
A Short Series of Case Reports of COVID-19 in Immunocompromised Patients

 , , , ,
, , , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Samples, Viral RNA Extraction, and Real-Time PCR
2.2. SARS-CoV-2 Genomic Sequencing
2.3. ELISA and End-Point Absolute Viral Neutralization Assay
2.4. HCoV Peptide Array Experiment and Data Analysis
3. Results
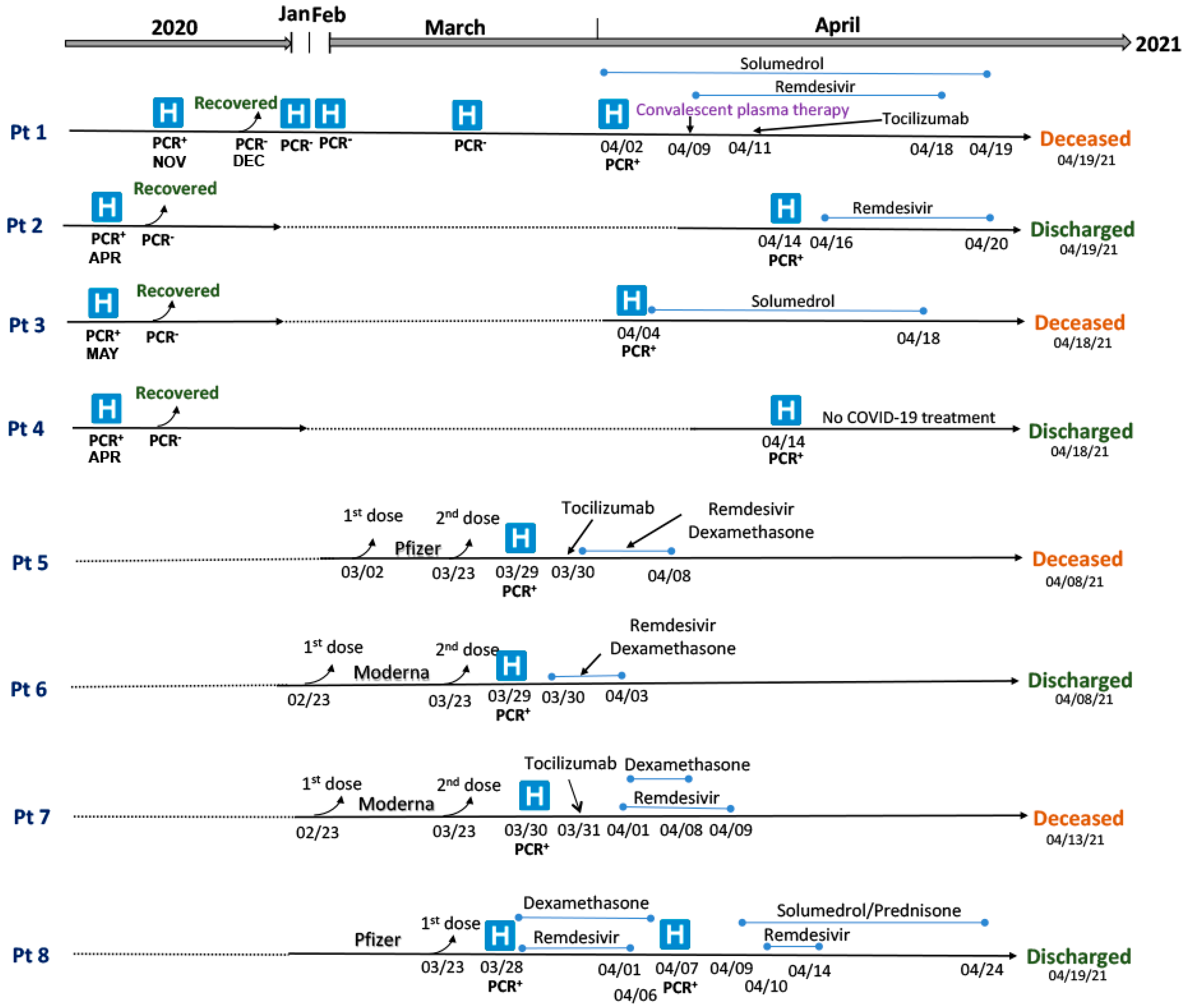
3.1. Clinical Summary of Patients with SARS-CoV-2 Infections and Re-Infections

{kind=link}
{kind=link}
{kind=link}
| Patient IDs | Sex (~Age Range in Years) | Latest Hospitalization Date * | COVID-19 Symptoms | COVID-19 Outcome (Days Since Hospitalization) | Immunocompromised Conditions | Immunosuppressed Treatment | Other Co-Morbidities |
|---|---|---|---|---|---|---|---|
| Pt 1 | M (70–74) | 2 April 2021 | Hemoptysis, persistent high fevers, diarrhea, cough | Died (17 d) due to cardiac arrest as a complication of COVID-19 infection | Diffuse large B-cell lymphoma (DLBCL) | Rituximab | Hypertension, non-ST-elevation myocardial infarction, coronary artery disease (CAD), coronary artery bypass graft (CABG) |
| Pt 2 | F (50–54) | 14 April 2021 | Fever, nausea, chills, severe headache | Recovered (5 d) | Renal transplant (02/2021) and Neutropenia | Tacrolimus, Mycophenolate, plasmapheresis, IVIG | Hypothyroidism, DM2, hypertension |
| Pt 3 | F (60–64) | 4 April 2021 | Shortness of breath, malaise | Died (14 d) due to multiorgan failure (cardiopulmonary, hepatic, renal), acute cerebrovascular accident (CVA) | Chronic obstructive pulmonary disease (COPD) | NA | Opioid use, alcohol use, hypothyroidism |
| Pt 4 | F (30–34) | 14 April 2021 | Fatigue | Recovered (4 d) | Systemic lupus erythematous (SLE) and lupus nephritis | High-dose Prednisone | Sickle cell trait, hypertension |
| Pt 5 | M (55–59) | 29 March 2021 | Shortness of breath, dyspnea, cough, diarrhea, fever, chills | Died (10 d) due to multiorgan failure (renal, pulmonary, cardiac), acute CVA | Leukemia and bone marrow transplant (twice in 2014 and 2016) | Ruxolitinib, Methotrexate | Hypertension, asthma |
| Pt 6 | F (60–64) | 29 March 2021 | shortness of breath, cough, fatigue | Recovered (10 d) | Follicular lymphoma | R-CHOP (monoclonal antibody rituximab with cyclophosphamide, doxorubicin, vincristine, and prednisone) | Hypertension, asthma |
| Pt 7 | F (65–69) | 30 March 2021 | fever, myalgia, loss of taste, and tachypnea | Died (14 d) due to multiorgan failure (cardiac, pulmonary) | Pulmonary hypertension | NA | Hypothyroidism, hypertension, history rheumatic fever |
| Pt 8 | F (80–84) | 28 March 2021 | Malaise, sore throat, and cough | Recovered (11 d) | Breast cancer | Exemestane (aromatase inhibitors) | Nonischemic cardiomyopathy, s/p implantable cardioverter-defibrillator (ICD), hypertension |
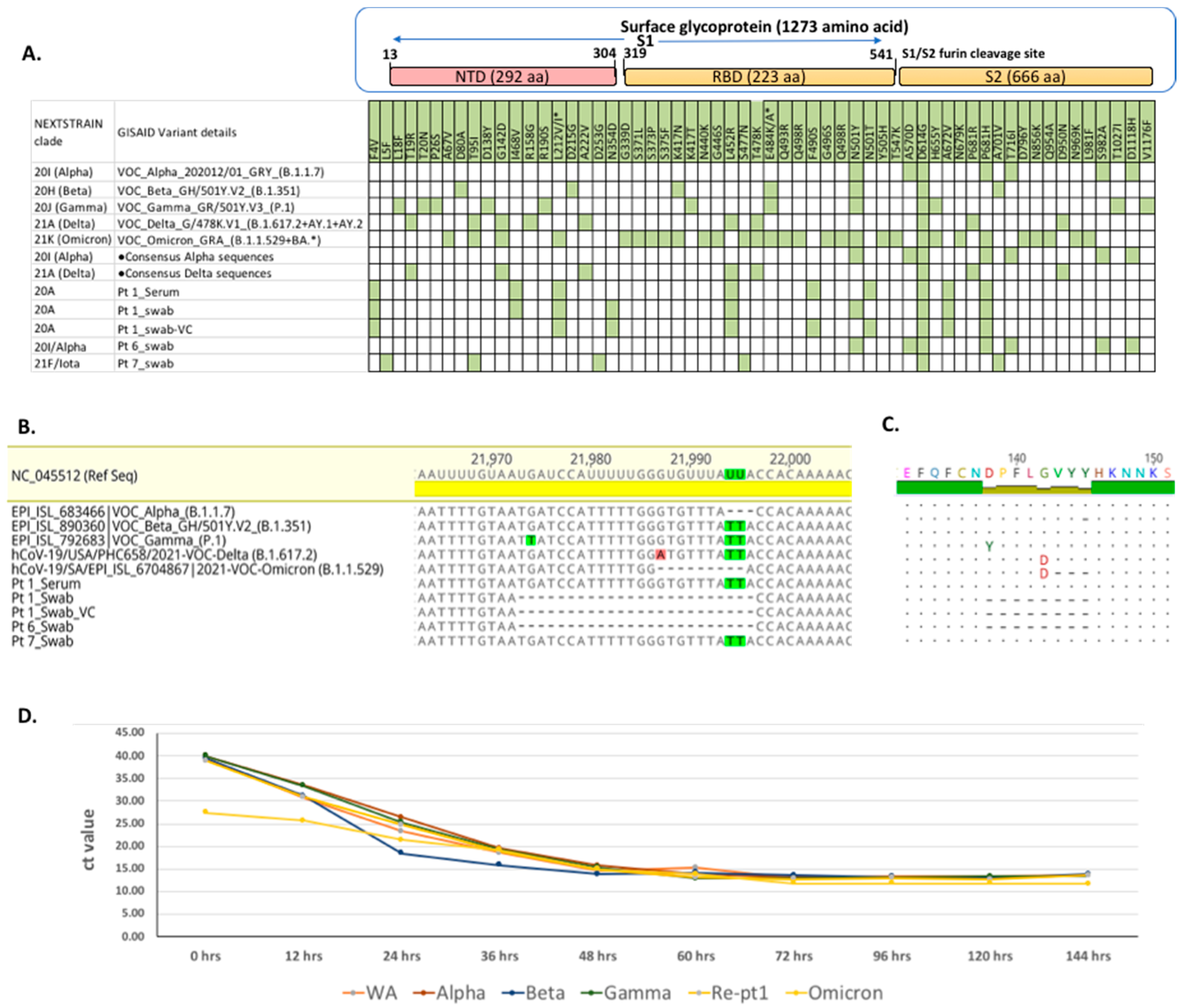
3.2. SARS-CoV-2 Genome Analyses
3.3. Serological Analyses
3.3.1. ELISA Assays
3.3.2. End-Point Absolute Viral Neutralization Assay
| EUROIMMUN ELISAs | Neutralizing Antibody Titers against Different SARS-CoV-2 Variants | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Patient ID | COVID-19 Outcome | Serum Collected (Days Post-Infection) | Surface Glycoprotein | Nucleocapsid | WA1 Variants | Alpha (B.1.1.7) | Gamma (P.1) | Beta (B.1.351) | Delta (B.1.617) | Omicron (B.1.1.529) | 20 A (Pt 1) |
| Pt 1 | Fatal | 04 | Negative | Positive | ≤1:20 | ≤1:20 | ≤1:20 | ≤1:20 | ≤1:20 | ≤1:20 | ≤1:20 |
| 10 | Negative | Positive | 1:80 | ≤1:20 | ≤1:20 | ≤1:20 | ≤1:20 | ≤1:20 | ≤1:20 | ||
| 17 | Negative | Positive | ≤1:20 | ≤1:20 | ≤1:20 | ≤1:20 | ≤1:20 | ≤1:20 | ≤1:20 | ||
| Pt 2 | Survival | 0 | Negative | Negative | ≤1:20 | ≤1:20 | ≤1:20 | ≤1:20 | ≤1:20 | ≤1:20 | ≤1:20 |
| 05 | Negative | Negative | ≤1:20 | ≤1:20 | ≤1:20 | ≤1:20 | ≤1:20 | ≤1:20 | ≤1:20 | ||
| Pt 3 | Fatal | 10 | Negative | Negative | ≤1:20 | ≤1:20 | ≤1:20 | ≤1:20 | ≤1:20 | ≤1:20 | ≤1:20 |
| 14 | Negative | Negative | ≤1:20 | ≤1:20 | ≤1:20 | ≤1:20 | ≤1:20 | ≤1:20 | ≤1:20 | ||
| Pt 4 | Survival | 0 | Negative | Negative | ≤1:20 | ≤1:20 | ≤1:20 | ≤1:20 | ≤1:20 | ≤1:20 | ≤1:20 |
| 04 | Negative | Negative | ≤1:20 | ≤1:20 | ≤1:20 | ≤1:20 | ≤1:20 | ≤1:20 | ≤1:20 | ||
| Pt 5 | Fatal | 05 | Positive | Positive | 1:320 | ≤1:20 | ≤1:20 | ≤1:20 | ≤1:20 | ≤1:20 | ≤1:20 |
| 10 | Positive | Positive | 1:1280 | 1:640 | ≤1:20 | 1:40 | ≤1:20 | ≤1:20 | 1:80 | ||
| Pt 6 | Survival | 05 | Negative | Negative | ≤1:20 | ≤1:20 | ≤1:20 | ≤1:20 | ≤1:20 | ≤1:20 | ≤1:20 |
| 10 | Negative | Negative | ≤1:20 | ≤1:20 | ≤1:20 | ≤1:20 | ≤1:20 | ≤1:20 | ≤1:20 | ||
| Pt 7 | Fatal | 09 | Positive | Positive | 1:160 | ≤1:20 | ≤1:20 | ≤1:20 | ≤1:20 | ≤1:20 | 1:80 |
| 14 | Positive | Positive | 1:640 | ≤1:20 | 1:40 | ≤1:20 | ≤1:20 | ≤1:20 | 1:160 | ||
| Pt 8 | Survival | 17 | Positive | Positive | 1:1280 | 1:160 | 1:320 | ≤1:20 | ≤1:20 | ≤1:20 | 1:40 |
| 22 | Positive | Positive | 1:640 | 1:320 | 1:640 | 1:80 | ≤1:20 | ≤1:20 | ≤1:20 | ||
| CON-0132 | Survived | 37 | Positive | Negative | 1:40 | ≤1:20 | NA | NA | 1:40 | <1:20 | 1:320 |
| CON-0133 | Survived | 8 | Positive | Negative | 1:80 | 1:160 | NA | NA | 1:80 | 1:40 | 1:160 |
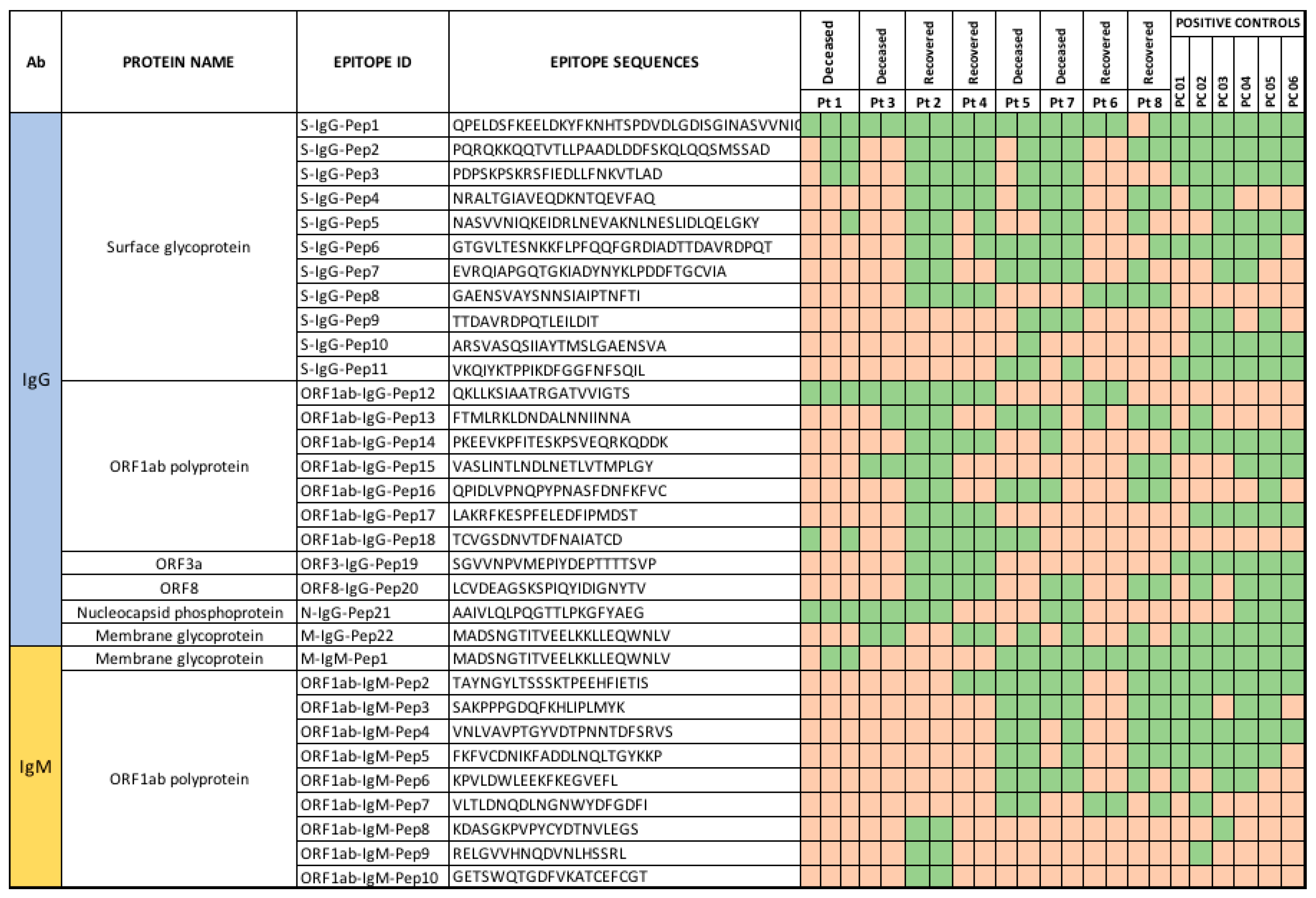
3.3.3. Peptide Array Analysis
4. Discussion
Limitations of the Study
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Fung, M.; Babik, J.M. COVID-19 in Immunocompromised Hosts: What We Know So Far. Clin. Infect. Dis. 2021, 72, 340–350. [Google Scholar] [CrossRef] [PubMed]
- Boyarsky, B.J.; Werbel, W.A.; Avery, R.K.; Tobian, A.A.R.; Massie, A.B.; Segev, D.L.; Garonzik-Wang, J.M. Antibody Response to 2-Dose SARS-CoV-2 mRNA Vaccine Series in Solid Organ Transplant Recipients. JAMA 2021, 325, 2204–2206. [Google Scholar] [CrossRef] [PubMed]
- Boyarsky, B.J.; Werbel, W.A.; Avery, R.K.; Tobian, A.A.R.; Massie, A.B.; Segev, D.L.; Garonzik-Wang, J.M. Immunogenicity of a Single Dose of SARS-CoV-2 Messenger RNA Vaccine in Solid Organ Transplant Recipients. JAMA 2021, 325, 1784–1786. [Google Scholar] [CrossRef] [PubMed]
- Walsh, E.E.; Frenck, R.W., Jr.; Falsey, A.R.; Kitchin, N.; Absalon, J.; Gurtman, A.; Lockhart, S.; Neuzil, K.; Mulligan, M.J.; Bailey, R.; et al. Safety and Immunogenicity of Two RNA-Based Covid-19 Vaccine Candidates. N. Engl. J. Med. 2020, 383, 2439–2450. [Google Scholar] [CrossRef]
- Jackson, L.A.; Anderson, E.J.; Rouphael, N.G.; Roberts, P.C.; Makhene, M.; Coler, R.N.; McCullough, M.P.; Chappell, J.D.; Denison, M.R.; Stevens, L.J.; et al. An mRNA Vaccine against SARS-CoV-2-Preliminary Report. N. Engl. J. Med. 2020, 383, 1920–1931. [Google Scholar] [CrossRef]
- Dashdorj, N.J.; Dashdorj, N.D.; Mishra, M.; Danzig, L.; Briese, T.; Lipkin, W.I.; Mishra, N. Molecular and serological investigation of the 2021 COVID-19 case surge in Mongolian vaccinees. medRxiv 2021. [Google Scholar] [CrossRef]
- Garcia-Beltran, W.F.; St Denis, K.J.; Hoelzemer, A.; Lam, E.C.; Nitido, A.D.; Sheehan, M.L.; Berrios, C.; Ofoman, O.; Chang, C.C.; Hauser, B.M.; et al. mRNA-based COVID-19 vaccine boosters induce neutralizing immunity against SARS-CoV-2 Omicron variant. Cell 2022, 185, 457–466.e4. [Google Scholar] [CrossRef]
- Shroff, R.T.; Chalasani, P.; Wei, R.; Pennington, D.; Quirk, G.; Schoenle, M.V.; Peyton, K.L.; Uhrlaub, J.L.; Ripperger, T.J.; Jergović, M.; et al. Immune responses to two and three doses of the BNT162b2 mRNA vaccine in adults with solid tumors. Nat. Med. 2021, 27, 2002–2011. [Google Scholar] [CrossRef]
- Avanzato, V.A.; Matson, M.J.; Seifert, S.N.; Pryce, R.; Williamson, B.N.; Anzick, S.L.; Barbian, K.; Judson, S.D.; Fischer, E.R.; Martens, C.; et al. Case Study: Prolonged Infectious SARS-CoV-2 Shedding from an Asymptomatic Immunocompromised Individual with Cancer. Cell 2020, 183, 1901–1912.e9. [Google Scholar] [CrossRef]
- Truong, T.T.; Ryutov, A.; Pandey, U.; Yee, R.; Goldberg, L.; Bhojwani, D.; Aguayo-Hiraldo, P.; Pinsky, B.A.; Pekosz, A.; Shen, L.; et al. Persistent SARS-CoV-2 infection and increasing viral variants in children and young adults with impaired humoral immunity. medRxiv 2021, 67, 103355. [Google Scholar]
- Choi, B.; Choudhary, M.C.; Regan, J.; Sparks, J.A.; Padera, R.F.; Qiu, X.; Solomon, I.H.; Kuo, H.H.; Boucau, J.; Bowman, K.; et al. Persistence and Evolution of SARS-CoV-2 in an Immunocompromised Host. N. Engl. J. Med. 2020, 383, 2291–2293. [Google Scholar] [CrossRef] [PubMed]
- CUIMC. Columbia University Laboratory of Personalized Genomic Medicine-Triplex CII-SARS-CoV-2 rRT-PCR Test. In Vitro Diagnostic Use. 2020. Available online: https://www.fda.gov/media/137983/download (accessed on 31 March 2022).
- Mishra, N.; Ng, T.F.F.; Marine, R.L.; Jain, K.; Ng, J.; Thakkar, R.; Caciula, A.; Price, A.; Garcia, J.A.; Burns, J.C.; et al. Antibodies to Enteroviruses in Cerebrospinal Fluid of Patients with Acute Flaccid Myelitis. MBio 2019, 10, e01903–e01919. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- GISAID. Tracking of Variants. Available online: https://www.gisaid.org/hcov19-variants (accessed on 15 September 2021).
- Next-Strain. Real-Time Tracking of Pathogen Evolution. Available online: https://nextstrain.org (accessed on 15 September 2021).
- Mishra, N.; Huang, X.; Joshi, S.; Guo, C.; Ng, J.; Thakkar, R.; Wu, Y.; Dong, X.; Li, Q.; Pinapati, R.S.; et al. Immunoreactive peptide maps of SARS-CoV-2. Commun. Biol. 2021, 4, 225. [Google Scholar] [CrossRef] [PubMed]
- Mishra, N.; Caciula, A.; Price, A.; Thakkar, R.; Ng, J.; Chauhan, L.V.; Jain, K.; Che, X.; Espinosa, D.A.; Montoya Cruz, M.; et al. Diagnosis of Zika Virus Infection by Peptide Array and Enzyme-Linked Immunosorbent Assay. MBio 2018, 9, e00095-18. [Google Scholar] [CrossRef] [Green Version]
- Tokarz, R.; Mishra, N.; Tagliafierro, T.; Sameroff, S.; Caciula, A.; Chauhan, L.; Patel, J.; Sullivan, E.; Gucwa, A.; Fallon, B.; et al. A multiplex serologic platform for diagnosis of tick-borne diseases. Sci. Rep. 2018, 8, 3158. [Google Scholar] [CrossRef] [Green Version]
- Tokarz, R.; Tagliafierro, T.; Caciula, A.; Mishra, N.; Thakkar, R.; Chauhan, L.V.; Sameroff, S.; Delaney, S.; Wormser, G.P.; Marques, A.; et al. Identification of immunoreactive linear epitopes of Borrelia miyamotoi. Ticks Tick-Borne Dis. 2020, 11, 101314. [Google Scholar] [CrossRef]
- Buus, S.; Rockberg, J.; Forsström, B.; Nilsson, P.; Uhlen, M.; Schafer-Nielsen, C. High-resolution mapping of linear antibody epitopes using ultrahigh-density peptide microarrays. Mol. Cell Proteom. 2012, 11, 1790–1800. [Google Scholar] [CrossRef] [Green Version]
- Valentini, D.; Rao, M.; Rane, L.; Rahman, S.; Axelsson-Robertson, R.; Heuchel, R.; Löhr, M.; Hoft, D.; Brighenti, S.; Zumla, A.; et al. Peptide microarray-based characterization of antibody responses to host proteins after bacille Calmette-Guerin vaccination. Int. J. Infect. Dis. 2017, 56, 140–154. [Google Scholar] [CrossRef] [Green Version]
- Tavakolpour, S.; Rakhshandehroo, T.; Wei, E.X.; Rashidian, M. Lymphopenia during the COVID-19 infection: What it shows and what can be learned. Immunol. Lett. 2020, 225, 31–32. [Google Scholar] [CrossRef]
- Tan, L.; Wang, Q.; Zhang, D.; Ding, J.; Huang, Q.; Tang, Y.Q.; Wang, Q.; Miao, H. Lymphopenia predicts disease severity of COVID-19: A descriptive and predictive study. Signal Transduct. Target. Ther. 2020, 5, 33. [Google Scholar]
- Dashdorj, N.J.; Dashdorj, N.D.; Mishra, M.; Danzig, L.; Briese, T.; Lipkin, W.I.; Mishra, N. Molecular and Serologic Investigation of the 2021 COVID-19 Case Surge among Vaccine Recipients in Mongolia. JAMA Netw. Open 2022, 5, e2148415. [Google Scholar] [CrossRef]
- Shobayo, B.; Mishra, M.; Sameroff, S.; Petrosov, A.; Ng, J.; Gokden, A.; MaCauley, J.; Jain, K.; Renken, C.; Duworko, J.T.; et al. SARS-CoV-2 Sequence Analysis during COVID-19 Case Surge, Liberia, 2021. Emerg. Infect. Dis. 2021, 27, 3185–3188. [Google Scholar] [CrossRef]
- Poh, C.M.; Carissimo, G.; Wang, B.; Amrun, S.N.; Lee, C.Y.; Chee, R.S.; Fong, S.W.; Yeo, N.K.; Lee, W.H.; Torres-Ruesta, A.; et al. Two linear epitopes on the SARS-CoV-2 spike protein that elicit neutralising antibodies in COVID-19 patients. Nat. Commun. 2020, 11, 2806. [Google Scholar] [CrossRef] [PubMed]
- Almaghrabi, R.S.; Alhamlan, F.S.; Dada, A.; Al-Tawfiq, J.A.; Al Hroub, M.K.; Saeedi, M.F.; Alamri, M.; Alhothaly, B.; Alqasabi, A.; Al-Qahtani, A.A.; et al. Outcome of SARS-CoV-2 variant breakthrough infection in fully immunized solid organ transplant recipients. J. Infect. Public Health 2022, 15, 51–55. [Google Scholar] [CrossRef]
- Lafarge, A.; Mabrouki, A.; Yvin, E.; Bredin, S.; Binois, Y.; Clere-Jehl, R.; Azoulay, E. Coronavirus disease 2019 in immunocompromised patients: A comprehensive review of coronavirus disease 2019 in hematopoietic stem cell recipients. Curr. Opin. Crit. Care 2022, 28, 83–89. [Google Scholar] [CrossRef] [PubMed]
- Puckett, Y.; Gabbar, A.; Bokhari, A.A. Prednisone. In StatPearls; StatPearls Publishing: Treasure Island, FL, USA, 2022. [Google Scholar]
- Barshes, N.R.; Goodpastor, S.E.; Goss, J.A. Pharmacologic immunosuppression. Front Biosci. 2004, 9, 411–420. [Google Scholar] [CrossRef] [Green Version]
- Allison, A.C. Mechanisms of action of mycophenolate mofetil. Lupus 2005, 14 (Suppl. 1), s2–s8. [Google Scholar] [CrossRef] [PubMed]
- Elli, E.M.; Baratè, C.; Mendicino, F.; Palandri, F.; Palumbo, G.A. Mechanisms Underlying the Anti-inflammatory and Immunosuppressive Activity of Ruxolitinib. Front Oncol. 2019, 9, 1186. [Google Scholar] [CrossRef] [Green Version]
- Parampalli Yajnanarayana, S.; Stübig, T.; Cornez, I.; Alchalby, H.; Schönberg, K.; Rudolph, J.; Triviai, I.; Wolschke, C.; Heine, A.; Brossart, P.; et al. JAK1/2 inhibition impairs T cell function in vitro and in patients with myeloproliferative neoplasms. Br. J. Haematol. 2015, 169, 824–833. [Google Scholar] [CrossRef] [PubMed]
- Taneja, V. Sex Hormones Determine Immune Response. Front Immunol. 2018, 9, 1931. [Google Scholar] [CrossRef]
- Mueller, A.L.; McNamara, M.S.; Sinclair, D.A. Why does COVID-19 disproportionately affect older people? Aging 2020, 12, 9959–9981. [Google Scholar] [CrossRef]
- Pu, S.L.; Zhang, X.Y.; Liu, D.S.; Ye, B.N.; Li, J.Q. Unexplained elevation of erythrocyte sedimentation rate in a patient recovering from COVID-19: A case report. World J. Clin. Cases 2021, 9, 1394–1401. [Google Scholar] [CrossRef] [PubMed]
- Hu, R.; Han, C.; Pei, S.; Yin, M.; Chen, X. Procalcitonin levels in COVID-19 patients. Int. J. Antimicrob. Agents 2020, 56, 106051. [Google Scholar] [CrossRef]
- Liu, F.; Li, L.; Xu, M.; Wu, J.; Luo, D.; Zhu, Y.; Li, B.; Song, X.; Zhou, X. Prognostic value of interleukin-6, C-reactive protein, and procalcitonin in patients with COVID-19. J. Clin. Virol. 2020, 127, 104370. [Google Scholar] [CrossRef] [PubMed]
- Gaitzsch, E.; Passerini, V.; Khatamzas, E.; Strobl, C.D.; Muenchhoff, M.; Scherer, C.; Osterman, A.; Heide, M.; Reischer, A.; Subklewe, M.; et al. COVID-19 in Patients Receiving CD20-depleting Immunochemotherapy for B-cell Lymphoma. Hemasphere 2021, 5, e603. [Google Scholar] [CrossRef] [PubMed]
- Boyarsky, B.J.; Ruddy, J.A.; Connolly, C.M.; Ou, M.T.; Werbel, W.A.; Garonzik-Wang, J.M.; Segev, D.L.; Paik, J.J. Antibody response to a single dose of SARS-CoV-2 mRNA vaccine in patients with rheumatic and musculoskeletal diseases. Ann. Rheum Dis. 2021, 80, 1098–1099. [Google Scholar] [CrossRef]
- Aydillo, T.; Gonzalez-Reiche, A.S.; Aslam, S.; van de Guchte, A.; Khan, Z.; Obla, A.; Dutta, J.; van Bakel, H.; Aberg, J.; García-Sastre, A.; et al. Shedding of Viable SARS-CoV-2 after Immunosuppressive Therapy for Cancer. N. Engl. J. Med. 2020, 383, 2586–2588. [Google Scholar] [CrossRef]
- Fendler, A.; de Vries, E.G.E.; GeurtsvanKessel, C.H.; Haanen, J.B.; Wörmann, B.; Turajlic, S.; von Lilienfeld-Toal, M. COVID-19 vaccines in patients with cancer: Immunogenicity, efficacy and safety. Nat. Rev. Clin. Oncol. 2022, 11, 1–17. [Google Scholar] [CrossRef]
- Harvey, W.T.; Carabelli, A.M.; Jackson, B.; Gupta, R.K.; Thomson, E.C.; Harrison, E.M.; Ludden, C.; Reeve, R.; Rambaut, A. SARS-CoV-2 variants, spike mutations and immune escape. Nat. Rev. Microbiol. 2021, 19, 409–424. [Google Scholar] [CrossRef]
- Moss, P. The T cell immune response against SARS-CoV-2. Nat. Immunol. 2022, 23, 186–193. [Google Scholar] [CrossRef]
- Baden, L.R.; El Sahly, H.M.; Essink, B.; Kotloff, K.; Frey, S.; Novak, R.; Diemert, D.; Spector, S.A.; Rouphael, N.; Creech, C.B.; et al. Efficacy and Safety of the mRNA-1273 SARS-CoV-2 Vaccine. N. Engl. J. Med. 2021, 384, 403–416. [Google Scholar] [CrossRef] [PubMed]
- Polack, F.P.; Thomas, S.J.; Kitchin, N.; Absalon, J.; Gurtman, A.; Lockhart, S.; Perez, J.L.; Pérez Marc, G.; Moreira, E.D.; Zerbini, C.; et al. Safety and Efficacy of the BNT162b2 mRNA Covid-19 Vaccine. N. Engl. J. Med. 2020, 383, 2603–2615. [Google Scholar] [CrossRef] [PubMed]
- McCallum, M.; De Marco, A.; Lempp, F.A.; Tortorici, M.A.; Pinto, D.; Walls, A.C.; Beltramello, M.; Chen, A.; Liu, Z.; Zatta, F.; et al. N-terminal domain antigenic mapping reveals a site of vulnerability for SARS-CoV-2. Cell 2021, 184, 2332–2347.e16. [Google Scholar] [CrossRef] [PubMed]
- Heffron, A.S.; McIlwain, S.J.; Amjadi, M.F.; Baker, D.A.; Khullar, S.; Armbrust, T.; Halfmann, P.J.; Kawaoka, Y.; Sethi, A.K.; Palmenberg, A.C.; et al. The landscape of antibody binding in SARS-CoV-2 infection. PLoS Biol. 2021, 19, e3001265. [Google Scholar] [CrossRef]



Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mishra, M.; Zahra, A.; Chauhan, L.V.; Thakkar, R.; Ng, J.; Joshi, S.; Spitzer, E.D.; Marcos, L.A.; Lipkin, W.I.; Mishra, N. A Short Series of Case Reports of COVID-19 in Immunocompromised Patients. Viruses 2022, 14, 934. https://doi.org/10.3390/v14050934
Mishra M, Zahra A, Chauhan LV, Thakkar R, Ng J, Joshi S, Spitzer ED, Marcos LA, Lipkin WI, Mishra N. A Short Series of Case Reports of COVID-19 in Immunocompromised Patients. Viruses. 2022; 14(5):934. https://doi.org/10.3390/v14050934
Chicago/Turabian StyleMishra, Mitali, Aleena Zahra, Lokendra V. Chauhan, Riddhi Thakkar, James Ng, Shreyas Joshi, Eric D. Spitzer, Luis A. Marcos, W. Ian Lipkin, and Nischay Mishra. 2022. "A Short Series of Case Reports of COVID-19 in Immunocompromised Patients" Viruses 14, no. 5: 934. https://doi.org/10.3390/v14050934