonas fluorescens. J. Biotechnol. 2007, 128, 290–296. [Google Scholar] [CrossRef]
- Lipke, P.N.; Ovalle, R. Cell wall architecture in yeast: New structure and new challenges. J. Bacteriol. 1998, 180, 3735–3740. [Google Scholar] [CrossRef] [Green Version]
- Saraswat, S.; Athmaram, T.N.; Parida, M.; Agarwal, A.; Saha, A.; Dash, P.K. Expression and Characterization of Yeast Derived Chikungunya Virus Like Particles (CHIK-VLPs) and Its Evaluation as a Potential Vaccine Candidate. PLOS Negl. Trop. Dis. 2016, 10, e0004782. [Google Scholar] [CrossRef] [Green Version]
- Qian, C.; Liu, X.; Xu, Q.; Wang, Z.; Chen, J.; Li, T.; Zheng, Q.; Yu, H.; Gu, Y.; Li, S.; et al. Recent Progress on the Versatility of Virus-Like Particles. Vaccines 2020, 8, 139. [Google Scholar] [CrossRef] [Green Version]
- Shirbaghaee, Z.; Bolhassani, A. Different applications of virus-like particles in biology and medicine: Vaccination and delivery systems. Biopolymers 2016, 105, 113–132. [Google Scholar] [CrossRef]
- Mazalovska, M.; Kouokam, J.C. Progress in the Production of Virus-Like Particles for Vaccination against Hepatitis E Virus. Viruses 2020, 12, 826. [Google Scholar] [CrossRef]
- Tagliamonte, M.; Visciano, M.L.; Tornesello, M.L.; De Stradis, A.; Buonaguro, F.M.; Buonaguro, L. HIV-Gag VLPs presenting trimeric HIV-1 gp140 spikes constitutively expressed in stable double transfected insect cell line. Vaccine 2011, 29, 4913–4922. [Google Scholar] [CrossRef] [PubMed]
- BioSpace. Medicago Develops Plant-Based Coronavirus Vaccine Candidate. Available online: https://www.biospace.com/article/medicago-successfully-produces-a-viable-vaccine-candidate-for-covid-19/ (accessed on 4 October 2021).
- Mi, Y.; Xie, T.; Zhu, B.; Tan, J.; Li, X.; Luo, Y.; Li, F.; Niu, H.; Han, J.; Lv, W.; et al. Production of SARS-CoV-2 Virus-Like Particles in Insect Cells. Vaccines 2021, 9, 554. [Google Scholar] [CrossRef] [PubMed]
- Opriessnig, T.; Mattei, A.A.; Karuppannan, A.K.; Halbur, P.G. Future perspectives on swine viral vaccines: Where are we headed? Porc. Health Manag. 2021, 7, 1. [Google Scholar] [CrossRef] [PubMed]
- Zochowska, M.; Paca, A.; Schoehn, G.; Andrieu, J.P.; Chroboczek, J.; Dublet, B.; Szolajska, E. Adenovirus dodecahedron, as a drug delivery vector. PLoS ONE 2009, 4, e5569. [Google Scholar] [CrossRef] [PubMed]
- Zochowska, M.; Piguet, A.C.; Jemielity, J.; Kowalska, J.; Szolajska, E.; Dufour, J.F.; Chroboczek, J. Virus-like particle-mediated intracellular delivery of mRNA cap analog with in vivo activity against hepatocellular carcinoma. Nanomed. Nanotechnol. Biol. Med. 2015, 11, 67–76. [Google Scholar] [CrossRef]
- Pan, Y.; Jia, T.; Zhang, Y.; Zhang, K.; Zhang, R.; Li, J.; Wang, L. MS2 VLP-based delivery of microRNA-146a inhibits autoantibody production in lupus-prone mice. Int. J. Nanomed. 2012, 7, 5957–5967. [Google Scholar] [CrossRef] [Green Version]
- Ashley, C.E.; Carnes, E.C.; Phillips, G.K.; Durfee, P.N.; Buley, M.D.; Lino, C.A.; Padilla, D.P.; Phillips, B.; Carter, M.B.; Willman, C.L.; et al. Cell-specific delivery of diverse cargos by bacteriophage MS2 virus-like particles. ACS Nano 2011, 5, 5729–5745. [Google Scholar] [CrossRef] [Green Version]
- Li, C.; Liu, F.; Liang, M.; Zhang, Q.; Wang, X.; Wang, T.; Li, J.; Li, D. Hantavirus-like particles generated in CHO cells induce specific immune responses in C57BL/6 mice. Vaccine 2010, 28, 4294–4300. [Google Scholar] [CrossRef]
- Purdy, D.E.; Chang, G.J. Secretion of noninfectious dengue virus-like particles and identification of amino acids in the stem region involved in intracellular retention of envelope protein. Virology 2005, 333, 239–250. [Google Scholar] [CrossRef] [Green Version]
- Thompson, C.M.; Petiot, E.; Lennaertz, A.; Henry, O.; Kamen, A.A. Analytical technologies for influenza virus-like particle candidate vaccines: Challenges and emerging approaches. Virol. J. 2013, 10, 141. [Google Scholar] [CrossRef] [Green Version]
- Xu, R.; Shi, M.; Li, J.; Song, P.; Li, N. Construction of SARS-CoV-2 Virus-Like Particles by Mammalian Expression System. Front. Bioeng. Biotechnol. 2020, 8, 862. [Google Scholar] [CrossRef] [PubMed]
- Marek, M.; van Oers, M.M.; Devaraj, F.F.; Vlak, J.M.; Merten, O.W. Engineering of baculovirus vectors for the manufacture of virion-free biopharmaceuticals. Biotechnol. Bioeng. 2011, 108, 1056–1067. [Google Scholar] [CrossRef] [PubMed]
- Yang, L.; Song, Y.; Li, X.; Huang, X.; Liu, J.; Ding, H.; Zhu, P.; Zhou, P. HIV-1 virus-like particles produced by stably transfected Drosophila S2 cells: A desirable vaccine component. J. Virol. 2012, 86, 7662–7676. [Google Scholar] [CrossRef] [Green Version]
- Gleba, Y.; Klimyuk, V.; Marillonnet, S. Magnifection—A new platform for expressing recombinant vaccines in plants. Vaccine 2005, 23, 2042–2048. [Google Scholar] [CrossRef] [PubMed]
- Huang, Z.; Chen, Q.; Hjelm, B.; Arntzen, C.; Mason, H. A DNA replicon system for rapid high-level production of virus-like particles in plants. Biotechnol. Bioeng. 2009, 103, 706–714. [Google Scholar] [CrossRef] [Green Version]
- Santi, L.; Batchelor, L.; Huang, Z.; Hjelm, B.; Kilbourne, J.; Arntzen, C.J.; Chen, Q.; Mason, H.S. An efficient plant viral expression system generating orally immunogenic Norwalk virus-like particles. Vaccine 2008, 26, 1846–1854. [Google Scholar] [CrossRef] [Green Version]
- Balke, I.; Zeltins, A. Recent Advances in the Use of Plant Virus-Like Particles as Vaccines. Viruses 2020, 12, 270. [Google Scholar] [CrossRef] [Green Version]
- Rybicki, E.P. Plant molecular farming of virus-like nanoparticles as vaccines and reagents. WIREs Nanomed. Nanobiotechnology 2020, 12, e1587. [Google Scholar] [CrossRef]
- Santoni, M.; Zampieri, R.; Avesani, L. Plant Virus Nanoparticles for Vaccine Applications. Curr. Protein Pept. Sci. 2020, 21, 344–356. [Google Scholar] [CrossRef]
- Hill, B.D.; Zak, A.; Khera, E.; Wen, F. Engineering Virus-like Particles for Antigen and Drug Delivery. Curr. Protein Pept. Sci. 2018, 19, 112–127. [Google Scholar] [CrossRef]
- Jeong, H.; Seong, B.L. Exploiting virus-like particles as innovative vaccines against emerging viral infections. J. Microbiol. 2017, 55, 220–230. [Google Scholar] [CrossRef] [PubMed]
- Zhang, N.; Li, C.; Jiang, S.; Du, L. Recent Advances in the Development of Virus-Like Particle-Based Flavivirus Vaccines. Vaccines 2020, 8, 481. [Google Scholar] [CrossRef] [PubMed]
- Collins, K.A.; Snaith, R.; Cottingham, M.G.; Gilbert, S.C.; Hill, A.V.S. Enhancing protective immunity to malaria with a highly immunogenic virus-like particle vaccine. Sci. Rep. 2017, 7, 46621. [Google Scholar] [CrossRef] [Green Version]
- Balke, I.; Zeltins, A. Use of plant viruses and virus-like particles for the creation of novel vaccines. Adv. Drug Deliv. Rev. 2019, 145, 119–129. [Google Scholar] [CrossRef]
- Kong, Q.; Richter, L.; Yang, Y.F.; Arntzen, C.J.; Mason, H.S.; Thanavala, Y. Oral immunization with hepatitis B surface antigen expressed in transgenic plants. Proc. Natl. Acad. Sci. USA 2001, 98, 11539–11544. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mason, H.S.; Ball, J.M.; Shi, J.J.; Jiang, X.; Estes, M.K.; Arntzen, C.J. Expression of Norwalk virus capsid protein in transgenic tobacco and potato and its oral immunogenicity in mice. Proc. Natl. Acad. Sci. USA 1996, 93, 5335–5340. [Google Scholar] [CrossRef] [Green Version]
- Medicago. Medicago Announces Phase 3 Study of VLP Quadrivalent Influenza Vaccine. Available online: https://medicago.com/en/press-release/medicago-announces-phase-3-study-of-vlp-quadrivalent-influenza-vaccine/ (accessed on 6 October 2021).
- Patra, J.K.; Das, G.; Fraceto, L.F.; Campos, E.V.R.; Rodriguez-Torres, M.D.P.; Acosta-Torres, L.S.; Diaz-Torres, L.A.; Grillo, R.; Swamy, M.K.; Sharma, S.; et al. Nano based drug delivery systems: Recent developments and future prospects. J. Nanobiotechnol. 2018, 16, 71. [Google Scholar] [CrossRef] [Green Version]
- Comas-Garcia, M.; Colunga-Saucedo, M.; Rosales-Mendoza, S. The Role of Virus-Like Particles in Medical Biotechnology. Mol. Pharm. 2020, 17, 4407–4420. [Google Scholar] [CrossRef]
- Rohovie, M.J.; Nagasawa, M.; Swartz, J.R. Virus-like particles: Next-generation nanoparticles for targeted therapeutic delivery. Bioeng. Transl. Med. 2017, 2, 43–57. [Google Scholar] [CrossRef]
- Zdanowicz, M.; Chroboczek, J. Virus-like particles as drug delivery vectors. Acta Biochim. Pol. 2016, 63, 469–473. [Google Scholar] [CrossRef]
- Shan, W.; Zhang, D.; Wu, Y.; Lv, X.; Hu, B.; Zhou, X.; Ye, S.; Bi, S.; Ren, L.; Zhang, X. Modularized peptides modified HBc virus-like particles for encapsulation and tumor-targeted delivery of doxorubicin. Nanomed. Nanotechnol. Biol. Med. 2018, 14, 725–734. [Google Scholar] [CrossRef] [PubMed]
- Wei, B.; Wei, Y.; Zhang, K.; Wang, J.; Xu, R.; Zhan, S.; Lin, G.; Wang, W.; Liu, M.; Wang, L.; et al. Development of an antisense RNA delivery system using conjugates of the MS2 bacteriophage capsids and HIV-1 TAT cell-penetrating peptide. Biomed. Pharmacother. Biomed. Pharmacother. 2009, 63, 313–318. [Google Scholar] [CrossRef] [PubMed]
- Somiya, M.; Qiushi, L.; Kuroda, S.I. Current Progress of Virus-mimicking Nanocarriers for Drug Delivery. Nanotheranostics 2017, 1, 415–429. [Google Scholar] [CrossRef] [PubMed]
- World Health Organization. Hepatitis B vaccines: WHO position paper—July 2017—Vaccins anti-hépatite B: Note de synthèse de l’OMS—juillet 2017. Wkly. Epidemiol. Rec. Relev. Épidémiologique Hebd. 2017, 92, 369–392. [Google Scholar]
- CDC. Hepatitis B Vaccines, Safety Information. Available online: https://www.cdc.gov/vaccinesafety/vaccines/hepatitis-b-vaccine.html (accessed on 12 April 2022).
- World Health Organization. Hepatitis B vaccines: WHO position paper--recommendations. Vaccine 2010, 28, 589–590. [Google Scholar] [CrossRef]
- Heine, M.; Heck, J.; Ciuraszkiewicz, A.; Bikbaev, A. Dynamic compartmentalization of calcium channel signalling in neurons. Neuropharmacology 2020, 169, 107556. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Production Platforms | Advantages | Disadvantages | Yield Range | Type of VLP Produced | References |
|---|---|---|---|---|---|
| E. coli | Ease of expression Fast growth rate High-level expression Simple process scale-up Low production cost | No PTMs Limited protein solubility Misfolded proteins Endotoxin contamination Production of simple VLPs | 0.75 to 700 μg/mL | Non-enveloped | [45,51,52,53] |
| Yeast | Capacity of simple PTMs Ease of expression High-level expression Ability to scale-up Low production cost | Limited PTMs Risk of incorrect folding and assembly Production of simple VLPs (cell wall) | 0.75 to 700 μg/mL | Non-enveloped; Enveloped (if using yeast spheroplasts); Unique and multiple structural proteins | [41,45,54,55,56] |
| Insect cells | Capacity of most eukaryotic-type PTMs Cell culture in suspension, without CO2 Free of mammalian pathogens Production of complex VLPs | Simpler N-glycosylation compared to mammalian cells Low yield High production cost Difficult to scale-up Baculovirus contamination | 0.2 and 18 μg/mL | Non-enveloped; Enveloped; Unique and multiple structural proteins | [42,45,48,54,57,58,59,60,61,62,63,64,65] |
| Mammalian cells | Complex PTMs Assembly and folding Possible cell culture in suspension Production of complex VLPs | Low cell growth rate Long expression time Low yield High production cost Difficult to scale-up Risk of contamination by mammalian pathogens | 0.018 and 10 μg/mL | Non-enveloped; Enveloped; Unique and multiple structural proteins | [45,48,66,67,68] |
| Plants | Complex PTMs (nucleus) Ease of expression High expression levels of up to 80% total soluble protein Ability to scale-up Low production cost VLPs storage (protected in plants) Potential oral immunization by simply ingesting VLPs in edible plant parts | No PTMs (chloroplasts) Time-consuming production of stable transgenic plants Low-level expression Low VLP assembly and stability Production of simple VLPs Technical issues (transgenic plants) | 4 to 2380 pg/mg of leaf | Non-enveloped; Unique and multiple structural proteins | [45,60,69,70,71,72,73,74,75,76] |
| Pathology | VLPs Composition | VLP Type | Expression System | Status | References | |
|---|---|---|---|---|---|---|
| VACCINES | Hepatitis B virus (HBV) infection | HBsAg | NE | Yeast (S. cerevisiae) | Licensed (Engenerix-B® and Recombivax HB®) | [45,81,82] |
| S, pre-S1, and pre-S2 | Mammalian cells (CHO) | Licensed (Sci-B-Vac®) | ||||
| Human papillomavirus (HPV) infection | HPV 6/11/16/18 L1 | NE | Yeast (S. cerevisiae) | Licensed (Gardasil®) | [45,69,81,82] | |
| HPV 6/11/16/18/31/33/45 /52/58 L1 | Yeast (S. cerevisiae) | Licensed (Gardasil 9®) | ||||
| HPV 16/18 L1 | Baculovirus/Insect cells (High-Five™) | Licensed (Cervarix®) | ||||
| Hepatitis E virus (HEV) infection | p239 | NE | Bacteria (E. coli) | Licensed (China) (Hecolin®) | [81,82,83] | |
| peptide | Baculovirus/Insect cells (Sf9) | Clinical trial phase | ||||
| Malaria | CSP into the HBsAg | NE | Yeast (S. cerevisiae) | Licensed (Mosquirix®) | [45,81] | |
| Human immunodeficiency virus (HIV) infection | p17 and p24 | E | Yeast (S. cerevisiae) | Clinical trial phase | [45,84] | |
| Gag or Env | Mammalian cells (HEK293) | |||||
| Gag or Env | Baculovirus/Insect cells (High-Five™) | |||||
| Human parvovirus B19 infection | VP1 and VP2 | NE | Baculovirus/Insect cells (Sf9) | Clinical trial phase | [45,82] | |
| Influenzavirus A infection | HA quadrivalent | E | Baculovirus/Insect cell (Sf9) | Licensed (Supemtek®) | [43,45,69,85] | |
| Plant (Nicotiana benthamania) | Clinical trial phase | |||||
| SARS-CoV infection | SP, EP, MP | NE | Baculovirus/Insect cells (Sf9) | Clinical trial phase | [37,85,86] | |
| Plant (Nicotiana benthamania) | Clinical trial phase | |||||
| Porcine circovirus type 2 infection | ORF2 | NE | Baculovirus/Insect cells (Sf9) | Licensed (Circumvent®) | [69,87] | |
| MOLECULE DELIVERY | Cancers | Bleomycin cross-linked at the surface of Dd-Ad3 VLPs | NE | Baculovirus/Insect cells (High-Five™) | In vitro research | [88] |
| Hepatocellular carcinoma (HCC) | Cap structure analog or Doxorubicin cross-linked at the surface of Dd-Ad3 VLPs | NE | Baculovirus/Insect cells (High-Five™) | Preclinical research | [89] | |
| Systemic lupus erythematosus | miRNA-146a packaged into conjugated MS2 bacteriophage capsid coated with HIV Tat47-57 peptide | NE | Bacteria (E. coli) | Preclinical research | [90] | |
| Hepatocellular carcinoma (HCC) | Doxorubicin, cisplatin, 5-fluorouracil, or SiRNA packaged into MS2 bacteriophage capsid coated with SP94 targeting or histidine-rich fusogenic peptides | NE | Bacteria (E. coli) | In vitro research | [91] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Deshayes, C.; Gosselin-Grenet, A.-S.; Ogliastro, M.; Lapied, B.; Apaire-Marchais, V. Can Virus-like Particles Be Used as Synergistic Agent in Pest Management? Viruses 2022, 14, 943. https://doi.org/10.3390/v14050943
Deshayes C, Gosselin-Grenet A-S, Ogliastro M, Lapied B, Apaire-Marchais V. Can Virus-like Particles Be Used as Synergistic Agent in Pest Management? Viruses. 2022; 14(5):943. https://doi.org/10.3390/v14050943
Chicago/Turabian StyleDeshayes, Caroline, Anne-Sophie Gosselin-Grenet, Mylène Ogliastro, Bruno Lapied, and Véronique Apaire-Marchais. 2022. "Can Virus-like Particles Be Used as Synergistic Agent in Pest Management?" Viruses 14, no. 5: 943. https://doi.org/10.3390/v14050943