
Rational Design of a Skin- and Neuro-Attenuated Live Varicella Vaccine: A Review and Future Perspectives


{kind=link}
Abstract
:1. Introduction
2. Current Status of Varicella Vaccine
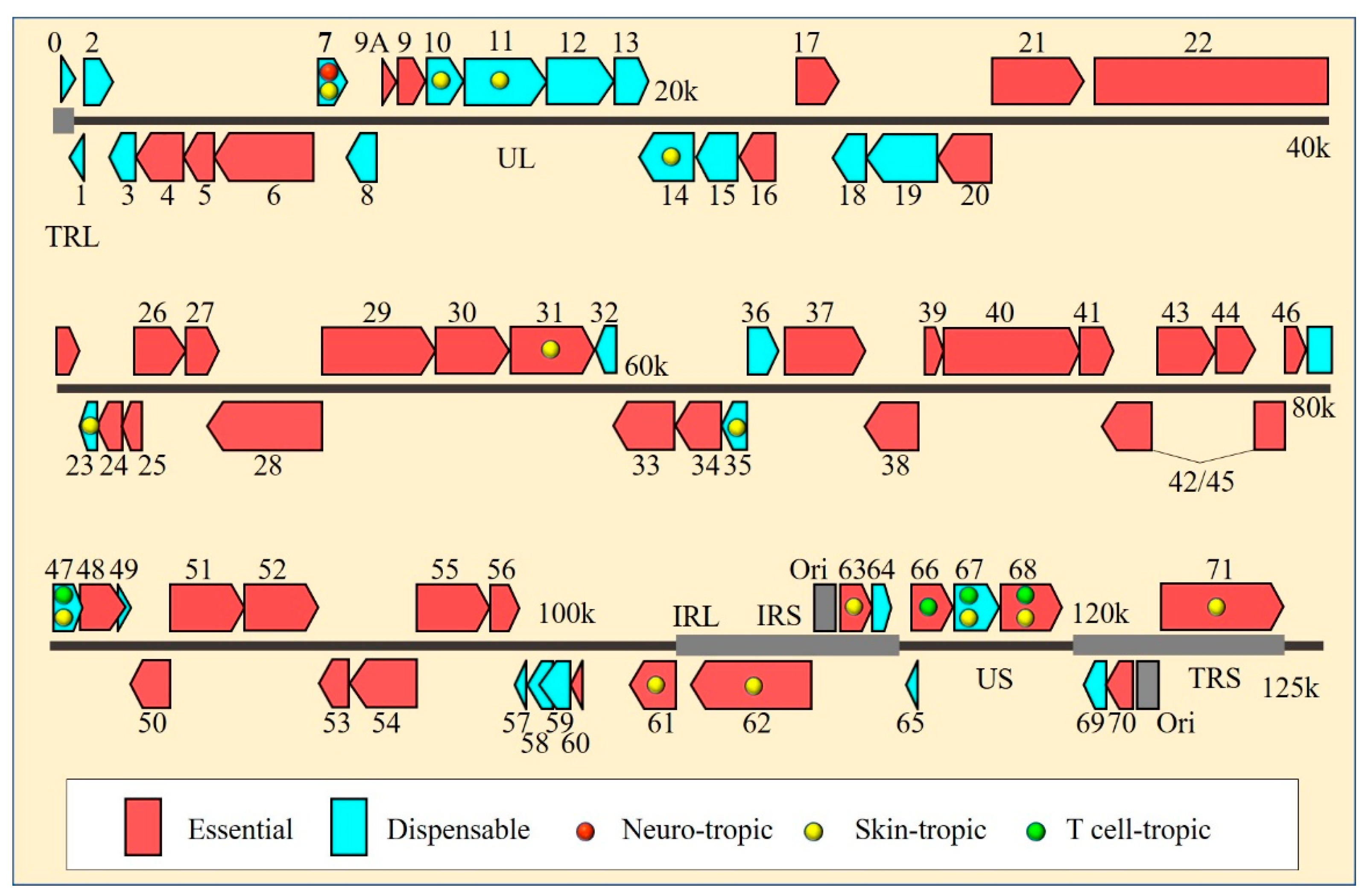
3. Identification of ORF7 as a Skin- and Neuro-Tropic Factor of VZV
4. Development of an ORF7-Deficient, Live-Attenuated Varicella Vaccine, v7D
5. Future Directions
5.1. Safety of the v7D Varicella Vaccine in Humans
5.2. Immunogenicity and Efficacy of the v7D Varicella Vaccine in Humans
5.3. Mechanisms of v7D Attenuation and Vaccine-Induced Protective Immunity
6. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Cohen, J.I.; Straus, S.E.; Arvin, A.M. Fields Virology, 5th ed.; Lippincott Williams & Wilkins: Philadelphia, PA, USA, 2007. [Google Scholar]
- Gilden, D.H.; Kleinschmidt-DeMasters, B.K.; LaGuardia, J.J.; Mahalingam, R.; Cohrs, R.J. Neurologic complications of the reactivation of varicella-zoster virus. N. Engl. J. Med. 2000, 342, 635–645. [Google Scholar] [CrossRef] [PubMed]
- Gnann, J.W., Jr. Varicella-zoster virus: Atypical presentations and unusual complications. J. Infect. Dis. 2002, 186 (Suppl. 1), S91–S98. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, M.; Otsuka, T.; Okuno, Y.; Asano, Y.; Yazaki, T.; Isomura, S. Live vaccine used to prevent the spread of varicella in children in hospital. Lancet 1974, 304, 1288–1290. [Google Scholar] [CrossRef]
- Takahashi, M.; Okuno, Y.; Otsuka, T.; Osame, J.; Takamizawa, A. Development of a live attenuated varicella vaccine. Biken J. 1975, 18, 25–33. [Google Scholar] [PubMed]
- Depledge, D.P.; Kundu, S.; Jensen, N.J.; Gray, E.R.; Jones, M.; Steinberg, S.; Gershon, A.; Kinchington, P.R.; Schmid, D.S.; Balloux, F.; et al. Deep sequencing of viral genomes provides insight into the evolution and pathogenesis of varicella zoster virus and its vaccine in humans. Mol. Biol. Evol. 2014, 31, 397–409. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Depledge, D.P.; Yamanishi, K.; Gomi, Y.; Gershon, A.A.; Breuer, J. Deep Sequencing of Distinct Preparations of the Live Attenuated Varicella-Zoster Virus Vaccine Reveals a Conserved Core of Attenuating Single-Nucleotide Polymorphisms. J. Virol. 2016, 90, 8698–8704. [Google Scholar] [CrossRef] [Green Version]
- Quinlivan, M.; Breuer, J. Clinical and molecular aspects of the live attenuated Oka varicella vaccine. Rev. Med. Virol. 2014, 24, 254–273. [Google Scholar] [CrossRef]
- Wolfson, L.J.; Daniels, V.J.; Altland, A.; Black, W.; Huang, W.; Ou, W. The Impact of Varicella Vaccination on the Incidence of Varicella and Herpes Zoster in the United States: Updated Evidence From Observational Databases, 1991–2016. Clin. Infect. Dis. 2020, 70, 995–1002. [Google Scholar] [CrossRef]
- Morino, S.; Tanaka-Taya, K.; Satoh, H.; Arai, S.; Takahashi, T.; Sunagawa, T.; Oishi, K. Descriptive epidemiology of varicella based on national surveillance data before and after the introduction of routine varicella vaccination with two doses in Japan, 2000–2017. Vaccine 2018, 36, 5977–5982. [Google Scholar] [CrossRef]
- Woodward, M.; Marko, A.; Galea, S.; Eagel, B.; Straus, W. Varicella Virus Vaccine Live: A 22-Year Review of Postmarketing Safety Data. In Open Forum Infectious Diseases; Oxford University Press: Oxford, UK, 2019; Volume 6. [Google Scholar] [CrossRef] [Green Version]
- Gershon, A.A.; Gershon, M.D.; Shapiro, E.D. Live Attenuated Varicella Vaccine: Prevention of Varicella and of Zoster. J. Infect. Dis. 2021, 224, S387–S397. [Google Scholar] [CrossRef]
- Harpaz, R. Do varicella vaccination programs change the epidemiology of herpes zoster? A comprehensive review, with focus on the United States. Expert Rev. Vaccines 2019, 18, 793–811. [Google Scholar] [CrossRef] [PubMed]
- Chaves, S.S.; Haber, P.; Walton, K.; Wise, R.P.; Izurieta, H.S.; Schmid, D.S.; Seward, J.F. Safety of varicella vaccine after licensure in the United States: Experience from reports to the vaccine adverse event reporting system, 1995–2005. J. Infect. Dis. 2008, 197 (Suppl. 2), S170–S177. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Galea, S.A.; Sweet, A.; Beninger, P.; Steinberg, S.P.; LaRussa, P.S.; Gershon, A.A.; Sharrar, R.G. The safety profile of varicella vaccine: A 10-year review. J. Infect. Dis. 2008, 197 (Suppl. 2), S165–S169. [Google Scholar] [CrossRef] [PubMed]
- Goulleret, N.; Mauvisseau, E.; Essevaz-Roulet, M.; Quinlivan, M.; Breuer, J. Safety profile of live varicella virus vaccine (Oka/Merck): Five-year results of the European Varicella Zoster Virus Identification Program (EU VZVIP). Vaccine 2010, 28, 5878–5882. [Google Scholar] [CrossRef]
- Yoshikawa, T.; Ando, Y.; Nakagawa, T.; Gomi, Y. Safety profile of the varicella vaccine (Oka vaccine strain) based on reported cases from 2005 to 2015 in Japan. Vaccine 2016, 34, 4943–4947. [Google Scholar] [CrossRef] [Green Version]
- Weinmann, S.; Chun, C.; Schmid, D.S.; Roberts, M.; Vandermeer, M.; Riedlinger, K.; Bialek, S.R.; Marin, M. Incidence and clinical characteristics of herpes zoster among children in the varicella vaccine era, 2005–2009. J. Infect. Dis. 2013, 208, 1859–1868. [Google Scholar] [CrossRef] [Green Version]
- Sharrar, R.G.; LaRussa, P.; Galea, S.A.; Steinberg, S.P.; Sweet, A.R.; Keatley, R.M.; Wells, M.E.; Stephenson, W.P.; Gershon, A.A. The postmarketing safety profile of varicella vaccine. Vaccine 2000, 19, 916–923. [Google Scholar] [CrossRef]
- Wise, R.P.; Salive, M.E.; Braun, M.M.; Mootrey, G.T.; Seward, J.F.; Rider, L.G.; Krause, P.R. Postlicensure safety surveillance for varicella vaccine. JAMA 2000, 284, 1271–1279. [Google Scholar] [CrossRef] [Green Version]
- Krause, P.R.; Klinman, D.M. Varicella vaccination: Evidence for frequent reactivation of the vaccine strain in healthy children. Nat. Med. 2000, 6, 451–454. [Google Scholar] [CrossRef]
- Heusel, E.H.; Grose, C. Twelve Children with Varicella Vaccine Meningitis: Neuropathogenesis of Reactivated Live Attenuated Varicella Vaccine Virus. Viruses 2020, 12, 1078. [Google Scholar] [CrossRef]
- Zerboni, L.; Ku, C.-C.; Jones, C.D.; Zehnder, J.L.; Arvin, A.M. Varicella-zoster virus infection of human dorsal root ganglia in vivo. Proc. Natl. Acad. Sci. USA 2005, 102, 6490–6495. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sadaoka, T.; Depledge, D.P.; Rajbhandari, L.; Venkatesan, A.; Breuer, J.; Cohen, J.I. In vitro system using human neurons demonstrates that varicella-zoster vaccine virus is impaired for reactivation, but not latency. Proc. Natl. Acad. Sci. USA 2016, 113, E2403–E2412. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weinmann, S.; Naleway, A.L.; Koppolu, P.; Baxter, R.; Belongia, E.A.; Hambidge, S.J.; Irving, S.A.; Jackson, M.L.; Klein, N.P.; Lewin, B.; et al. Incidence of Herpes Zoster Among Children: 2003–2014. Pediatrics 2019, 144, e20182917. [Google Scholar] [CrossRef] [PubMed]
- Civen, R.; Chaves, S.S.; Jumaan, A.; Wu, H.; Mascola, L.; Gargiullo, P.; Seward, J.F. The incidence and clinical characteristics of herpes zoster among children and adolescents after implementation of varicella vaccination. Pediatr. Infect. Dis. J. 2009, 28, 954–959. [Google Scholar] [CrossRef]
- L’Huillier, A.G.; Hirzel, C.; Ferreira, V.H.; Ierullo, M.; Ku, T.; Selzner, N.; Schiff, J.; Juvet, S.; Miao, C.; Schmid, D.S.; et al. Evaluation of Recombinant Herpes Zoster Vaccine for Primary Immunization of Varicella-seronegative Transplant Recipients. Transplantation 2021, 105, 2316–2323. [Google Scholar] [CrossRef]
- Levin, M.J.; Weinberg, A. Immune Responses to Varicella-Zoster Virus Vaccines. In Current Topics in Microbiology and Immunology; Springer: Berlin/Heidelberg, Germany, 2022. [Google Scholar] [CrossRef]
- Cohen, J.I.; Seidel, K.E. Generation of varicella-zoster virus (VZV) and viral mutants from cosmid DNAs: VZV thymidylate synthetase is not essential for replication in vitro. Proc. Natl. Acad. Sci. USA 1993, 90, 7376–7380. [Google Scholar] [CrossRef] [Green Version]
- Niizuma, T.; Zerboni, L.; Sommer, M.H.; Ito, H.; Hinchliffe, S.; Arvin, A.M. Construction of varicella-zoster virus recombinants from parent Oka cosmids and demonstration that ORF65 protein is dispensable for infection of human skin and T cells in the SCID-hu mouse model. J. Virol. 2003, 77, 6062–6065. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Z.; Huang, Y.; Zhu, H. A highly efficient protocol of generating and analyzing VZV ORF deletion mutants based on a newly developed luciferase VZV BAC system. J. Virol. Methods 2008, 148, 197–204. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Z.; Selariu, A.; Warden, C.; Huang, G.; Huang, Y.; Zaccheus, O.; Cheng, T.; Xia, N.; Zhu, H. Genome-wide mutagenesis reveals that ORF7 is a novel VZV skin-tropic factor. PLoS Pathog. 2010, 6, e1000971. [Google Scholar] [CrossRef] [Green Version]
- Selariu, A.; Cheng, T.; Tang, Q.; Silver, B.; Yang, L.; Liu, C.; Ye, X.; Markus, A.; Goldstein, R.S.; Cruz-Cosme, R.S.; et al. ORF7 of varicella-zoster virus is a neurotropic factor. J. Virol. 2012, 86, 8614–8624. [Google Scholar] [CrossRef] [Green Version]
- Zerboni, L.; Sen, N.; Oliver, S.L.; Arvin, A.M. Molecular mechanisms of varicella zoster virus pathogenesis. Nat. Rev. Microbiol. 2014, 12, 197–210. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Z.; Rowe, J.; Wang, W.; Sommer, M.; Arvin, A.; Moffat, J.; Zhu, H. Genetic analysis of varicella-zoster virus ORF0 to ORF4 by use of a novel luciferase bacterial artificial chromosome system. J. Virol. 2007, 81, 9024–9033. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Che, X.; Zerboni, L.; Sommer, M.H.; Arvin, A.M. Varicella-zoster virus open reading frame 10 is a virulence determinant in skin cells but not in T cells in vivo. J. Virol. 2006, 80, 3238–3248. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moffat, J.F.; Zerboni, L.; Kinchington, P.R.; Grose, C.; Kaneshima, H.; Arvin, A.M. Attenuation of the vaccine Oka strain of varicella-zoster virus and role of glycoprotein C in alphaherpesvirus virulence demonstrated in the SCID-hu mouse. J. Virol. 1998, 72, 965–974. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moffat, J.F.; Zerboni, L.; Sommer, M.H.; Heineman, T.C.; Cohen, J.I.; Kaneshima, H.; Arvin, A.M. The ORF47 and ORF66 putative protein kinases of varicella-zoster virus determine tropism for human T cells and skin in the SCID-hu mouse. Proc. Natl. Acad. Sci. USA 1998, 95, 11969–11974. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, W.; Pan, D.; Fu, W.; Ye, X.; Han, J.; Yang, L.; Jia, J.; Liu, J.; Zhu, R.; Zhang, Y.; et al. Development of a skin- and neuro-attenuated live vaccine for varicella. Nat. Commun. 2022, 13, 824. [Google Scholar] [CrossRef]
- Cohen, J.I.; Nguyen, H. Varicella-zoster virus ORF61 deletion mutants replicate in cell culture, but a mutant with stop codons in ORF61 reverts to wild-type virus. Virology 1998, 246, 306–316. [Google Scholar] [CrossRef] [Green Version]
- Weibel, R.E.; Kuter, B.J.; Neff, B.J.; Rothenberger, C.A.; Fitzgerald, A.J.; Connor, K.A.; Morton, D.; Mclean, A.A.; Scolnick, E.M. Live Oka/Merck Varicella Vaccine in Healthy-Children—Further Clinical and Laboratory Assessment. JAMA 1985, 254, 2435–2439. [Google Scholar] [CrossRef]
- Perella, D.; Wang, C.; Civen, R.; Viner, K.; Kuguru, K.; Daskalaki, I.; Schmid, D.S.; Lopez, A.S.; Tseng, H.F.; Newbern, E.C.; et al. Varicella Vaccine Effectiveness in Preventing Community Transmission in the 2-Dose Era. Pediatrics 2016, 137, e20152802. [Google Scholar] [CrossRef] [Green Version]
- Leung, J.; Lopez, A.S.; Blostein, J.; Thayer, N.; Zipprich, J.; Clayton, A.; Buttery, V.; Andersen, J.; Thomas, C.A.; Del Rosario, M.; et al. Impact of the US Two-dose Varicella Vaccination Program on the Epidemiology of Varicella Outbreaks: Data from Nine States, 2005–2012. Pediatr. Infect. Dis. J. 2015, 34, 1105–1109. [Google Scholar] [CrossRef]
- Jiang, H.-F.; Wang, W.; Jiang, X.; Zeng, W.-B.; Shen, Z.-Z.; Song, Y.-G.; Yang, H.; Liu, X.-J.; Dong, X.; Zhou, J.; et al. ORF7 of Varicella-Zoster Virus Is Required for Viral Cytoplasmic Envelopment in Differentiated Neuronal Cells. J. Virol. 2017, 91, e00127-17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grigoryan, S.; Kinchington, P.R.; Yang, I.H.; Selariu, A.; Zhu, H.; Yee, M.; Goldstein, R.S. Retrograde axonal transport of VZV: Kinetic studies in hESC-derived neurons. J. Neurovirol. 2012, 18, 462–470. [Google Scholar] [CrossRef] [Green Version]
- Wang, W.; Fu, W.; Pan, D.; Cai, L.; Ye, J.; Liu, J.; Liu, C.; Que, Y.; Xia, N.; Zhu, H.; et al. Varicella-zoster virus ORF7 interacts with ORF53 and plays a role in its trans-Golgi network localization. Virol. Sin. 2017, 32, 387–395. [Google Scholar] [CrossRef] [PubMed]
- Butt, B.G.; Owen, D.J.; Jeffries, C.M.; Ivanova, L.; Hill, C.H.; Houghton, J.W.; Ahmed, M.F.; Antrobus, R.; Svergun, D.I.; Welch, J.J.; et al. Insights into herpesvirus assembly from the structure of the pUL7:pUL51 complex. eLife 2020, 9, e53789. [Google Scholar] [CrossRef] [PubMed]
- Mahalingam, R.; Messaoudi, I.; Gilden, D. Simian varicella virus pathogenesis. Curr. Top. Microbiol. Immunol. 2010, 342, 309–321. [Google Scholar] [CrossRef] [Green Version]

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, W.; Pan, D.; Cheng, T.; Zhu, H. Rational Design of a Skin- and Neuro-Attenuated Live Varicella Vaccine: A Review and Future Perspectives. Viruses 2022, 14, 848. https://doi.org/10.3390/v14050848
Wang W, Pan D, Cheng T, Zhu H. Rational Design of a Skin- and Neuro-Attenuated Live Varicella Vaccine: A Review and Future Perspectives. Viruses. 2022; 14(5):848. https://doi.org/10.3390/v14050848
Chicago/Turabian StyleWang, Wei, Dequan Pan, Tong Cheng, and Hua Zhu. 2022. "Rational Design of a Skin- and Neuro-Attenuated Live Varicella Vaccine: A Review and Future Perspectives" Viruses 14, no. 5: 848. https://doi.org/10.3390/v14050848