
Deconstructing the Phage–Bacterial Biofilm Interaction as a Basis to Establish New Antibiofilm Strategies


Abstract
:1. Introduction
2. Biofilms as a Defense Mechanism against Phage Predation
2.1. The EPS Matrix as a Phage Adsorption Trap
2.2. Diffusion Inhibition
2.3. Physiological Refuges
2.4. Shielding Sensitive Bacteria by the ‘Wall Effect’
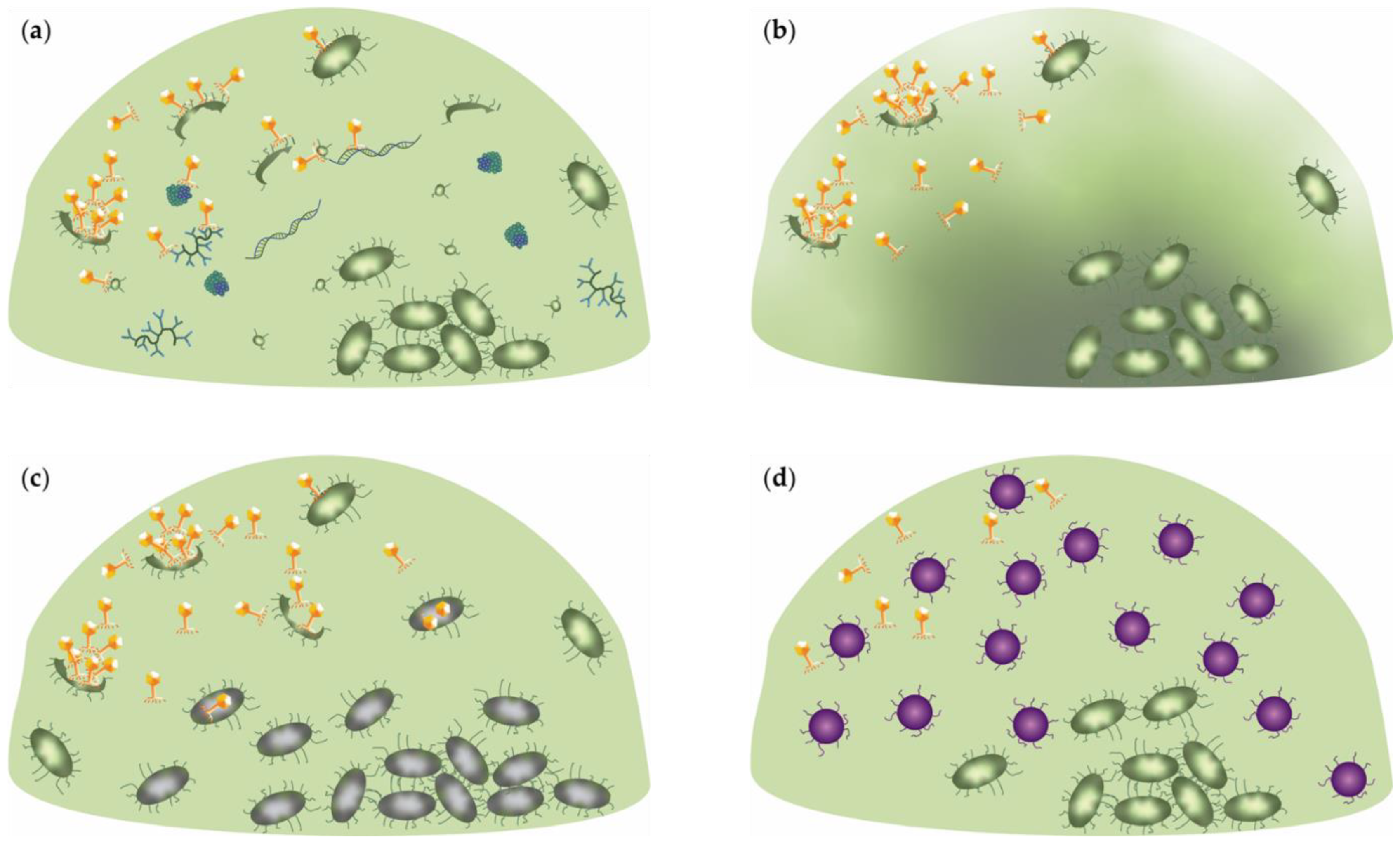
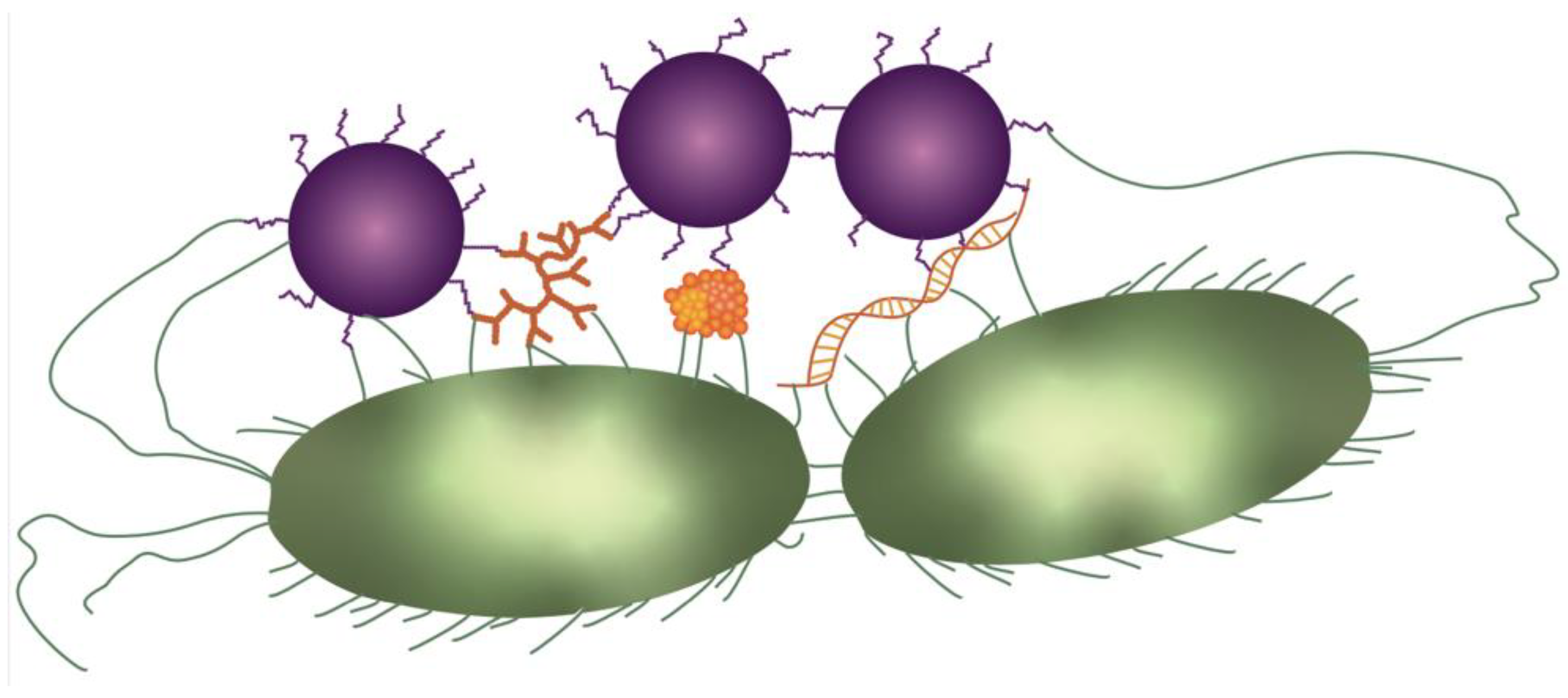
2.5. Phage Receptor-Driven Aggregation
2.6. Environment-Induced Mutations and Horizontal Gene Transfer as Drivers for Biofilm Diversity
3. Co-Existence of Bacteria and Phages within Biofilms
4. Phage Mechanisms to Overcome the Biofilm Barrier
4.1. Virion-Associated Exopolysaccharide Degrading Enzymes
4.2. Peptidoglycan Hydrolase Release
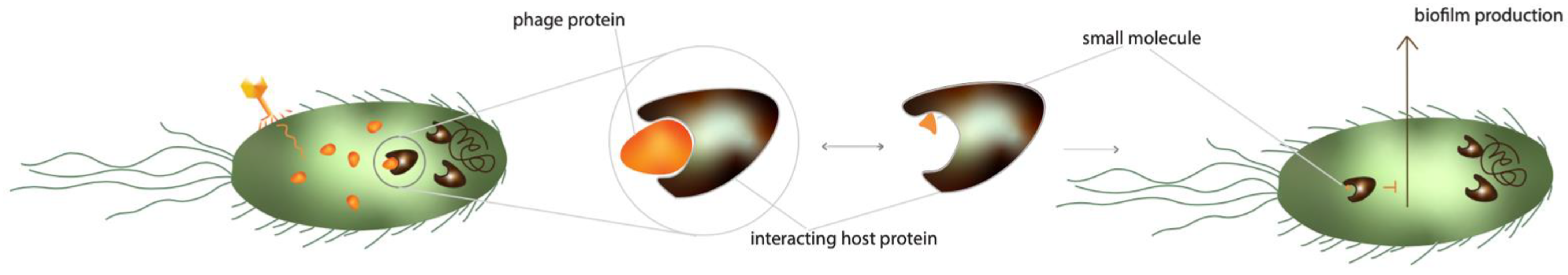
4.3. Phage-Mediated Inhibition of Cellular Communication
4.4. The Untapped Potential of Phage-Encoded DNases
5. The Search for Undiscovered Bacteriophage Antibiofilm Mechanisms
6. Conclusions and Outlook
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Fleming, A. Classics in Infectious Diseases: On the Antibacterial Action of Cultures of a Penicillium, with Special Reference to Their Use in the Isolation of B. Influenzae. Br. J. Exp. Pathol. 1929, 10, 226–236. [Google Scholar]
- Bjarnsholt, T. The Role of Bacterial Biofilms in Chronic Infections. Apmis 2013, 121, 1–51. [Google Scholar] [CrossRef] [PubMed]
- Costerton, J.W.; Geesey, G.G.; Cheng, K.J. How Bacteria Stick. Sci. Am. 1978, 238, 86–95. [Google Scholar] [CrossRef] [PubMed]
- Lewis, K.I.M. Riddle of Biofilm Resistance. Antimicrob. Agents Chemother. 2001, 45, 999–1007. [Google Scholar] [CrossRef] [Green Version]
- Kazemian, H.; Ghafourian, S.; Heidari, H.; Amiri, P.; Yamchi, J.K.; Shavalipour, A.; Houri, H.; Maleki, A.; Sadeghifard, N. Antibacterial, Anti-Swarming and Anti-Biofilm Formation Activities of Chamaemelum Nobile against Pseudomonas aeruginosa. Rev. Soc. Bras. Med. Trop. 2015, 48, 432–436. [Google Scholar] [CrossRef] [Green Version]
- Costerton, J.; Lewandowski, Z.; Caldwell, D.; Korber, D.; Lapin-Scott, H. Microbial Biofilms. Annu. Rev. Microbiol. 1995, 49, 711–745. [Google Scholar] [CrossRef]
- Flemming, H.C.; Wingender, J. The Biofilm Matrix. Nat. Rev. Microbiol. 2010, 8, 623–633. [Google Scholar] [CrossRef]
- Spiers, A.J. A Mechanistic Explanation Linking Adaptive Mutation, Niche Change, and Fitness Advantage for the Wrinkly Spreader. Int. J. Evol. Biol. 2014, 2014, 675432. [Google Scholar] [CrossRef] [Green Version]
- Chang, C.Y. Surface Sensing for Biofilm Formation in Pseudomonas Aeruginosa. Front. Microbiol. 2018, 8, 2671. [Google Scholar] [CrossRef]
- Konopka, A. What Is Microbial Community Ecology? ISME J. 2009, 3, 1223–1230. [Google Scholar] [CrossRef] [Green Version]
- Donlan, R.M.; Costerton, J.W. Biofilms: Survival Mechanisms of Clinically Relevant Microorganisms. Clin. Microbiol. Rev. 2002, 15, 167–193. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jefferson, K.K. What Drives Bacteria to Produce a Biofilm? FEMS Microbiol. Lett. 2004, 236, 163–173. [Google Scholar] [CrossRef] [PubMed]
- Fey, P.D.; Olson, M.E. Current Concepts in Biofilm Formation of Staphylococcus Epidermidis. Future Microbiol. 2010, 5, 917–933. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McCrea, K.W.; Hartford, O.; Davis, S.; Eidhin, D.N.; Lina, G.; Speziale, P.; Foster, T.J.; Höök, M. The Serine-Aspartate Repeat (Sdr) Protein Family in Staphylococcus Epidermidis. Microbiology 2000, 146, 1535–1546. [Google Scholar] [CrossRef] [PubMed]
- Williams, R.J.; Henderson, B.; Sharp, L.J.; Nair, S.P. Identification of a Fibronectin-Binding Protein from Staphylococcus Epidermidis. Infect. Immun. 2002, 70, 6805–6810. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hussain, M.; Heilmann, C.; Peters, G.; Herrmann, M. Teichoic Acid Enhances Adhesion of Staphylococcus Epidermidis to Immobilized Fibronectin. Microb. Pathog. 2001, 31, 261–270. [Google Scholar] [CrossRef]
- Conlon, B.P.; Geoghegan, J.A.; Waters, E.M.; McCarthy, H.; Rowe, S.E.; Davies, J.R.; Schaeffer, C.R.; Foster, T.J.; Fey, P.D.; O’Gara, J.P. Role for the A Domain of Unprocessed Accumulation-Associated Protein (Aap) in the Attachment Phase of the Staphylococcus Epidermidis Biofilm Phenotype. J. Bacteriol. 2014, 196, 4268–4275. [Google Scholar] [CrossRef] [Green Version]
- Qin, Z.; Ou, Y.; Yang, L.; Zhu, Y.; Tolker-Nielsen, T.; Molin, S.; Qu, D. Role of Autolysin-Mediated DNA Release in Biofilm Formation of Staphylococcus Epidermidis. Microbiology 2007, 153, 2083–2092. [Google Scholar] [CrossRef] [Green Version]
- O’Toole, G.A.; Kolter, R. Flagellar and Twitching Motility Are Necessary for Pseudomonas Aeruginosa Biofilm Development. Mol. Microbiol. 1998, 30, 295–304. [Google Scholar] [CrossRef]
- Ha, D.-G.; O’Toole, G.A. C-Di-GMP and Its Effects on Biofilm Formation and Dispersion: A Pseudomonas Aeruginosa Review. Microbiol. Spectr. 2015, 3, MB-0003-2014. [Google Scholar] [CrossRef] [Green Version]
- Vasseur, P.; Vallet-Gely, I.; Soscia, C.; Genin, S.; Filloux, A. The Pel Genes of the Pseudomonas Aeruginosa PAK Strain Are Involved at Early and Late Stages of Biofilm Formation. Microbiology 2005, 151, 985–997. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ma, L.; Jackson, K.D.; Landry, R.M.; Parsek, M.R.; Wozniak, D.J. Analysis of Pseudomonas Aeruginosa Conditional Psl Variants Reveals Roles for the Psl Polysaccharide in Adhesion and Maintaining Biofilm Structure Postattachment. J. Bacteriol. 2006, 188, 8213–8221. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Colvin, K.M.; Irie, Y.; Tart, C.S.; Urbano, R.; Whitney, J.C.; Ryder, C.; Howell, P.L.; Wozniak, D.J.; Parsek, M.R. The Pel and Psl Polysaccharides Provide Pseudomonas Aeruginosa Structural Redundancy within the Biofilm Matrix. Environ. Microbiol. 2012, 14, 1913–1928. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vallet, I.; Olson, J.W.; Lory, S.; Lazdunski, A.; Filloux, A. The Chaperone/Usher Pathways of Pseudomonas Aeruginosa: Identification of Fimbrial Gene Clusters (Cup) and Their Involvement in Biofilm Formation. Proc. Natl. Acad. Sci. USA 2001, 98, 6911–6916. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rohde, H.; Frankenberger, S.; Zähringer, U.; Mack, D. Structure, Function and Contribution of Polysaccharide Intercellular Adhesin (PIA) to Staphylococcus Epidermidis Biofilm Formation and Pathogenesis of Biomaterial-Associated Infections. Eur. J. Cell Biol. 2010, 89, 103–111. [Google Scholar] [CrossRef] [PubMed]
- Christner, M.; Franke, G.C.; Schommer, N.N.; Wendt, U.; Wegert, K.; Pehle, P.; Kroll, G.; Schulze, C.; Buck, F.; MacK, D.; et al. The Giant Extracellular Matrix-Binding Protein of Staphylococcus Epidermidis Mediates Biofilm Accumulation and Attachment to Fibronectin. Mol. Microbiol. 2010, 75, 187–207. [Google Scholar] [CrossRef] [PubMed]
- Rohde, H.; Burdelski, C.; Bartscht, K.; Hussain, M.; Buck, F.; Horstkotte, M.A.; Knobloch, J.K.M.; Heilmann, C.; Herrmann, M.; Mack, D. Induction of Staphylococcus Epidermidis Biofilm Formation via Proteolytic Processing of the Accumulation-Associated Protein by Staphylococcal and Host Proteases. Mol. Microbiol. 2005, 55, 1883–1895. [Google Scholar] [CrossRef]
- Christner, M.; Heinze, C.; Busch, M.; Franke, G.; Hentschke, M.; Bayard Dühring, S.; Büttner, H.; Kotasinska, M.; Wischnewski, V.; Kroll, G.; et al. SarA Negatively Regulates Staphylococcus Epidermidis Biofilm Formation by Modulating Expression of 1MDa Extracellular Matrix Binding Protein and Autolysis-Dependent Release of EDNA. Mol. Microbiol. 2012, 86, 394–410. [Google Scholar] [CrossRef]
- Büttner, H.; Mack, D.; Rohde, H.; Arciola, C.R. Structural Basis of Staphylococcus Epidermidis Biofilm Formation: Mechanisms and Molecular Interactions. Front. Cell. Infect. Microbiol. 2015, 5, 14. [Google Scholar] [CrossRef] [Green Version]
- Das, T.; Sehar, S.; Koop, L.; Wong, Y.K.; Ahmed, S.; Siddiqui, K.S.; Manefield, M. Influence of Calcium in Extracellular DNA Mediated Bacterial Aggregation and Biofilm Formation. PLoS ONE 2014, 9, e91935. [Google Scholar] [CrossRef] [Green Version]
- Ma, L.; Conover, M.; Lu, H.; Parsek, M.R.; Bayles, K.; Wozniak, D.J. Assembly and Development of the Pseudomonas Aeruginosa Biofilm Matrix. PLoS Pathog. 2009, 5, e1000354. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Colvin, K.M.; Gordon, V.D.; Murakami, K.; Borlee, B.R.; Wozniak, D.J.; Wong, G.C.L.; Parsek, M.R. The Pel Polysaccharide Can Serve a Structural and Protective Role in the Biofilm Matrix of Pseudomonas Aeruginosa. PLoS Pathog. 2011, 7, e1001264. [Google Scholar] [CrossRef] [PubMed]
- Jennings, L.K.; Storek, K.M.; Ledvina, H.E.; Coulon, C.; Marmont, L.S.; Sadovskaya, I.; Secor, P.R.; Tseng, B.S.; Scian, M.; Filloux, A.; et al. Pel Is a Cationic Exopolysaccharide That Cross-Links Extracellular DNA in the Pseudomonas Aeruginosa Biofilm Matrix. Proc. Natl. Acad. Sci. USA 2015, 112, 11353–11358. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jennings, L.K.; Dreifus, J.E.; Reichhardt, C.; Storek, K.M.; Secor, P.R.; Wozniak, D.J.; Hisert, K.B.; Parsek, M.R. Pseudomonas Aeruginosa Aggregates in Cystic Fibrosis Sputum Produce Exopolysaccharides That Likely Impede Current Therapies. Cell Rep. 2021, 34, 108782. [Google Scholar] [CrossRef] [PubMed]
- Hay, I.D.; Gatland, K.; Campisano, A.; Jordens, J.Z.; Rehm, B.H.A. Impact of Alginate Overproduction on Attachment and Biofilm Architecture of a Supermucoid Pseudomonas Aeruginosa Strain. Appl. Environ. Microbiol. 2009, 75, 6022–6025. [Google Scholar] [CrossRef] [Green Version]
- Costerton, J.W.; Stewart, P.S.; Greenberg, E.P. Bacterial Biofilms: A Common Cause of Persistent Infections. Science 1999, 284, 1318–1322. [Google Scholar] [CrossRef] [Green Version]
- Alhede, M.M.; Bjarnsholt, T.; Givskov, M.; Alhede, M.M. Pseudomonas Aeruginosa Biofilms. Mechanisms of Immune Evasion. Adv. Appl. Microbiol. 2014, 86, 1–40. [Google Scholar] [CrossRef]
- Le, K.Y.; Park, M.D.; Otto, M. Immune Evasion Mechanisms of Staphylococcus Epidermidis Biofilm Infection. Front. Microbiol. 2018, 9, 359. [Google Scholar] [CrossRef]
- Kirketerp-Møller, K.; Jensen, P.Ø.; Fazli, M.; Madsen, K.G.; Pedersen, J.; Moser, C.; Tolker-Nielsen, T.; Høiby, N.; Givskov, M.; Bjarnsholt, T. Distribution, Organization, and Ecology of Bacteria in Chronic Wounds. J. Clin. Microbiol. 2008, 46, 2717–2722. [Google Scholar] [CrossRef] [Green Version]
- Donlan, R.M. Preventing Biofilms of Clinically Relevant Organisms Using Bacteriophage. Trends Microbiol. 2009, 17, 66–72. [Google Scholar] [CrossRef]
- Abedon, S. Ecology of Anti-Biofilm Agents I: Antibiotics versus Bacteriophages. Pharmaceuticals 2015, 8, 525–558. [Google Scholar] [CrossRef] [PubMed]
- Espeland, E.M.; Wetzel, R.G. Complexation, Stabilization, and UV Photolysis of Extracellular and Surface-Bound Glucosidase and Alkaline Phosphatase: Implications for Biofilm Microbiota. Microb. Ecol. 2001, 42, 572–585. [Google Scholar] [CrossRef] [PubMed]
- Le Magrex-Debar, E.; Lemoine, J.; Gellé, M.-P.; Jacquelin, L.-F.; Choisy, C. Evaluation of Biohazards in Dehydrated Biofilms on Foodstuff Packaging. Int. J. Food Microbiol. 2000, 55, 239–243. [Google Scholar] [CrossRef]
- Leid, J.G.; Shirtliff, M.E.; Costerton, J.W.; Stoodley, P. Human Leukocytes Adhere to, Penetrate, and Respond to Staphylococcus Aureus Biofilms. Infect. Immun. 2002, 70, 6339–6345. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McNeill, K.; Hamilton, I.R. Acid Tolerance Response of Biofilm Cells of Streptococcus mutans. FEMS Microbiol. Lett. 2003, 221, 25–30. [Google Scholar] [CrossRef] [Green Version]
- Teitzel, G.M.; Parsek, M.R. Heavy Metal Resistance of Biofilm and Planktonic Pseudomonas Aeruginosa. Appl. Environ. Microbiol. 2003, 69, 2313–2320. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Soler-Arango, J.; Figoli, C.; Muraca, G.; Bosch, A.; Brelles-Mariño, G. The Pseudomonas Aeruginosa Biofilm Matrix and Cells Are Drastically Impacted by Gas Discharge Plasma Treatment: A Comprehensive Model Explaining Plasma-Mediated Biofilm Eradication. PLoS ONE 2019, 14, 0216817. [Google Scholar] [CrossRef] [Green Version]
- Matz, C. Biofilms as Refuge against Predation. Lab. Chip 2009, 10, 238. [Google Scholar]
- Lebeaux, D.; Ghigo, J.-M.; Beloin, C. Biofilm-Related Infections: Bridging the Gap between Clinical Management and Fundamental Aspects of Recalcitrance toward Antibiotics. Microbiol. Mol. Biol. Rev. 2014, 78, 510–543. [Google Scholar] [CrossRef] [Green Version]
- Millman, A.; Bernheim, A.; Stokar-Avihail, A.; Fedorenko, T.; Voichek, M.; Leavitt, A.; Oppenheimer-Shaanan, Y.; Sorek, R. Bacterial Retrons Function in Anti-Phage Defense. Cell 2020, 183, 1551–1561. [Google Scholar] [CrossRef]
- Labrie, S.J.; Samson, J.E.; Moineau, S. Bacteriophage Resistance Mechanisms. Nat. Rev. Microbiol. 2010, 8, 317–327. [Google Scholar] [CrossRef] [PubMed]
- Song, S.; Wood, T.K. Post-Segregational Killing and Phage Inhibition Are Not Mediated by Cell Death Through Toxin/Antitoxin Systems. Front. Microbiol. 2018, 9, 814. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lacqua, A.; Wanner, O.; Colangelo, T.; Martinotti, M.G.; Landini, P. Emergence of Biofilm-Forming Subpopulations upon Exposure of Escherichia Coli to Environmental Bacteriophages. Appl. Environ. Microbiol. 2006, 72, 956–959. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seed, K.D. Battling Phages: How Bacteria Defend against Viral Attack. PLoS Pathog. 2015, 11, e1004847. [Google Scholar] [CrossRef] [Green Version]
- Hughes, K.A.; Sutherland, I.W.; Jones, M. v Biofilm Susceptibility to Bacteriophage Attack: The Role of Phage-Borne Polysaccharide Depolymerase. Microbiology 1998, 144, 3039–3047. [Google Scholar] [CrossRef] [Green Version]
- Pires, D.P.; Dötsch, A.; Anderson, E.M.; Hao, Y.; Khursigara, C.M.; Lam, J.S.; Sillankorva, S.; Azeredo, J. A Genotypic Analysis of Five P. Aeruginosa Strains after Biofilm Infection by Phages Targeting Different Cell Surface Receptors. Front. Microbiol. 2017, 8, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Reyes-Robles, T.; Dillard, R.S.; Cairns, L.S.; Silva-Valenzuela, C.A.; Housman, M.; Ali, A.; Wright, E.R. Vibrio Cholerae Outer Membrane Vesicles Inhibit Bacteriophage Infection. J. Bacteriol. 2018, 200, e00792-17. [Google Scholar] [CrossRef] [Green Version]
- Azeredo, J.; Sutherland, I.W. The Use of Phages for the Removal of Infectious Biofilms. Curr. Pharm. Biotechnol. 2014, 9, 261–266. [Google Scholar] [CrossRef]
- Hu, J.; Miyanaga, K.; Tanji, Y. Diffusion of Bacteriophages through Artificial Biofilm Models. Biotechnol. Prog. 2012, 28, 319–326. [Google Scholar] [CrossRef]
- Henriksen, K.; Rørbo, N.; Rybtke, M.L.; Martinet, M.G.; Tolker-Nielsen, T.; Høiby, N.; Middelboe, M.; Ciofu, O.P. Aeruginosa Flow-Cell Biofilms Are Enhanced by Repeated Phage Treatments but Can Be Eradicated by Phage–Ciprofloxacin Combination. Pathog. Dis. 2019, 77, ftz011. [Google Scholar] [CrossRef]
- Gödeke, J.; Paul, K.; Lassak, J.; Thormann, K.M. Phage-Induced Lysis Enhances Biofilm Formation in Shewanella Oneidensis MR-1. ISME J. 2011, 5, 613–626. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Heilmann, S.; Sneppen, K.; Krishna, S. Coexistence of Phage and Bacteria on the Boundary of Self-Organized Refuges. Proc. Natl. Acad. Sci. USA 2012, 109, 12828–12833. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schrag, S.J.; Mittler, J.E. Host-Parasite Coexistence: The Role of Spatial Refuges in Stabilizing Bacteria-Phage Interactions. Am. Nat. 1996, 148, 348–377. [Google Scholar] [CrossRef]
- Eriksen, R.S.; Svenningsen, S.L.; Sneppen, K.; Mitarai, N. A Growing Microcolony Can Survive and Support Persistent Propagation of Virulent Phages. Proc. Natl. Acad. Sci. USA 2017, 115, 337–342. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Altindis, E.; Fu, Y.; Mekalanos, J.J. Proteomic Analysis of Vibrio Cholerae Outer Membrane Vesicles. Proc. Natl. Acad. Sci. USA 2014, 111, E1548–E1556. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stewart, P.S. Diffusion in Biofilms. J. Bacteriol. 2003, 185, 1485–1491. [Google Scholar] [CrossRef] [Green Version]
- Ghanem, N.; Kiesel, B.; Kallies, R.; Harms, H.; Chatzinotas, A.; Wick, L.Y. Marine Phages As Tracers: Effects of Size, Morphology, and Physico–Chemical Surface Properties on Transport in a Porous Medium. Environ. Sci. Technol. 2016, 50, 12816–12824. [Google Scholar] [CrossRef]
- Marcotte, L.; Therien-Aubin, H.; Sandt, C.; Barbeau, J.; Lafleur, M. Biofouling The Journal of Bioadhesion and Biofilm Research Solute Size Effects on the Diffusion in Biofilms of Streptococcus Mutans. Biofouling 2004, 20, 189–201. [Google Scholar] [CrossRef]
- Gallet, R.; Shao, Y.; Wang, I.-N. High Adsorption Rate Is Detrimental to Bacteriophage Fitness in a Biofilm-like Environment. BMC Evol. Biol. 2009, 9, 241. [Google Scholar] [CrossRef] [Green Version]
- Cornelissen, A.; Ceyssens, P.-J.J.; T’Syen, J.; Van Praet, H.; Noben, J.-P.P.; Shaburova, O.V.; Krylov, V.N.; Volckaert, G.; Lavigne, R. The T7-Related Pseudomonas Putida Phage Varphi15 Displays Virion-Associated Biofilm Degradation Properties. PLoS ONE 2011, 6, e18597. [Google Scholar] [CrossRef] [Green Version]
- Gutiérrez, D.; Rodríguez-Rubio, L.; Martínez, B.; Rodríguez, A.; García, P. Bacteriophages as Weapons against Bacterial Biofilms in the Food Industry. Front. Microbiol. 2016, 7, 825. [Google Scholar] [CrossRef] [PubMed]
- Von Ohle, C.; Gieseke, A.; Nistico, L.; Decker, E.M.; Debeer, D.; Stoodley, P. Real-Time Microsensor Measurement of Local Metabolic Activities in Ex Vivo Dental Biofilms Exposed to Sucrose and Treated with Chlorhexidine. Appl. Environ. Microbiol. 2010, 76, 2326–2334. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stewart, P.S.; Zhang, T.; Xu, R.; Pitts, B.; Walters, M.C.; Roe, F.; Kikhney, J.; Moter, A. Reaction-Diffusion Theory Explains Hypoxia and Heterogeneous Growth within Microbial Biofilms Associated with Chronic Infections. NPJ Biofilms Microb. 2016, 2, 16012. [Google Scholar] [CrossRef] [PubMed]
- Sillankorva, S.; Oliveira, R.; Vieira, M.J.; Sutherland, I.W.; Azeredo, J. Bacteriophage Φ S1 Infection of Pseudomonas Fluorescens Planktonic Cells versus Biofilms. Biofouling 2004, 20, 133–138. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fraser, D.; Kærn, M. A Chance at Survival: Gene Expression Noise and Phenotypic Diversification Strategies. Mol. Microbiol. 2009, 71, 1333–1340. [Google Scholar] [CrossRef] [PubMed]
- Verstraeten, N.; Knapen, W.; Fauvart, M.; Michiels, J. A Historical Perspective on Bacterial Persistence. In Methods in Molecular Biology; Springer: Berlin/Heidelberg, Germany, 2016; Volume 1333, pp. 3–13. ISBN 9781493928545. [Google Scholar]
- Sutherland, I.W.; Hughes, K.A.; Skillman, L.C.; Tait, K. The Interaction of Phage and Biofilms. FEMS Microbiol. Lett. 2004, 232, 1–6. [Google Scholar] [CrossRef]
- Hadas, H.; Einav, M.; Fishov, I.; Zaritsky, A. Bacteriophage T4 Development Depends on the Physiology of Its Host Escherichia Coli. Microbiology 1997, 143, 179–185. [Google Scholar] [CrossRef] [Green Version]
- Parasion, S.; Kwiatek, M.; Gryko, R.; Mizak, L.; Malm, A. Bacteriophages as an Alternative Strategy for Fighting Biofilm Development. Pol. J. Microbiol. 2014, 63, 137–145. [Google Scholar] [CrossRef]
- Cerca, N.; Oliveira, R.; Azeredo, J. Susceptibility of Staphylococcus Epidermidis Planktonic Cells and Biofilms to the Lytic Action of Staphylococcus Bacteriophage K. Lett. Appl. Microbiol. 2007, 45, 313–317. [Google Scholar] [CrossRef] [Green Version]
- Jamal, M.; Hussain, T.; Rajanna Das, C.; Andleeb, S. Characterization of Siphoviridae Phage Z and Studying Its Efficacy against Multidrug-Resistant Klebsiella Pneumoniae Planktonic Cells and Biofilm. J. Med. Microbiol. 2015, 64, 454–462. [Google Scholar] [CrossRef] [Green Version]
- Hanlon, G.W.; Denyer, S.P.; Olliff, C.J.; Ibrahim, L.J. Reduction in Exopolysaccharide Viscosity as an Aid to Bacteriophage Penetration through Pseudomonas Aeruginosa Biofilms. Appl. Environ. Microbiol. 2001, 67, 2746–2753. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hosseinidoust, Z.; Tufenkji, N.; van de Ven, T.G.M. Formation of Biofilms under Phage Predation: Considerations Concerning a Biofilm Increase. Biofouling 2013, 29, 457–468. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chan, T.; Lee, K.; Roulin, P. Escherichia Coli Biofilm Development and Bacteriophage T4 Susceptibility. J. Exp. Microbiol. Immunol. 2007, 11, 73–78. [Google Scholar]
- Sillankorva, S.; Neubauer, P.; Azeredo, J. Pseudomonas Fluorescens Biofilms Subjected to Phage PhiIBB-PF7A. BMC Biotechnol. 2008, 8, 79. [Google Scholar] [CrossRef] [Green Version]
- Tkhilaishvili, T.; Lombardi, L.; Klatt, A.-B.; Trampuz, A.; Di Luca, M. Bacteriophage Sb-1 Enhances Antibiotic Activity against Biofilm, Degrades Exopolysaccharide Matrix and Targets Persisters of Staphylococcus Aureus. Int. J. Antimicrob. Agents 2018, 52, 842–853. [Google Scholar] [CrossRef]
- Pearl, S.; Gabay, C.; Kishony, R.; Oppenheim, A.; Balaban, N.Q. Nongenetic Individuality in the Host-Phage Interaction. PLoS Biol. 2008, 6, 120. [Google Scholar] [CrossRef] [Green Version]
- Bryan, D.; El-Shibiny, A.; Hobbs, Z.; Porter, J.; Kutter, E.M. Bacteriophage T4 Infection of Stationary Phase E. Coli: Life after Log from a Phage Perspective. Front. Microbiol. 2016, 7, 1391. [Google Scholar] [CrossRef] [Green Version]
- Rickard, A.H.; Gilbert, P.; High, N.J.; Kolenbrander, P.E.; Handley, P.S. Bacterial Coaggregation: An Integral Process in the Development of Multi-Species Biofilms. Trends Microbiol. 2003, 11, 94–100. [Google Scholar] [CrossRef]
- Vilain, S.; Pretorius, J.M.; Theron, J.; Brözel, V.S. DNA as an Adhesin: Bacillus Cereus Requires Extracellular DNA to Form Biofilms. Appl. Environ. Microbiol. 2009, 75, 2861–2868. [Google Scholar] [CrossRef] [Green Version]
- Tait, K.; Skillman, L.C.; Sutherland, I.W. The Efficacy of Bacteriophage as a Method of Biofilm Eradication. Biofouling 2002, 18, 305–311. [Google Scholar] [CrossRef] [Green Version]
- González, S.; Fernández, L.; Campelo, A.B.; Gutiérrez, D.; Martínez, B.; Rodríguez, A.; García, P. The Behavior of Staphylococcus Aureus Dual-Species Biofilms Treated with Bacteriophage PhiIPLA-RODI Depends on the Accompanying Microorganism. Appl. Environ. Microbiol. 2017, 83, e02821-16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Testa, S.; Berger, S.; Piccardi, P.; Oechslin, F.; Resch, G.; Mitri, S. Spatial Structure Affects Phage Efficacy in Infecting Dual-Strain Biofilms of Pseudomonas Aeruginosa. Commun. Biol. 2019, 2, 405. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chao, L.; Ramsdell, G. The Effects of Wall Populations on Coexistence of Bacteria in the Liquid Phase of Chemostat Cultures. J. Gen. Microbiol. 1985, 131, 1229–1236. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weiss, M.; Denou, E.; Bruttin, A.; Serra-Moreno, R.; Dillmann, M.L.; Brüssow, H. In Vivo Replication of T4 and T7 Bacteriophages in Germ-Free Mice Colonized with Escherichia Coli. Virology 2009, 393, 16–23. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Darch, S.E.; Kragh, K.N.; Abbott, E.A.; Bjarnsholt, T.; Bull, J.J.; Whiteley, M. Phage Inhibit Pathogen Dissemination by Targeting Bacterial Migrants in a Chronic Infection Model. mBio 2017, 8, e00240-17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boles, B.R.; Singh, P.K.; Welsh, M.J. Endogenous Oxidative Stress Produces Diversity and Adaptability in Biofilm Communities. Proc. Natl. Acad. Sci. USA 2008, 105, 12503–12508. [Google Scholar] [CrossRef] [Green Version]
- Kivisaar, M. Mechanisms of Stationary-Phase Mutagenesis in Bacteria: Mutational Processes in Pseudomonads. FEMS Microbiol. Lett. 2010, 312, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Bjedov, I.; Tenaillon, O.; Gérard, B.; Souza, V.; Denamur, E.; Radman, M.; Taddei, F.; Matic, I. Stress-Induced Mutagenesis in Bacteria. Science 2003, 300, 1404–1409. [Google Scholar] [CrossRef]
- Madsen, J.S.; Burmølle, M.; Hansen, L.H.; Sørensen, S.J. The Interconnection between Biofilm Formation and Horizontal Gene Transfer. FEMS Immunol. Med. Microbiol. 2012, 65, 183–195. [Google Scholar] [CrossRef] [Green Version]
- Hendrickx, L.; Hausner, M.; Wuertz, S. Natural Genetic Transformation in Monoculture Acinetobacter Sp. Strain BD413 Biofilms. Appl. Environ. Microbiol. 2003, 69, 1721–1727. [Google Scholar] [CrossRef] [Green Version]
- Maeda, S.; Ito, M.; Ando, T.; Ishimoto, Y.; Fujisawa, Y.; Takahashi, H.; Matsuda, A.; Sawamura, A.; Kato, S. Horizontal Transfer of Nonconjugative Plasmids in a Colony Biofilm of Escherichia coli. FEMS Microbiol. Lett. 2006, 255, 115–120. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Etchuuya, R.; Ito, M.; Kitano, S.; Shigi, F.; Sobue, R.; Maeda, S. Cell-to-Cell Transformation in Escherichia Coli: A Novel Type of Natural Transformation Involving Cell-Derived DNA and a Putative Promoting Pheromone. PLoS ONE 2011, 6, e16355. [Google Scholar] [CrossRef]
- Ghigo, J.-M. Natural Conjugative Plasmids Induce Bacterial Biofilm Development. Nature 2001, 412, 442–445. [Google Scholar] [CrossRef] [PubMed]
- Nobrega, F.L.; Costa, A.R.; Kluskens, L.D.; Azeredo, J. Revisiting Phage Therapy: New Applications for Old Resources. Trends Microbiol. 2015, 23, 185–191. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lopez, C.A.; Winter, S.E.; Rivera-Chávez, F.; Xavier, M.N.; Poon, V.; Nuccio, S.-P.; Tsolis, R.M.; Bäumler, A.J. Phage-Mediated Acquisition of a Type III Secreted Effector Protein Boosts Growth of Salmonella by Nitrate Respiration. mBio 2012, 3, e00143-12. [Google Scholar] [CrossRef] [Green Version]
- Muhly, T.B.; Semeniuk, C.; Massolo, A.; Hickman, L.; Musiani, M. Human Activity Helps Prey Win the Predator-Prey Space Race. PLoS ONE 2011, 6, e17050. [Google Scholar] [CrossRef]
- Luttbeg, B.; Fraker, M.E. Predator-Prey Space Use and the Spatial Distribution of Predation Events. Behaviour 2012, 149, 555–574. [Google Scholar] [CrossRef] [Green Version]
- Abedon, S. Ecology of Anti-Biofilm Agents II: Bacteriophage Exploitation and Biocontrol of Biofilm Bacteria. Pharmaceuticals 2015, 8, 559–589. [Google Scholar] [CrossRef] [Green Version]
- Kay, M.K.; Erwin, T.C.; McLean, R.J.C.; Aron, G.M. Bacteriophage Ecology in Escherichia Coli and Pseudomonas Aeruginosa Mixed-Biofilm Communities. Appl. Environ. Microbiol. 2011, 77, 821–829. [Google Scholar] [CrossRef] [Green Version]
- Ali, Q. Non-Conventional Therapeutic Technique to Replace CRISPR Bacteria from Biofilm by Inducible Lysogen. J. Biol. Dyn. 2019, 13, 151–178. [Google Scholar] [CrossRef] [Green Version]
- Blokesch, M.; Schoolnik, G.K. Serogroup Conversion of Vibrio Cholerae in Aquatic Reservoirs. PLoS Pathog. 2007, 3, e81. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Kim, Y.; Ma, Q.; Hong, S.H.; Pokusaeva, K.; Sturino, J.M.; Wood, T.K. Cryptic Prophages Help Bacteria Cope with Adverse Environments. Nat. Commun. 2010, 1, 147. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Toba, F.A.; Thompson, M.G.; Campbell, B.R.; Junker, L.M.; Rueggeberg, K.-G.; Hay, A.G. Role of DLP12 Lysis Genes in Escherichia Coli Biofilm Formation. Microbiology 2011, 157, 1640–1650. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yamane, T.; Enokida, H.; Hayami, H.; Kawahara, M.; Nakagawa, M. Genome-Wide Transcriptome Analysis of Fluoroquinolone Resistance in Clinical Isolates of Escherichia Coli. Int. J. Urol. 2012, 19, 360–368. [Google Scholar] [CrossRef] [PubMed]
- Müller, M.G.; Ing, J.Y.; Cheng, M.K.-W.; Flitter, B.A.; Moe, G.R. Identification of a Phage-Encoded Ig-Binding Protein from Invasive Neisseria Meningitidis. J. Immunol. 2013, 191, 3287–3296. [Google Scholar] [CrossRef] [Green Version]
- Secor, P.R.; Jennings, L.K.; Michaels, L.A.; Sweere, J.M.; Singh, P.K.; Parks, W.C.; Bollyky, P.L. Biofilm Assembly Becomes Crystal Clear—Filamentous Bacteriophage Organize the Pseudomonas Aeruginosa Biofilm Matrix into a Liquid Crystal. Microb. Cell 2016, 3, 49–52. [Google Scholar] [CrossRef]
- Martínez, E.; Campos-Gómez, J. Pf Filamentous Phage Requires UvrD for Replication in Pseudomonas Aeruginosa. mSphere 2016, 1, e00104-15. [Google Scholar] [CrossRef] [Green Version]
- Davies, E.V.; James, C.E.; Williams, D.; O’Brien, S.; Fothergill, J.L.; Haldenby, S.; Paterson, S.; Winstanley, C.; Brockhurst, M.A. Temperate Phages Both Mediate and Drive Adaptive Evolution in Pathogen Biofilms. Proc. Natl. Acad. Sci. USA 2016, 113, 8266–8271. [Google Scholar] [CrossRef] [Green Version]
- Martin, M.; Dragoš, A.; Hölscher, T.; Maróti, G.; Bálint, B.; Westermann, M.; Kovács, Á.T. De Novo Evolved Interference Competition Promotes the Spread of Biofilm Defectors. Nat. Commun. 2017, 8, 15127. [Google Scholar] [CrossRef] [PubMed]
- Shidore, T.; Triplett, L.R. Toxin-Antitoxin Systems: Implications for Plant Disease. Annu. Rev. Phytopathol. 2017, 55, 161–179. [Google Scholar] [CrossRef]
- Rice, S.A.; Tan, C.H.; Mikkelsen, P.J.; Kung, V.; Woo, J.; Tay, M.; Hauser, A.; McDougald, D.; Webb, J.S.; Kjelleberg, S. The Biofilm Life Cycle and Virulence of Pseudomonas Aeruginosa Are Dependent on a Filamentous Prophage. ISME J. 2009, 3, 271–282. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fu, Q.; Li, S.; Wang, Z.; Shan, W.; Ma, J.; Cheng, Y.; Wang, H.; Yan, Y.; Sun, J. H-NS Mutation-Mediated CRISPR-Cas Activation Inhibits Phage Release and Toxin Production of Escherichia Coli Stx2 Phage Lysogen. Front. Microbiol. 2017, 8, 652. [Google Scholar] [CrossRef] [PubMed]
- Nanda, A.M.; Thormann, K.; Frunzke, J. Impact of Spontaneous Prophage Induction on the Fitness of Bacterial Populations and Host-Microbe Interactions. J. Bacteriol. 2015, 197, 410–419. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Obeng, N.; Pratama, A.A.; van Elsas, J.D. The Significance of Mutualistic Phages for Bacterial Ecology and Evolution. Trends Microbiol. 2016, 24, 440–449. [Google Scholar] [CrossRef] [PubMed]
- Hui, J.G.K.; Mai-Prochnow, A.; Kjelleberg, S.; McDougald, D.; Rice, S.A. Environmental Cues and Genes Involved in Establishment of the Superinfective Pf4 Phage of Pseudomonas Aeruginosa. Front. Microbiol. 2014, 5, 654. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Whiteley, M.; Bangera, M.G.; Bumgarner, R.E.; Parsek, M.R.; Teitzel, G.M.; Lory, S.; Greenberg, E.P. Gene Expression in Pseudomonas Aeruginosa Biofilms. Nature 2001, 413, 860–864. [Google Scholar] [CrossRef]
- Couto, N.; Belas, A.; Oliveira, M.; Almeida, P.; Clemente, C.; Pomba, C. Comparative RNA-Seq-Based Transcriptome Analysis of the Virulence Characteristics of Methicillin-Resistant and -Susceptible Staphylococcus Pseudintermedius Strains Isolated from Small Animals. Antimicrob. Agents Chemother. 2016, 60, 962–967. [Google Scholar] [CrossRef] [Green Version]
- García-Contreras, R.; Zhang, X.S.; Kim, Y.; Wood, T.K. Protein Translation and Cell Death: The Role of Rare TRNAs in Biofilm Formation and in Activating Dormant Phage Killer Genes. PLoS ONE 2008, 3, e0002394. [Google Scholar] [CrossRef]
- Mitchell, H.L.; Dashper, S.G.; Catmull, D.V.; Paolini, R.A.; Cleal, S.M.; Slakeski, N.; Tan, K.H.; Reynolds, E.C. Treponema Denticola Biofilm-Induced Expression of a Bacteriophage, Toxin-Antitoxin Systems and Transposases. Microbiology 2010, 156, 774–788. [Google Scholar] [CrossRef] [Green Version]
- Sauer, K.; Cullen, M.C.; Rickard, A.H.; Zeef, L.A.H.; Davies, D.G.; Gilbert, P. Characterization of Nutrient-Induced Dispersion in Pseudomonas Aeruginosa PAO1 Biofilm. J. Bacteriol. 2004, 186, 7312–7326. [Google Scholar] [CrossRef] [Green Version]
- Webb, J.S.; Lau, M.; Kjelleberg, S. Bacteriophage and Phenotypic Variation in Pseudomonas Aeruginosa Biofilm Development. J. Bacteriol. 2004, 186, 8066–8073. [Google Scholar] [CrossRef] [Green Version]
- Schuch, R.; Fischetti, V.A. The Secret Life of the Anthrax Agent Bacillus Anthracis: Bacteriophage-Mediated Ecological Adaptations. PLoS ONE 2009, 4, e6532. [Google Scholar] [CrossRef] [Green Version]
- Secor, P.R.; Sweere, J.M.; Michaels, L.A.; Malkovskiy, A.V.; Lazzareschi, D.; Katznelson, E.; Rajadas, J.; Birnbaum, M.E.; Arrigoni, A.; Braun, K.R.; et al. Filamentous Bacteriophage Promote Biofilm Assembly and Function. Cell Host Microbe 2015, 18, 549–559. [Google Scholar] [CrossRef] [Green Version]
- Binnenkade, L.; Teichmann, L.; Thormann, K.M. Iron Triggers ΛSo Prophage Induction and Release of Extracellular DNA in Shewanella Oneidensis MR-1 Biofilms. Appl. Environ. Microbiol. 2014, 80, 5304–5316. [Google Scholar] [CrossRef] [Green Version]
- Resch, A.; Fehrenbacher, B.; Eisele, K.; Schaller, M.; Götz, F. Phage Release from Biofilm and Planktonic Staphylococcus Aureus Cells. FEMS Microbiol. Lett. 2005, 252, 89–96. [Google Scholar] [CrossRef] [Green Version]
- Webb, J.S.; Thompson, L.S.; James, S.; Charlton, T.; Tolker-Nielsen, T.; Koch, B.; Givskov, M.; Kjelleberg, S. Cell Death in Pseudomonas Aeruginosa Biofilm Development. J. Bacteriol. 2003, 185, 4585–4592. [Google Scholar] [CrossRef] [Green Version]
- Rossmann, F.S.; Racek, T.; Wobser, D.; Puchalka, J.; Rabener, E.M.; Reiger, M.; Hendrickx, A.P.A.; Diederich, A.-K.; Jung, K.; Klein, C.; et al. Phage-Mediated Dispersal of Biofilm and Distribution of Bacterial Virulence Genes Is Induced by Quorum Sensing. PLoS Pathog. 2015, 11, e1004653. [Google Scholar] [CrossRef]
- Kirov, S.M.; Webb, J.S.; O’May, C.Y.; Reid, D.W.; Woo, J.K.K.; Rice, S.A.; Kjelleberg, S. Biofilm Differentiation and Dispersal in Mucoid Pseudomonas Aeruginosa Isolates from Patients with Cystic Fibrosis. Microbiology 2007, 153, 3264–3274. [Google Scholar] [CrossRef] [Green Version]
- D’Argenio, D.A.; Calfee, M.W.; Rainey, P.B.; Pesci, E.C. Autolysis and Autoaggregation in Pseudomonas Aeruginosa Colony Morphology Mutants. J. Bacteriol. 2002, 184, 6481–6489. [Google Scholar] [CrossRef] [Green Version]
- Fernández-Piñar, R.; Cámara, M.; Dubern, J.-F.; Ramos, J.L.; Espinosa-Urgel, M. The Pseudomonas Aeruginosa Quinolone Quorum Sensing Signal Alters the Multicellular Behaviour of Pseudomonas Putida KT2440. Res. Microbiol. 2011, 162, 773–781. [Google Scholar] [CrossRef]
- Muranaka, L.S.; Takita, M.A.; Olivato, J.C.; Kishi, L.T.; de Souza, A.A. Global Expression Profile of Biofilm Resistance to Antimicrobial Compounds in the Plant-Pathogenic Bacterium Xylella Fastidiosa Reveals Evidence of Persister Cells. J. Bacteriol. 2012, 194, 4561–4569. [Google Scholar] [CrossRef]
- Moons, P.; Faster, D.; Aertsen, A. Lysogenic Conversion and Phage Resistance Development in Phage Exposed Escherichia Coli Biofilms. Viruses 2013, 5, 150–161. [Google Scholar] [CrossRef] [Green Version]
- Hazan, R.; Que, Y.A.; Maura, D.; Strobel, B.; Majcherczyk, P.A.; Hopper, L.R.; Wilbur, D.J.; Hreha, T.N.; Barquera, B.; Rahme, L.G. Auto Poisoning of the Respiratory Chain by a Quorum-Sensing-Regulated Molecule Favors Biofilm Formation and Antibiotic Tolerance. Curr. Biol. 2016, 26, 195–206. [Google Scholar] [CrossRef] [Green Version]
- Guiton, P.S.; Hung, C.S.; Kline, K.A.; Roth, R.; Kau, A.L.; Hayes, E.; Heuser, J.; Dodson, K.W.; Caparon, M.G.; Hultgren, S.J. Contribution of Autolysin and Sortase a during Enterococcus Faecalis DNA-Dependent Biofilm Development. Infect. Immun. 2009, 77, 3626–3638. [Google Scholar] [CrossRef] [Green Version]
- Jin, H.; Squier, T.C.; Long, P.E. Dying for Good: Virus-Bacterium Biofilm Co-Evolution Enhances Environmental Fitness. Biochem. Insights 2012, 5, BCI.S9553. [Google Scholar] [CrossRef]
- Carrolo, M.; Frias, M.J.; Pinto, F.R.; Melo-Cristino, J.; Ramirez, M. Prophage Spontaneous Activation Promotes DNA Release Enhancing Biofilm Formation in Streptococcus Pneumoniae. PLoS ONE 2010, 5, e15678. [Google Scholar] [CrossRef] [Green Version]
- Mulcahy, H.; Charron-mazenod, L.; Lewenza, S. Extracellular DNA Chelates Cations and Induces Antibiotic Resistance in Pseudomonas Aeruginosa Biofilms. PLoS Pathog. 2008, 4, e1000213. [Google Scholar] [CrossRef] [Green Version]
- Leigh, B.; Karrer, C.; Cannon, J.P.; Breitbart, M.; Dishaw, L.J. Isolation and Characterization of a Shewanella Phage-Host System from the Gut of the Tunicate, Ciona Intestinalis. Viruses 2017, 9, 60. [Google Scholar] [CrossRef]
- Proctor, R.A.; von Eiff, C.; Kahl, B.C.; Becker, K.; McNamara, P.; Herrmann, M.; Peters, G. Small Colony Variants: A Pathogenic Form of Bacteria That Facilitates Persistent and Recurrent Infections. Nat. Rev. Microbiol. 2006, 4, 295–305. [Google Scholar] [CrossRef]
- Whitchurch, C.B.; Tolker-nielsen, T.; Ragas, P.C.; Mattick, J.S.; Mattickl, J.S. Extracellular DNA Required for Bacterial Biofilm Formation. Science 2002, 295, 1487. [Google Scholar] [CrossRef]
- Fernández, L.; González, S.; Quiles-Puchalt, N.; Gutiérrez, D.; Penadés, J.R.; García, P.; Rodríguez, A. Lysogenization of Staphylococcus Aureus RN450 by Phages Φ11 and Φ80α Leads to the Activation of the SigB Regulon. Sci. Rep. 2018, 8, 12662. [Google Scholar] [CrossRef]
- Uhlich, G.A.; Chen, C.-Y.; Cottrell, B.J.; Hofmann, C.S.; Dudley, E.G.; Strobaugh, T.P.; Nguyen, L.-H. Phage Insertion in MlrA and Variations in RpoS Limit Curli Expression and Biofilm Formation in Escherichia Coli Serotype O157:H7. Microbiology 2013, 159, 1586–1596. [Google Scholar] [CrossRef] [Green Version]
- Tikhomirova, A.; Trappetti, C.; Paton, J.C.; Kidd, S.P. The Outcome of H. Influenzae and S. Pneumoniae Inter-Species Interactions Depends on PH, Nutrient Availability and Growth Phase. Int. J. Med. Microbiol. 2015, 305, 881–892. [Google Scholar] [CrossRef]
- Ripp, S.; Miller, R.V. The Role of Pseudolysogeny in Bacteriophage-Host Interactions in a Natural Freshwater Environment. Microbiology 1997, 143, 2065–2070. [Google Scholar] [CrossRef] [Green Version]
- Weinbauer, M.G. Ecology of Prokaryotic Viruses. FEMS Microbiol. Rev. 2004, 28, 127–181. [Google Scholar] [CrossRef] [Green Version]
- Chan, B.; Abedon, S. Bacteriophages and Their Enzymes in Biofilm Control. Curr. Pharm. Des. 2014, 21, 85–99. [Google Scholar] [CrossRef]
- Vidakovic, L.; Singh, P.K.; Hartmann, R.; Nadell, C.D.; Drescher, K. Dynamic Biofilm Architecture Confers Individual and Collective Mechanisms of Viral Protection. Nat. Microbiol. 2018, 3, 26–31. [Google Scholar] [CrossRef]
- Lu, T.K.; Collins, J.J. Dispersing Biofilms with Engineered Enzymatic Bacteriophage. Proc. Natl. Acad. Sci. USA 2007, 104, 11197–11202. [Google Scholar] [CrossRef] [Green Version]
- Nale, J.Y.; Chutia, M.; Carr, P.; Hickenbotham, P.T.; Clokie, M.R.J. “Get in Early”; Biofilm and Wax Moth (Galleria Mellonella) Models Reveal New Insights into the Therapeutic Potential of Clostridium Difficile Bacteriophages. Front. Microbiol. 2016, 7, 1383. [Google Scholar] [CrossRef] [Green Version]
- Glonti, T.; Chanishvili, N.; Taylor, P.W. Bacteriophage-Derived Enzyme That Depolymerizes the Alginic Acid Capsule Associated with Cystic Fibrosis Isolates of Pseudomonas Aeruginosa. J. Appl. Microbiol. 2010, 108, 695–702. [Google Scholar] [CrossRef]
- Mushtaq, N.; Redpath, M.B.; Luzio, J.P.; Taylor, P.W. Treatment of Experimental Escherichia Coli Infection with Recombinant Bacteriophage-Derived Capsule Depolymerase. J. Antimicrob. Chemother. 2005, 56, 160–165. [Google Scholar] [CrossRef] [Green Version]
- Motlagh, A.M.; Bhattacharjee, A.S.; Goel, R. Biofilm Control with Natural and Genetically-Modified Phages. World J. Microbiol. Biotechnol. 2016, 32, 67. [Google Scholar] [CrossRef]
- Pires, D.P.; Oliveira, H.; Melo, L.D.R.; Sillankorva, S.; Azeredo, J. Bacteriophage-Encoded Depolymerases: Their Diversity and Biotechnological Applications. Appl. Microbiol. Biotechnol. 2016, 100, 2141–2151. [Google Scholar] [CrossRef] [Green Version]
- Harper, D.; Parracho, H.; Walker, J.; Sharp, R.; Hughes, G.; Werthén, M.; Lehman, S.; Morales, S. Bacteriophages and Biofilms. Antibiotics 2014, 3, 270–284. [Google Scholar] [CrossRef]
- Al-Wrafy, F.; Brzozowska, E.; Górska, S.; Gamian, A. Pathogenic Factors of Pseudomonas Aeruginosa—The Role of Biofilm in Pathogenicity and as a Target for Phage Therapy. Postepy Hig. Med. Dosw. 2017, 71, 78–91. [Google Scholar] [CrossRef]
- Lelchat, F.; Mocaer, P.Y.; Ojima, T.; Michel, G.; Sarthou, G.; Bucciarelli, E.; Cérantola, S.; Colliec-Jouault, S.; Boisset, C.; Baudoux, A.-C. Viral Degradation of Marine Bacterial Exopolysaccharides. FEMS Microbiol. Ecol. 2019, 95, fiz079. [Google Scholar] [CrossRef]
- Sutherland, I.W. Polysaccharide Lyases. FEMS Microbiol. Rev. 1995, 16, 323–347. [Google Scholar] [CrossRef]
- Ding, H.; Yang, W.; Tang, H.; Feng, P.-M.; Huang, J.; Chen, W.; Lin, H. PHYPred: A Tool for Identifying Bacteriophage Enzymes and Hydrolases. Virol. Sin. 2016, 31, 350–352. [Google Scholar] [CrossRef]
- Alcántara, A.R.; Hernaiz, M.-J.; Sinisterra, J.-V. Biocatalyzed Production of Fine Chemicals. Compr. Biotechnol. 2011, 309–331. [Google Scholar] [CrossRef]
- Cheng, D.; Jiang, C.; Xu, J.; Liu, Z.; Mao, X. Characteristics and Applications of Alginate Lyases: A Review. Int. J. Biol. Macromol. 2020, 164, 1304–1320. [Google Scholar] [CrossRef]
- Zhu, B.; Yin, H. Alginate Lyase: Review of Major Sources and Classification, Properties, Structure-Function Analysis and Applications. Bioengineered 2015, 6, 125–131. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jakobsson, E.; Schwarzer, D.; Jokilammi, A.; Finne, J. Endosialidases: Versatile Tools for the Study of Polysialic Acid. Top. Curr. Chem. 2015, 367, 29–73. [Google Scholar] [CrossRef] [PubMed]
- Wollin, R.; Eriksson, U.; Lindberg, A.A. Salmonella Bacteriophage Glycanases: Endorhamnosidase Activity of Bacteriophages P27, 9NA, and KB1. J. Virol. 1981, 38, 1025–1033. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brzozowska, E.; Pyra, A.; Pawlik, K.; Janik, M.; Górska, S.; Urbańska, N.; Drulis-Kawa, Z.; Gamian, A. Hydrolytic Activity Determination of Tail Tubular Protein A of Klebsiella Pneumoniae Bacteriophages towards Saccharide Substrates. Sci. Rep. 2017, 7, 18048. [Google Scholar] [CrossRef] [Green Version]
- Paul, P.E.V.; Sangeetha, V.; Deepika, R.G. Emerging Trends in the Industrial Production of Chemical Products by Microorganisms. Recent Dev. Appl. Microbiol. Biochem. 2019, 107–125. [Google Scholar] [CrossRef]
- Bardal, S.K.; Waechter, J.E.; Martin, D.S.; Bardal, S.K.; Waechter, J.E.; Martin, D.S. Endocrinology. Appl. Pharmacol. 2011, 143–176. [Google Scholar] [CrossRef]
- Eun, H.-M.; Eun, H.-M. Marker/Reporter Enzymes. Enzymol. Primer Recomb. DNA Technol. 1996, 567–645. [Google Scholar] [CrossRef]
- Ramos, O.S.; Malcata, F.X. Food-Grade Enzymes. Compr. Biotechnol. 2011, 555–569. [Google Scholar] [CrossRef]
- Sun, A.; Chang, I.J.; Lam, C.; Berry, G.T. Lysosomal Storage Disorders. Emery Rimoin’s Princ. Pract. Med. Genet. Genom. 2021, 563–682. [Google Scholar] [CrossRef]
- Ito, M. Degradation of Glycolipids. Compr. Glycosci. 2007, 193–208. [Google Scholar] [CrossRef]
- Moracci, M.; Cobucci Ponzano, B.; Trincone, A.; Fusco, S.; De Rosa, M.; van Der Oost, J.; Sensen, C.W.; Charlebois, R.L.; Rossi, M. Identification and Molecular Characterization of the First Alpha -Xylosidase from an Archaeon. J. Biol. Chem. 2000, 275, 22082–22089. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Larsbrink, J.; Izumi, A.; Ibatullin, F.M.; Nakhai, A.; Gilbert, H.J.; Davies, G.J.; Brumer, H. Structural and Enzymatic Characterization of a Glycoside Hydrolase Family 31 α-Xylosidase from Cellvibrio Japonicus Involved in Xyloglucan Saccharification. Biochem. J. 2011, 436, 567–580. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jensen, S.L.; Diemer, M.B.; Lundmark, M.; Larsen, F.H.; Blennow, A.; Mogensen, H.K.; Nielsen, T.H. Levanase from Bacillus Subtilis Hydrolyses β-2,6 Fructosyl Bonds in Bacterial Levans and in Grass Fructans. Int. J. Biol. Macromol. 2016, 85, 514–521. [Google Scholar] [CrossRef] [PubMed]
- Hynes, W.L.; Walton, S.L. Hyaluronidases of Gram-Positive Bacteria. FEMS Microbiol. Lett. 2000, 183, 201–207. [Google Scholar] [CrossRef] [PubMed]
- Bello-Perez, L.A.; Hoyos-Leyva, J.D. Development of Foods High in Slowly Digestible and Resistant Starch. Starch Food 2018, 827–854. [Google Scholar] [CrossRef]
- Gutiérrez, D.; Martínez, B.; Rodríguez, A.; García, P. Genomic Characterization of Two Staphylococcus Epidermidis Bacteriophages with Anti-Biofilm Potential. BMC Genom. 2012, 13, 228. [Google Scholar] [CrossRef] [Green Version]
- Bartell, P.F.; Orr, T.E. Origin of Polysaccharide Depolymerase Associated with Bacteriophage Infection. J. Virol. 1969, 3, 290–296. [Google Scholar] [CrossRef] [Green Version]
- Schmerer, M.; Molineux, I.J.; Ally, D.; Tyerman, J.; Cecchini, N.; Bull, J.J. Challenges in Predicting the Evolutionary Maintenance of a Phage Transgene. J. Biol. Eng. 2014, 8, 21. [Google Scholar] [CrossRef] [Green Version]
- Fenton, M.; Keary, R.; McAuliffe, O.; Ross, R.P.; O’Mahony, J.; Coffey, A. Bacteriophage-Derived Peptidase CHAP(K) Eliminates and Prevents Staphylococcal Biofilms. Int. J. Microbiol. 2013, 2013, 625341. [Google Scholar] [CrossRef] [Green Version]
- Shen, Y.; Koller, T.; Kreikemeyer, B.; Nelson, D.C. Rapid Degradation of Streptococcus Pyogenes Biofilms by PlyC, a Bacteriophage-Encoded Endolysin. J. Antimicrob. Chemother. 2013, 68, 1818–1824. [Google Scholar] [CrossRef]
- Son, J.-S.; Lee, S.-J.; Jun, S.Y.; Yoon, S.J.; Kang, S.H.; Paik, H.R.; Kang, J.O.; Choi, Y.-J. Antibacterial and Biofilm Removal Activity of a Podoviridae Staphylococcus Aureus Bacteriophage SAP-2 and a Derived Recombinant Cell-Wall-Degrading Enzyme. Appl. Microbiol. Biotechnol. 2010, 86, 1439–1449. [Google Scholar] [CrossRef] [PubMed]
- Sass, P.; Bierbaum, G. Lytic Activity of Recombinant Bacteriophage Phi11 and Phi12 Endolysins on Whole Cells and Biofilms of Staphylococcus Aureus. Appl. Environ. Microbiol. 2007, 73, 347–352. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gutiérrez, D.; Ruas-Madiedo, P.; Martínez, B.; Rodríguez, A.; García, P. Effective Removal of Staphylococcal Biofilms by the Endolysin LysH5. PLoS ONE 2014, 9, e0107307. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moak, M.; Molineux, I.J. Peptidoglycan Hydrolytic Activities Associated with Bacteriophage Virions. Mol. Microbiol. 2004, 51, 1169–1183. [Google Scholar] [CrossRef]
- Duarte, V.S.; Dias, R.S.; Kropinski, A.M.; Vidigal, P.M.P.; Sousa, F.O.; Xavier, A.S.; Silva, C.C.; de Paula, S.O. Complete Genome Sequence of VB_EcoM-UFV13, a New Bacteriophage Able To Disrupt Trueperella Pyogenes Biofilm. Genome Announc. 2016, 4, e01292-16. [Google Scholar] [CrossRef] [Green Version]
- Ruhal, R.; Kataria, R. Biofilm Patterns in Gram-Positive and Gram-Negative Bacteria. Microbiol. Res. 2021, 251, 126829. [Google Scholar] [CrossRef]
- Williams, P.; Winzer, K.; Chan, W.C.; Cámara, M. Look Who’s Talking: Communication and Quorum Sensing in the Bacterial World. Philos. Trans. R. Soc. B Biol. Sci. 2007, 362, 1119–1134. [Google Scholar] [CrossRef] [Green Version]
- Leblanc, C.; Caumont-Sarcos, A.; Comeau, A.M.; Krisch, H.M. Isolation and Genomic Characterization of the First Phage Infecting Iodobacteria: ΦPLPE, a Myovirus Having a Novel Set of Features. Environ. Microbiol. Rep. 2009, 1, 499–509. [Google Scholar] [CrossRef]
- Silpe, J.E.; Bassler, B.L. A Host-Produced Quorum-Sensing Autoinducer Controls a Phage Lysis-Lysogeny Decision. Cell 2019, 176, 268–280.e13. [Google Scholar] [CrossRef] [Green Version]
- Shah, M.; Taylor, V.L.; Bona, D.; Tsao, Y.; Stanley, S.Y.; Pimentel-Elardo, S.M.; McCallum, M.; Bondy-Denomy, J.; Howell, P.L.; Nodwell, J.R.; et al. A Phage-Encoded Anti-Activator Inhibits Quorum Sensing in Pseudomonas Aeruginosa. Mol. Cell 2021, 81, 571–583.e6. [Google Scholar] [CrossRef]
- Hendrix, H.; Zimmermann-Kogadeeva, M.; Zimmermann, M.; Sauer, U.; de Smet, J.; Muchez, L.; Lissens, M.; Staes, I.; Voet, M.; Wagemans, J.; et al. Metabolic Reprogramming of Pseudomonas Aeruginosa by Phage-Based Quorum Sensing Modulation. Cell Rep. 2022, 38, 110372. [Google Scholar] [CrossRef] [PubMed]
- Pei, R.; Lamas-Samanamud, G.R. Inhibition of Biofilm Formation by T7 Bacteriophages Producing Quorum-Quenching Enzymes. Appl. Environ. Microbiol. 2014, 80, 5340–5348. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Smet, J.; Hendrix, H.; Blasdel, B.G.; Danis-Wlodarczyk, K.; Lavigne, R. Pseudomonas Predators: Understanding and Exploiting Phage–Host Interactions. Nat. Rev. Microbiol. 2017, 15, 517–530. [Google Scholar] [CrossRef] [PubMed]
- Lammens, E.M.; Nikel, P.I.; Lavigne, R. Exploring the Synthetic Biology Potential of Bacteriophages for Engineering Non-Model Bacteria. Nat. Commun. 2020, 11, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Hatfull, G.F.; Hendrix, R.W. Bacteriophages and Their Genomes. Curr. Opin. Virol. 2011, 1, 298–303. [Google Scholar] [CrossRef] [Green Version]
- Rohwer, F.; Youle, M.; Rohwer, F.; Youle, M. Consider Something Viral in Your Search. Nat. Rev. Microbiol. 2011, 9, 308–309. [Google Scholar] [CrossRef]
- Hargreaves, K.R.; Kropinski, A.M.; Clokie, M.R. Bacteriophage Behavioral Ecology: How Phages Alter Their Bacterial Host’s Habits. Bacteriophage 2014, 4, e29866. [Google Scholar] [CrossRef] [Green Version]
- de Smet, J.; Wagemans, J.; Hendrix, H.; Staes, I.; Visnapuu, A.; Horemans, B.; Aertsen, A.; Lavigne, R. Bacteriophage-mediated Interference of the C-di-GMP Signalling Pathway in Pseudomonas aeruginosa. Microb. Biotechnol. 2021, 14, 967–978. [Google Scholar] [CrossRef]
- Ko, C.-C.; Hatfull, G.F. Identification of Mycobacteriophage Toxic Genes Reveals New Features of Mycobacterial Physiology and Morphology. Sci. Rep. 2020, 10, 14670. [Google Scholar] [CrossRef]
- Miller, E.S.; Kutter, E.; Mosig, G.; Arisaka, F.; Kunisawa, T.; Rüger, W. Bacteriophage T4 Genome. Microbiol. Mol. Biol. Rev. 2003, 67, 86–156. [Google Scholar] [CrossRef] [Green Version]
- Ang, D.; Georgopoulos, C. An ORFan No More: The Bacteriophage T4 39.2 Gene Product, NwgI, Modulates GroEL Chaperone Function. Genetics 2012, 190, 989–1000. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roucourt, B.; Lavigne, R. The Role of Interactions between Phage and Bacterial Proteins within the Infected Cell: A Diverse and Puzzling Interactome. Environ. Microbiol. 2009, 11, 2789–2805. [Google Scholar] [CrossRef] [PubMed]
- Typas, A.; Sourjik, V. Bacterial Protein Networks: Properties and Functions. Nat. Rev. Microbiol. 2015, 13, 559–572. [Google Scholar] [CrossRef] [PubMed]
- Park, S.J.; Cochran, J.R. Protein Engineering and Design; CRC Press: Boca Raton, FL, USA, 2010; ISBN 9781420076585. [Google Scholar]




{kind=link}
{kind=link}
{kind=link}
| Class | Biochemical Activity |
|---|---|
| Lyases | Break the bonds between carbon with another atom (such as oxygen, sulfur, or another carbon atom) by means other than hydrolysis and oxidation. Phages can contain several different lyases, which break down different exopolysaccharides. Identified lysases include: ●→polysaccharide lyases (act on various polyanionic substrates, cleave a hexose-1,4-alpha- or beta-uronic acid sequence by beta-elimination, yielding products in which the non-reducing terminus is modified to an unsaturated uronic acid); ●→alginate lyases (catalyze the alginate degradation by a beta-elimination mechanism, in which the glycosidic bond between the monomers gets broken, resulting in unsaturated oligosaccharides with a double bond between the C4 and C5 carbons of the sugar rings); ●→exopolygalacturonic acid lyases; ●→guluronan lyases; ●→hyaluronate lyases/hyaluronidases (degradation of hyaluronate); ●→pectate/pectin lyases. |
| Hydrolases | Cleave a covalent bond by using a water molecule. Bacteriophage hydrolases degrade both bacterial cell walls and exopolysaccharides. Examples of hydrolases of bacteriophages: ●→endorhamnosidases (hydrolysis of O-polysaccharide chain); ●→endosialidases (glycosyl hydrolases that specifically cleave polysialic acid); ●→amylases (glycoside hydrolases); ●→galactosidases (galactoside hydrolases); ●→glucosidases (hydrolases of complex carbohydrates, resulting in monosaccharides); ●→pullulanases (hydrolyses glycosidic bonds of polysaccharides); ●→dextranases (glucosidase); ●→cellulases (hydrolyzes β(1 → 4)-d-glucoside bond of cellulose); ●→sialidases (hydrolyzes the α-linked sialic acid from a variety of molecules, such as oligosaccharides and glycoproteins); ●→xylosidases (catalyze the hydrolysis of α- or β-glycosidic linkages); ●→levanase (hydrolysis of (2 → 6)-beta-D-fructofuranosidic bond in (2 → 6)-beta-D-fructans (levans)). |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Visnapuu, A.; Van der Gucht, M.; Wagemans, J.; Lavigne, R. Deconstructing the Phage–Bacterial Biofilm Interaction as a Basis to Establish New Antibiofilm Strategies. Viruses 2022, 14, 1057. https://doi.org/10.3390/v14051057
Visnapuu A, Van der Gucht M, Wagemans J, Lavigne R. Deconstructing the Phage–Bacterial Biofilm Interaction as a Basis to Establish New Antibiofilm Strategies. Viruses. 2022; 14(5):1057. https://doi.org/10.3390/v14051057
Chicago/Turabian StyleVisnapuu, Annegrete, Marie Van der Gucht, Jeroen Wagemans, and Rob Lavigne. 2022. "Deconstructing the Phage–Bacterial Biofilm Interaction as a Basis to Establish New Antibiofilm Strategies" Viruses 14, no. 5: 1057. https://doi.org/10.3390/v14051057