
A Qualitative Risk Assessment for Bluetongue Disease and African Horse Sickness: The Risk of Entry and Exposure at a UK Zoo

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Risk Assessment Methods
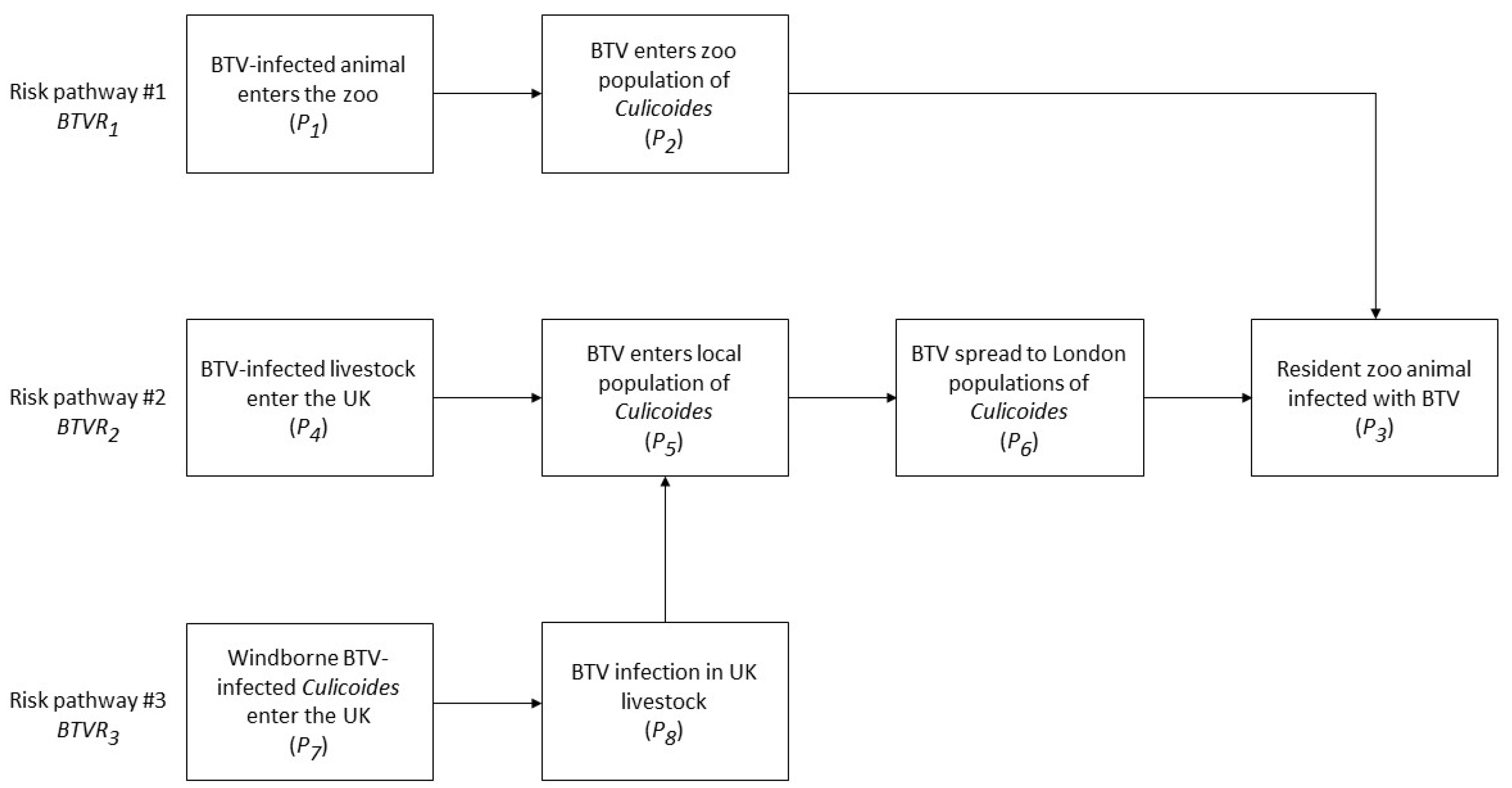
2.2. Risk Pathways
2.2.1. Qualitative Probabilities for BTV Risk Pathways
Estimation of P1: Probability of BTV-Infected Animal Entering the Zoo
Estimation of P2: Probability of BTV Entering Culicoides Population in the Zoo
Estimation of P3: Probability of Resident Zoo Animal Becoming Infected with BTV
Estimation of P4: Probability of BTV-Infected Livestock Entering the UK
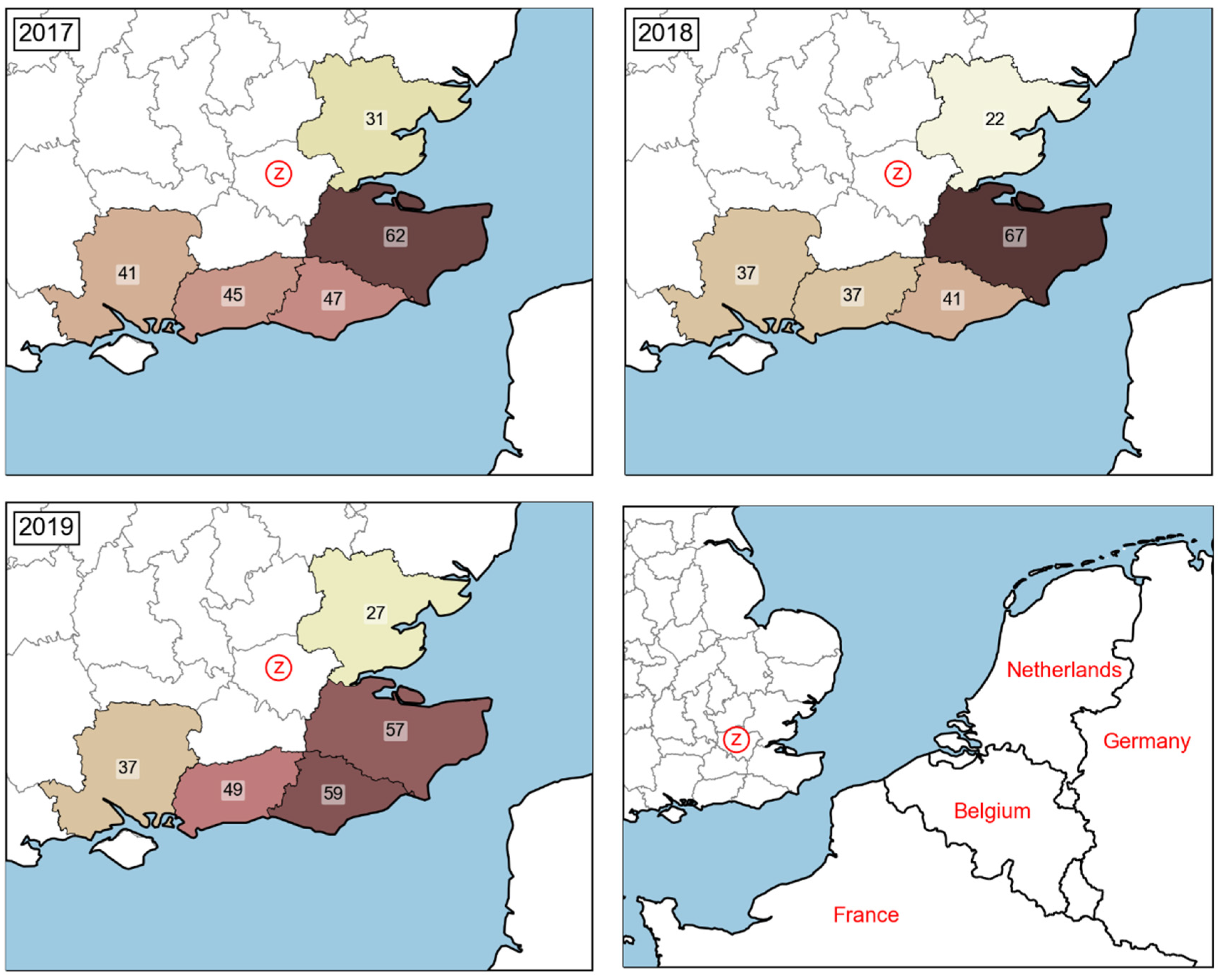
Estimation of P5: Probability of BTV Entering Local Culicoides Populations
Estimation of P6: Probability of Spread of BTV to London Culicoides Populations
Estimation of P7: Probability of Windborne BTV-Infected Culicoides Entering the UK
Estimation of P8: Probability of BTV-Infection in Native UK Livestock following Windborne Incursion of Infected Culicoides
2.2.2. Qualitative Probabilities for AHSV Risk Pathways
Estimation of P9: Probability of AHSV-Infected Animal Entering the Zoo
Estimation of P10: Probability of AHSV Entering Culicoides Populations in the Zoo
Estimation of P11: Probability of a Resident Zoo Animal Becoming Infected with AHSV
Estimation of P12: Probability of AHSV-Infected Equid Entering the UK
Estimation of P13: Probability of AHSV Entering Local UK Culicoides Populations
Estimation of P14: Probability of Spread of AHSV to London Culicoides Populations
3. Results
3.1. BTV Risk Pathways
3.2. AHSV Risk Pathways
4. Discussion
4.1. Key Assumptions
4.2. Negligible Risk Pathways
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Baylis, M. Potential impact of climate change on emerging vector-borne and other infections in the UK. Environ. Health 2017, 16, 112. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Connolly, C.; Keil, R.; Ali, S.H. Extended urbanisation and the spatialities of infectious disease: Demographic change, infrastructure and governance. Urban Stud. 2020, 58, 245–263. [Google Scholar] [CrossRef]
- Semenza, J.C.; Suk, J.E. Vector-Borne diseases and climate change: A European perspective. FEMS Microbiol. Lett. 2017, 365, fnx244. [Google Scholar] [CrossRef] [PubMed]
- Carpenter, S.; Wilson, A.; Mellor, P.S. Culicoides and the emergence of bluetongue virus in northern Europe. Trends Microbiol. 2009, 17, 172–178. [Google Scholar] [CrossRef]
- Jameson, L.J.; Morgan, P.J.; Medlock, J.M.; Watola, G.; Vaux, A.G.C. Importation of Hyalomma marginatum, vector of Crimean-Congo haemorrhagic fever virus, into the United Kingdom by migratory birds. Ticks Tick-Borne Dis. 2012, 3, 95–99. [Google Scholar] [CrossRef] [PubMed]
- Folly, A.J.; Lawson, B.; Lean, F.Z.; McCracken, F.; Spiro, S.; John, S.K.; Heaver, J.P.; Seilern-Moy, K.; Masters, N.; Hernández-Triana, L.M.; et al. Detection of Usutu virus infection in wild birds in the United Kingdom, 2020. Eurosurveillance 2020, 25, 2001732. [Google Scholar] [CrossRef]
- Holding, M.; Dowall, S.; Hewson, R. Detection of tick-borne encephalitis virus in the UK. Lancet 2020, 395, 411. [Google Scholar] [CrossRef] [Green Version]
- Golding, N.; Nunn, M.A.; Medlock, J.M.; Purse, B.V.; Vaux, A.G.C.; Schäfer, S.M. West Nile virus vector Culex modestus established in southern England. Parasites Vectors 2012, 5, 32. [Google Scholar] [CrossRef] [Green Version]
- Gale, P.; Stephenson, B.; Brouwer, A.; Martinez, M.; de la Torre, A.; Bosch, J.; Foley-Fisher, M.; Bonilauri, P.; Lindström, A.; Ulrich, R.G.; et al. Impact of climate change on risk of incursion of Crimean-Congo haemorrhagic fever virus in livestock in Europe through migratory birds. J. Appl. Microbiol. 2012, 112, 246–257. [Google Scholar] [CrossRef] [PubMed]
- Faes, C.; van der Stede, Y.; Guis, H.; Staubach, C.; Ducheyne, E.; Hendrickx, G.; Mintiens, K. Factors affecting bluetongue serotype 8 spread in Northern Europe in 2006: The geographical epidemiology. Prev. Vet. Med. 2013, 110, 149–158. [Google Scholar] [CrossRef]
- Pioz, M.; Guis, H.; Crespin, L.; Gay, E.; Calavas, D.; Durand, B.; Abrial, D.; Ducrot, C. Why did bluetongue spread the way it did? Environmental factors influencing the velocity of bluetongue virus serotype 8 epizootic wave in France. PLoS ONE 2012, 7, e43360. [Google Scholar] [CrossRef]
- Wilson, A.; Mellor, P. Bluetongue in Europe: Vectors, epidemiology and climate change. Parasitol. Res. 2008, 103, 69–77. [Google Scholar] [CrossRef]
- Gale, P.; Brouwer, A.; Ramnial, V.; Kelly, L.; Kosmider, R.; Fooks, A.R.; Snary, E.L. Assessing the impact of climate change on vector-borne viruses in the EU through the elicitation of expert opinion. Epidemiol. Infect. 2010, 138, 214–225. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gould, E.A.; Higgs, S.; Buckley, A.; Gritsun, T.S. Potential arbovirus emergence and implications for the United Kingdom. Emerg. Infect. Dis. 2006, 12, 549–555. [Google Scholar] [CrossRef] [PubMed]
- Szmaragd, C.; Wilson, A.J.; Carpenter, S.; Wood, J.L.N.; Mellor, P.S.; Gubbins, S. The spread of bluetongue virus serotype 8 in Great Britain and its control by vaccination. PLoS ONE 2010, 5, e9353. [Google Scholar] [CrossRef] [Green Version]
- Carpenter, S.; Wilson, A.; Mellor, P. Bluetongue virus and Culicoides in the UK: The impact of research on policy. Outlooks Pest Manag. 2009, 20, 161–164. [Google Scholar] [CrossRef]
- Szmaragd, C.; Gunn, G.J.; Gubbins, S. Assessing the consequences of an incursion of a vector-borne disease. II. Spread of bluetongue in Scotland and impact of vaccination. Epidemics 2010, 2, 139–147. [Google Scholar] [CrossRef] [PubMed]
- King, B.; O’Shea Brown, T.; Tarlinton, R.; Daly, J.M. Seroprevalence of Schmallenberg virus in the United Kingdom and the Republic of Ireland: 2011–2013. Vet. Microbiol. 2015, 180, 36–40. [Google Scholar] [CrossRef] [PubMed]
- Southwell, R.M.; Sherlock, K.; Baylis, M. Cross-sectional study of British wild deer for evidence of Schmallenberg virus infection. Vet. Rec. 2020, 187, e64. [Google Scholar] [CrossRef]
- Hoffmann, B.; Scheuch, M.; Höper, D.; Jungblut, R.; Holsteg, M.; Schirrmeier, H.; Eschbaumer, M.; Goller, K.V.; Wernike, K.; Fischer, M.; et al. Novel orthobunyavirus in cattle, Europe, 2011. Emerg. Infect. Dis. 2012, 18, 469–472. [Google Scholar] [CrossRef] [PubMed]
- Hateley, G. Bluetongue in northern Europe: The story so far. Practice 2009, 31, 202–209. [Google Scholar] [CrossRef]
- Sailleau, C.; Bréard, E.; Viarouge, C.; Vitour, D.; Romey, A.; Garnier, A.; Fablet, A.; Lowenski, S.; Gorna, K.; Caignard, G.; et al. Re-emergence of bluetongue virus serotype 8 in France, 2015. Transbound. Emerg. Dis. 2017, 64, 998–1000. [Google Scholar] [CrossRef] [PubMed]
- Vandenbussche, F.; Bourg, M.; Mathijs, E.; Lefebvre, D.J.; Leeuw, I.D.; Haegeman, A.; Aerts, L.; Borm, S.V.; Clercq, K.D.; Matthijnssens, J. Nearly complete genome sequences of two bluetongue viruses isolated during the 2020 outbreak in the Grand Duchy of Luxembourg. Microbiol. Resour. Announc. 2021, 10, e00210-21. [Google Scholar] [CrossRef]
- Ries, C.; Domes, U.; Janowetz, B.; Böttcher, J.; Burkhardt, K.; Miller, T.; Beer, M.; Hoffmann, B. Isolation and cultivation of a new isolate of BTV-25 and presumptive evidence for a potential persistent infection in healthy goats. Viruses 2020, 12, 983. [Google Scholar] [CrossRef] [PubMed]
- Carpenter, S.; Groschup, M.H.; Garros, C.; Felippe-Bauer, M.L.; Purse, B.V. Culicoides biting midges, arboviruses and public health in Europe. Antivir. Res 2013, 100, 102–113. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carpenter, S.; Veronesi, E.; Mullens, B.; Venter, G. Vector competence of Culicoides for arboviruses: Three major periods of research, their influence on current studies and future directions. Rev. Sci. Tech. 2015, 34, 97–112. [Google Scholar] [CrossRef]
- Veldhuis, A.M.B.; Santman-Berends, I.M.G.A.; Gethmann, J.M.; Mars, M.H.; van Wuyckhuise, L.; Vellema, P.; Holsteg, M.; Höreth-Böntgen, D.; Conraths, F.J.; van Schaik, G. Schmallenberg virus epidemic: Impact on milk production, reproductive performance and mortality in dairy cattle in the Netherlands and Kleve district, Germany. Prev. Vet. Med. 2014, 116, 412–422. [Google Scholar] [CrossRef] [PubMed]
- Maclachlan, N.J.; Zientara, S.; Wilson, W.C.; Richt, J.A.; Savini, G. Bluetongue and epizootic hemorrhagic disease viruses: Recent developments with these globally re-emerging arboviral infections of ruminants. Curr. Opin. Virol. 2019, 34, 56–62. [Google Scholar] [CrossRef] [PubMed]
- Flannery, J.; Frost, L.; Fay, P.; Hicks, H.; Henstock, M.; Smreczak, M.; Orłowska, A.; Rajko-Nenow, P.; Darpel, K.; Batten, C. BTV-14 infection in sheep elicits viraemia with mild clinical symptoms. Microorganisms 2020, 8, 892. [Google Scholar] [CrossRef] [PubMed]
- Eschbaumer, M.; Hoffmann, B.; Moss, A.; Savini, G.; Leone, A.; König, P.; Zemke, J.; Conraths, F.; Beer, M. Emergence of bluetongue virus serotype 6 in Europe—German field data and experimental infection of cattle. Vet. Microbiol. 2010, 143, 189–195. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Clercq, K.; Mertens, P.; De Leeuw, I.; Oura, C.; Houdart, P.; Potgieter, A.C.; Maan, S.; Hooyberghs, J.; Batten, C.; Vandemeulebroucke, E.; et al. Emergence of bluetongue serotypes in Europe, Part 2: The occurrence of a BTV-11 strain in Belgium. Transbound. Emerg. Dis. 2009, 56, 355–361. [Google Scholar] [CrossRef] [PubMed]
- Schulz, C.; Bréard, E.; Sailleau, C.; Jenckel, M.; Viarouge, C.; Vitour, D.; Palmarini, M.; Gallois, M.; Höper, D.; Hoffmann, B.; et al. Bluetongue virus serotype 27: Detection and characterization of two novel variants in Corsica, France. J. Gen. Virol. 2016, 97, 2073–2083. [Google Scholar] [CrossRef] [PubMed]
- Pascual-Linaza, A.V.; Martínez-López, B.; Pfeiffer, D.U.; Moreno, J.C.; Sanz, C.; Sánchez-Vizcaíno, J.M. Evaluation of the spatial and temporal distribution of and risk factors for bluetongue serotype 1 epidemics in sheep Extremadura (Spain), 2007–2011. Prev. Vet. Med. 2014, 116, 279–295. [Google Scholar] [CrossRef]
- Bluetongue Virus Serotype 8 in West Europe: Updated Outbreak Assessment #5; Department for Environment, Food and Rural Affairs: London, UK, 2020.
- GB Bluetongue Virus Disease Control Strategy; Department for Environment, Food and Rural Affairs: London, UK, 2014.
- Grace, K.E.F.; Papadopoulou, C.; Floyd, T.; Avigad, R.; Collins, S.; White, E.; Batten, C.; Flannery, J.; Gubbins, S.; Carpenter, S.T. Risk-Based surveillance for bluetongue virus in cattle on the south coast of England in 2017 and 2018. Vet. Rec. 2020, 187, e96. [Google Scholar] [CrossRef] [PubMed]
- Medlock, J.M.; Hansford, K.M.; Vaux, A.G.C.; Cull, B.; Gillingham, E.; Leach, S. Assessment of the public health threats posed by vector-borne disease in the United Kingdom (UK). Int. J. Environ. Res. Public Health 2018, 15, 2145. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sanders, C.J.; Shortall, C.R.; England, M.; Harrington, R.; Purse, B.; Burgin, L.; Carpenter, S.; Gubbins, S. Long-Term shifts in the seasonal abundance of adult Culicoides biting midges and their impact on potential arbovirus outbreaks. J. Appl. Ecol. 2019, 56, 1649–1660. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ewing, D.A.; Purse, B.V.; Cobbold, C.A.; White, S.M. A novel approach for predicting risk of vector-borne disease establishment in marginal temperate environments under climate change: West Nile virus in the UK. J. R. Soc. Interface 2021, 18, 20210049. [Google Scholar] [CrossRef] [PubMed]
- Carpenter, S.; Mellor, P.S.; Fall, A.G.; Garros, C.; Venter, G.J. African horse sickness virus: History, transmission, and current status. Annu. Rev. Entomol. 2017, 62, 343–358. [Google Scholar] [CrossRef] [PubMed]
- OIE. OIE Technical Disease Card: African Horse Sickness; OIE: Paris, France, 2013. [Google Scholar]
- Mellor, P.S.; Boned, J.; Hamblin, C.; Graham, S. Isolations of African Horse Sickness Virus from Vector Insects Made during the 1988 Epizootic in Spain. Epidemiol. Infect. 1990, 105, 447–454. [Google Scholar] [CrossRef] [Green Version]
- Lubroth, J. African horse sickness and the epizootic in Spain, 1987. Equine Pract. 1988, 10, 26–33. [Google Scholar]
- Porphyre, T.; Grewar, J.D. Assessing the potential of plains zebra to maintain African horse sickness in the Western Cape Province, South Africa. PLoS ONE 2019, 14, e0222366. [Google Scholar] [CrossRef] [Green Version]
- Grewar, J.D.; Kotze, J.L.; Parker, B.J.; van Helden, L.S.; Weyer, C.T. An entry risk assessment of African horse sickness virus into the controlled area of South Africa through the legal movement of equids. PLoS ONE 2021, 16, e0252117. [Google Scholar] [CrossRef] [PubMed]
- OIE. Guidelines on Preparedness and Implementation of Emergency Vaccination in the Asian Region; OIE: Paris, France, 2020; p. 32. [Google Scholar]
- Sergeant, E.S.; Grewar, J.D.; Weyer, C.T.; Guthrie, A.J. Quantitative risk assessment for African horse sickness in live horses exported from South Africa. PLoS ONE 2016, 11, e0151757. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Vos, C.J.; Hoek, C.A.; Nodelijk, G. Risk of introducing African horse sickness virus into the Netherlands by international equine movements. Prev. Vet. Med. 2012, 106, 108–122. [Google Scholar] [CrossRef] [PubMed]
- African Horse Sickness Control Strategy for Great Britain Version 1.1 ed.; Department for Environment, Food and Rural Affairs: London, UK, 2013.
- Maan, S.; Maan, N.S.; Ross-smith, N.; Batten, C.A.; Shaw, A.E.; Anthony, S.J.; Samuel, A.R.; Darpel, K.E.; Veronesi, E.; Oura, C.A.L.; et al. Sequence analysis of bluetongue virus serotype 8 from the Netherlands 2006 and comparison to other European strains. Virology 2008, 377, 308–318. [Google Scholar] [CrossRef] [Green Version]
- Mintiens, K.; Méroc, E.; Mellor, P.S.; Staubach, C.; Gerbier, G.; Elbers, A.R.W.; Hendrickx, G.; De Clercq, K. Possible routes of introduction of bluetongue virus serotype 8 into the epicentre of the 2006 epidemic in north-western Europe. Prev. Vet. Med. 2008, 87, 131–144. [Google Scholar] [CrossRef]
- Tarlinton, R.; Daly, J.; Dunham, S.; Kydd, J. The challenge of Schmallenberg virus emergence in Europe. Vet. J. 2012, 194, 10–18. [Google Scholar] [CrossRef] [PubMed]
- Wilson, A.J.; Mellor, P.S. Bluetongue in Europe: Past, present and future. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2009, 364, 2669–2681. [Google Scholar] [CrossRef] [PubMed]
- Carpenter, S. Culicoides and the spread of bluetongue: Finding the needle in the haystack. Vet. Rec. 2011, 168, 238–239. [Google Scholar] [CrossRef]
- Vilar, M.J.; Guis, H.; Krzywinski, J.; Sanderson, S.; Baylis, M. Culicoides vectors of bluetongue virus in Chester Zoo. Vet. Rec. 2011, 168, 242. [Google Scholar] [CrossRef] [PubMed]
- England, M.E.; Pearce-Kelly, P.; Brugman, V.A.; King, S.; Gubbins, S.; Sach, F.; Sanders, C.J.; Masters, N.J.; Denison, E.; Carpenter, S. Culicoides species composition and molecular identification of host blood meals at two zoos in the UK. Parasites Vectors 2020, 13, 139. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Terrestrial Animal Health Code; OIE: Paris, France, 2019; Volume 1.
- EFSA. Opinion of the Scientific Panel Animal Health and Welfare (AHAW) related with the Migratory Birds and their Possible Role in the Spread of Highly Pathogenic Avian Influenza. EFSA J. 2006, 357, 1–46. [Google Scholar]
- Gale, P.; Kelly, L.; Snary, E.L. Qualitative assessment of the entry of capripoxviruses into Great Britain from the European Union through importation of ruminant hides, skins and wool. Microb. Risk Anal. 2016, 1, 13–18. [Google Scholar] [CrossRef] [Green Version]
- Niedbalski, W. Monitoring studies of bluetongue disease in ruminants imported to Poland from EU. Pol. J. Vet. Sci. 2010, 13, 333–336. [Google Scholar] [PubMed]
- Defra. Bluetongue Virus Detected and Dealt with in Imported Cattle; Department for Environment, Food and Rural Affairs: London, UK, 2017.
- Defra. Bluetongue Virus Detected and Dealt with in Two Imported Cattle; Department for Environment, Food and Rural Affairs: London, UK, 2018.
- EAZA. The Modern Zoo: Foundations for Management and Development; EAZA: Amsterdam, The Netherlands, 2013. [Google Scholar]
- House, J.A.; Groocock, C.M.; Campbell, C.H. Antibodies to bluetongue viruses in animals imported into United States zoological gardens. Can. J. Comp. Med. Rev. Can. Med. Comp. 1982, 46, 154–159. [Google Scholar]
- BIAZA. Animal Transfer Policy (ATP); BIAZA: London, UK, 2014. [Google Scholar]
- Council Directive 92/65/EEC of 13 July 1992 Laying Down Animal Health Requirements Governing Trade in and Imports into the Community of Animals, Semen, Ova and Embryos Not Subject to Animal Health Requirements Laid Down in Specific Community Rules Referred to in Annex A (I) to Directive 90/425/EEC; FAO: Rome, Italy, 1992.
- EAZA. EAZA Population Management Manual: Standards, procedures and guidelines for population management within EAZA. In Appendix 22: EAZA Guidelines for Decision Making when Importing EEP Animals from the Wild; EAZA: Amsterdam, The Netherlands, 2020. [Google Scholar]
- Species360 Zoological Information Management System (ZIMS). 2019. Available online: zims.Species360.org (accessed on 25 January 2021).
- Alexander, K.A.; Kat, P.W.; House, J.; House, C.; O’Brien, S.J.; Laurenson, M.K.; McNutt, J.W.; Osburn, B.I. African horse sickness and African carnivores. Vet. Microbiol. 1995, 47, 133–140. [Google Scholar] [CrossRef]
- Oura, C.A.L.; El Harrak, M. Midge-transmitted bluetongue in domestic dogs. Epidemiol. Infect. 2011, 139, 1396–1400. [Google Scholar] [CrossRef] [Green Version]
- Baba, S.; Olaleye, O.; Ayanbadejo, O. Haemagglutination-inhibiting antibodies against African horse sickness virus in domestic animals in Nigeria. Vet. Res. 1993, 24, 483–487. [Google Scholar]
- OIE. World Animal Health Information Database (WAHIS Interface). Disease Distribution Maps: Bluetongue. Available online: https://www.oie.int/wahis_2/public/wahid.php/Diseaseinformation/Diseasedistributionmap (accessed on 10 July 2020).
- ProMED-Mail. Bluetongue—Europe. Available online: http://www.promedmail.org (accessed on 15 June 2020).
- Darpel, K.E. The Bluetongue Virus ‘Ruminant Host—Insect Vector’ Transmission Cycle; the Role of Culicoides Saliva Proteins in Infection. Ph.D. Thesis, Royal Veterinary College, London, UK, 2007. [Google Scholar]
- Takamatsu, H.; Mellor, P.S.; Mertens, P.P.C.; Kirkham, P.A.; Burroughs, J.N.; Parkhouse, R.M.E. A possible overwintering mechanism for bluetongue virus in the absence of the insect vector. J. Gen. Virol. 2003, 84, 227–235. [Google Scholar] [CrossRef]
- Bonneau, K.R.; DeMaula, C.D.; Mullens, B.A.; MacLachlan, N.J. Duration of viraemia infectious to Culicoides sonorensis in bluetongue virus-infected cattle and sheep. Vet. Microbiol. 2002, 88, 115–125. [Google Scholar] [CrossRef]
- MacLachlan, N.J. Bluetongue: Pathogenesis and duration of viraemia. Vet. Ital. 2004, 40, 462–467. [Google Scholar] [PubMed]
- Koumbati, M.; Mangana, O.; Nomikou, K.; Mellor, P.S.; Papadopoulos, O. Duration of bluetongue viraemia and serological responses in experimentally infected European breeds of sheep and goats. Vet. Microbiol. 1999, 64, 277–285. [Google Scholar] [CrossRef]
- Singer, R.S.; MacLachlan, N.J.; Carpenter, T.E. Maximal predicted duration of viremia in bluetongue virus-infected cattle. J. Vet. Diagn. Investig. 2001, 13, 43–49. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Regulation (EU) 2016/429 of the European Parliament and of the Council of 9 March 2016 on Transmissible Animal Diseases and Amending and Repealing Certain Acts in the Area of Animal Health (‘Animal Health Law’); EU: Brussels, Belgium, 2021.
- Commission Delegated Regulation (EU) 2020/689 of 17 December 2019 Supplementing Regulation (EU) 2016/429 of the European Parliament and of the Council as Regards Rules for Surveillance, Eradication Programmes, and Disease-Free Status for Certain Listed and Emerging Diseases; EU: Brussels, Belgium, 2021.
- Searle, K.R.; Barber, J.; Stubbins, F.; Labuschagne, K.; Carpenter, S.; Butler, A.; Denison, E.; Sanders, C.; Mellor, P.S.; Wilson, A.; et al. Environmental drivers of Culicoides phenology: How important is species-specific variation when determining disease policy? PLoS ONE 2014, 9, e111876. [Google Scholar] [CrossRef] [Green Version]
- Sanders, C.J.; Shortall, C.R.; Gubbins, S.; Burgin, L.; Gloster, J.; Harrington, R.; Reynolds, D.R.; Mellor, P.S.; Carpenter, S. Influence of season and meteorological parameters on flight activity of Culicoides biting midges. J. Appl. Ecol. 2011, 48, 1355–1364. [Google Scholar] [CrossRef]
- England, M.E.; (The Pirbright Institute, Woking, UK). Personal communication, 2021.
- Cuéllar, A.C.; Kjær, L.J.; Kirkeby, C.; Skovgard, H.; Nielsen, S.A.; Stockmarr, A.; Andersson, G.; Lindstrom, A.; Chirico, J.; Lühken, R.; et al. Spatial and temporal variation in the abundance of Culicoides biting midges (Diptera: Ceratopogonidae) in nine European countries. Parasites Vectors 2018, 11, 112. [Google Scholar] [CrossRef] [PubMed]
- Mehlhorn, H.; Walldorf, V.; Klimpel, S.; Jahn, B.; Jaeger, F.; Eschweiler, J.; Hoffmann, B.; Beer, M. First occurrence of Culicoides obsoletus-transmitted bluetongue virus epidemic in Central Europe. Parasitol. Res. 2007, 101, 219–228. [Google Scholar] [CrossRef]
- Tugwell, L.A.; England, M.E.; Gubbins, S.; Sanders, C.J.; Stokes, J.E.; Stoner, J.; Graham, S.P.; Blackwell, A.; Darpel, K.E.; Carpenter, S. Thermal limits for flight activity of field-collected Culicoides in the United Kingdom defined under laboratory conditions. Parasites Vectors 2021, 14, 55. [Google Scholar] [CrossRef] [PubMed]
- Carpenter, S.; Wilson, A.; Barber, J.; Veronesi, E.; Mellor, P.; Venter, G.; Gubbins, S. Temperature dependence of the extrinsic incubation period of Orbiviruses in Culicoides biting midges. PLoS ONE 2011, 6, e27987. [Google Scholar] [CrossRef] [PubMed]
- ZSL. London Zoo January 2019 Stocklist. Available online: https://www.zsl.org/sites/default/files/media/2019-04/ZSL%20Zoological%20Society%20of%20London%20-%20January%202019%20stocklist.pdf (accessed on 10 May 2020).
- Sanderson, S. Bluetongue in non-domestic ruminants: Experiences gained in EAZA zoos during the 2007 & 2008 BTV8 and BTV1 epizootics. In Transmissible Diseases Handbook; EAZWV: Brussels, Belgium, 2010. [Google Scholar]
- Elbers, A.R.W.; Backx, A.; Mintiens, K.; Gerbier, G.; Staubach, C.; Hendrickx, G.; van der Spek, A. Field observations during the bluetongue serotype 8 epidemic in 2006: II. Morbidity and mortality rate, case fatality and clinical recovery in sheep and cattle in the Netherlands. Prev. Vet. Med. 2008, 87, 31–40. [Google Scholar] [CrossRef]
- Conraths, F.J.; Gethmann, J.M.; Staubach, C.; Mettenleiter, T.C.; Beer, M.; Hoffmann, B. Epidemiology of bluetongue virus serotype 8, Germany. Emerg. Infect. Dis. 2009, 15, 433–435. [Google Scholar] [CrossRef] [PubMed]
- Elbers, A.R.W.; Backx, A.; Meroc, E.; Gerbier, G.; Staubach, C.; Hendrickx, G.; van der Spek, A.; Mintiens, K. Field observations during the bluetongue serotype 8 epidemic in 2006: I. Detection of first outbreaks and clinical signs in sheep and cattle in Belgium, France and the Netherlands. Prev. Vet. Med. 2008, 87, 21–30. [Google Scholar] [CrossRef] [PubMed]
- Climate-Data.org: London Climate Graph. Available online: https://en.climate-data.org/ (accessed on 25 June 2020).
- Bluetongue confirmed in UK cattle. Vet. Rec. 2017, 181, 435. [CrossRef] [PubMed]
- Batten, C.; Flannery, J. Combatting the looming threat of bluetongue. Vet. Pract. 2019, 51, 35–37. [Google Scholar]
- European Commission (Ed.) TRACES; Animal and Plant Health Agency: London, UK.
- Regulation (EU) 2017/625 of the European Parliament and of the Council of 15 March 2017 on Official Controls and Other Official Activities Performed to Ensure the Application of Food and Feed Law, Rules on Animal Health and Welfare, Plant Health and Plant Protection Products, Amending Regulations (EC) No 999/2001, (EC) No 396/2005, (EC) No 1069/2009, (EC) No 1107/2009, (EU) No 1151/2012, (EU) No 652/2014, (EU) 2016/429 and (EU) 2016/2031 of the European Parliament and of the Council, Council Regulations (EC) No 1/2005 and (EC) No 1099/2009 and Council Directives 98/58/EC, 1999/74/EC, 2007/43/EC, 2008/119/EC and 2008/120/EC, and Repealing Regulations (EC) No 854/2004 and (EC) No 882/2004 of the European Parliament and of the Council, Council Directives 89/608/EEC, 89/662/EEC, 90/425/EEC, 91/496/EEC, 96/23/EC, 96/93/EC and 97/78/EC and Council Decision 92/438/EEC (Official Controls Regulation); EU: Brussels, Belgium, 2019.
- Mayo, C.; Lee, J.; Kopanke, J.; MacLachlan, N.J. A review of potential bluetongue virus vaccine strategies. Vet. Microbiol. 2017, 206, 84–90. [Google Scholar] [CrossRef] [PubMed]
- Flannery, J.; Sanz-Bernardo, B.; Ashby, M.; Brown, H.; Carpenter, S.; Cooke, L.; Corla, A.; Frost, L.; Gubbins, S.; Hicks, H.; et al. Evidence of reduced viremia, pathogenicity and vector competence in a re-emerging European strain of bluetongue virus serotype 8 in sheep. Transbound. Emerg. Dis. 2019, 66, 1177–1185. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Steinrigl, A.; Revilla-Fernández, S.; Eichinger, M.; Koefer, J.; Winter, P. Bluetongue virus RNA detection by RT-qPCR in blood samples of sheep vaccinated with a commercially available inactivated BTV-8 vaccine. Vaccine 2010, 28, 5573–5581. [Google Scholar] [CrossRef] [PubMed]
- De Leeuw, I.; Garigliany, M.; Bertels, G.; Willems, T.; Desmecht, D.; De Clercq, K. Bluetongue Virus RNA Detection by Real-Time RT-PCR in Post-Vaccination Samples from Cattle. Transbound. Emerg. Dis. 2015, 62, 157–162. [Google Scholar] [CrossRef] [PubMed]
- Veronesi, E.; Hamblin, C.; Mellor, P.S. Live attenuated bluetongue vaccine viruses in Dorset Poll sheep, before and after passage in vector midges (Diptera: Ceratopogonidae). Vaccine 2005, 23, 5509–5516. [Google Scholar] [CrossRef] [PubMed]
- Carpenter, S.; Lunt, H.L.; Arav, D.; Venter, G.J.; Mellor, P.S. Oral susceptibility to bluetongue virus of Culicoides (Diptera: Ceratopogonidae) from the United Kingdom. J. Med. Entomol. 2006, 43, 73–78. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sanders, C.J.; Harrup, L.E.; Tugwell, L.A.; Brugman, V.A.; England, M.; Carpenter, S. Quantification of within- and between-farm dispersal of Culicoides biting midges using an immunomarking technique. J. Appl. Ecol. 2017, 54, 1429–1439. [Google Scholar] [CrossRef] [Green Version]
- Wittmann, E.J.; Mellor, P.S.; Baylis, M. Effect of temperature on the transmission of orbiviruses by the biting midge, Culicoides sonorensis. Med. Vet. Entomol. 2002, 16, 147–156. [Google Scholar] [CrossRef] [PubMed]
- Hope, A.; Gubbins, S.; Sanders, C.; Barber, J.; Stubbins, F.; Baylis, M.; Carpenter, S. Sheep breed and shearing influences attraction and blood-feeding behaviour of Culicoides (Diptera: Ceratopogonidae) on a UK farm. Parasites Vectors 2018, 11, 473. [Google Scholar] [CrossRef] [PubMed]
- Sumner, T.; Orton, R.J.; Green, D.M.; Kao, R.R.; Gubbins, S. Quantifying the roles of host movement and vector dispersal in the transmission of vector-borne diseases of livestock. PLoS Comput. Biol. 2017, 13, e1005470. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sedda, L.; Brown, H.E.; Purse, B.V.; Burgin, L.; Gloster, J.; Rogers, D.J. A new algorithm quantifies the roles of wind and midge flight activity in the bluetongue epizootic in northwest Europe. Proc. R. Soc. B Biol. Sci. 2012, 279, 2354–2362. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- APHA Livestock Demographic Data Group: Cattle Population Report 2020; APHA: London, UK, 2020.
- APHA Livestock Demographic Data Group: Sheep Population Report 2019; APHA: London, UK, 2019.
- APHA Livestock Demographic Data Group: Goat Population Report 2019; APHA: London, UK, 2019.
- Gloster, J.; Burgin, L.; Witham, C.; Athanassiadou, M.; Mellor, Y.S. Bluetongue in the United Kingdom and northern Europe in 2007 and key issues for 2008. Vet. Rec. 2008, 162, 298–302. [Google Scholar] [CrossRef] [PubMed]
- Burgin, L.E.; Gloster, J.; Sanders, C.; Mellor, P.S.; Gubbins, S.; Carpenter, S. Investigating incursions of bluetongue virus using a model of long-distance Culicoides biting midge dispersal. Transbound. Emerg. Dis. 2013, 60, 263–272. [Google Scholar] [CrossRef] [PubMed]
- Jones, A.; Thomson, D.; Hort, M.; Devenish, B. The U.K. Met Office’s Next-Generation Atmospheric Dispersion Model, NAME III; Springer: Boston, MA, USA, 2007; pp. 580–589. [Google Scholar]
- Gubbins, S.; Carpenter, S.; Baylis, M.; Wood, J.L.N.; Mellor, P.S. Assessing the risk of bluetongue to UK livestock: Uncertainty and sensitivity analyses of a temperature-dependent model for the basic reproduction number. J. R. Soc. Interface 2008, 5, 363–371. [Google Scholar] [CrossRef] [Green Version]
- Getting and Using a Horse Passport. Available online: https://www.gov.uk/horse-passport (accessed on 4 February 2021).
- Sumner, T.; Burgin, L.; Gloster, J.; Gubbins, S. Comparison of pre-emptive and reactive strategies to control an incursion of bluetongue virus serotype 1 to Great Britain by vaccination. Epidemiol. Infect. 2013, 141, 102–114. [Google Scholar] [CrossRef] [PubMed]
- Equines: Health Certificates. Available online: https://www.gov.uk/government/publications/equines-health-certificates (accessed on 4 February 2021).
- Rodriguez, M.; Hooghuis, H.; Castaño, M. African horse sickness in Spain. Vet. Microbiol. 1992, 33, 129–142. [Google Scholar] [CrossRef]
- Van Rijn, P.A.; Maris-Veldhuis, M.A.; Grobler, M.; Wright, I.M.; Erasmus, B.J.; Maartens, L.H.; Potgieter, C.A. Safety and efficacy of inactivated African horse sickness (AHS) vaccine formulated with different adjuvants. Vaccine 2020, 38, 7108–7117. [Google Scholar] [CrossRef]
- Van Sittertl, S.J.; Drew, T.M.; Kotze, J.L.; Strydom, T.; Weyer, C.T.; Guthrie, A.J. Occurrence of African horse sickness in a domestic dog without apparent ingestion of horse meat. J. S. Afr. Vet. Assoc. 2013, 84. [Google Scholar] [CrossRef] [Green Version]
- Capela, R.; Purse, B.V.; Pena, I.; Wittman, E.J.; Margarita, Y.; Capela, M.; Romão, L.; Mellor, P.S.; Baylis, M. Spatial distribution of Culicoides species in Portugal in relation to the transmission of African horse sickness and bluetongue viruses. Med. Vet. Entomol. 2003, 17, 165–177. [Google Scholar] [CrossRef] [PubMed]
- Attoui, H.; Nomikou, K.; Maan, S.; Belaganahalli, M.; Mertens, P.P.C. Orbiviruses. In Reference Module in Biomedical Sciences; Elsevier: Amsterdam, The Netherlands, 2016. [Google Scholar] [CrossRef]
- Attoui, H.; Mohd Jaafar, F. Zoonotic and emerging orbivirus infections. Rev. Sci. Tech. (Int. Off. Epizoot.) 2015, 34, 353–361. [Google Scholar] [CrossRef] [Green Version]
- Awad, F.I.; Amin, M.M.; Salama, S.A.; Khide, S. The role played by Hyalomma dromedarii in the transmission of African horse sickness virus in Egypt. Bull. Anim. Health Prod. Afr. 1981, 29, 337–340. [Google Scholar]
- Mellor, P.S.; Boorman, J. The transmission and geographical spread of African horse sickness and bluetongue viruses. Ann. Trop. Med. Parasitol. 1995, 89, 1–15. [Google Scholar] [CrossRef]
- Mellor, P.S.; Rawlings, P.; Baylis, M.; Wellby, M.P. Effect of temperature on African horse sickness virus infection in Culicoides. Arch. Virol. Suppl. 1998, 14, 155–163. [Google Scholar]
- Jaganmohan, M. Monthly Average Daily Temperatures in the United Kingdom (UK) from 2015 to 2021. Available online: https://www.statista.com/statistics/322658/monthly-average-daily-temperatures-in-the-united-kingdom-uk/ (accessed on 22 September 2021).
- World Bank Group: Climate Knowledge Portal. Available online: https://climateknowledgeportal.worldbank.org/country/south-africa/climate-data-historical (accessed on 22 September 2021).
- OIE. World Animal Health Information Database (WAHIS Interface). Disease Control Measures: African Horse Sickness. Available online: https://www.oie.int/wahis_2/public/wahid.php/Diseasecontrol/measures (accessed on 4 February 2021).
- Spickler, A.R. African Horse Sickness. Available online: http://www.cfsph.iastate.edu/DiseaseInfo/factsheets.php (accessed on 25 January 2021).
- Fassi-Fihri, O.; el Harrak, M.; Fassi-Fehri, M.M. Clinical, virological and immune responses of normal and immunosuppressed donkeys (Equus asinus africanus) after inoculation with African horse sickness virus. Arch. Virol. Suppl. 1998, 14, 49–56. [Google Scholar] [CrossRef]
- Fowler, V.L.; Howson, E.L.A.; Flannery, J.; Romito, M.; Lubisi, A.; Agüero, M.; Mertens, P.; Batten, C.A.; Warren, H.R.; Castillo-Olivares, J. Development of a Novel Reverse Transcription Loop-Mediated Isothermal Amplification Assay for the Rapid Detection of African Horse Sickness Virus. Transbound. Emerg. Dis. 2017, 64, 1579–1588. [Google Scholar] [CrossRef] [Green Version]
- Weyer, C.T.; Grewar, J.D.; Burger, P.; Rossouw, E.; Lourens, C.; Joone, C.; le Grange, M.; Coetzee, P.; Venter, E.; Martin, D.P.; et al. African Horse Sickness Caused by Genome Reassortment and Reversion to Virulence of Live, Attenuated Vaccine Viruses, South Africa, 2004-2014. Emerg. Infect. Dis. 2016, 22, 2087–2096. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Ark, H.; Meiswinkel, R. Subsampling of large light trap catches of Culicoides (Diptera: Ceratopogonidae). Onderstepoort. J. Vet. Res. 1992, 59, 183–189. [Google Scholar]
- Robin, M.; Archer, D.; Garros, C.; Gardès, L.; Baylis, M. The threat of midge-borne equine disease: Investigation of Culicoides species on UK equine premises. Vet. Rec. 2014, 174, 301. [Google Scholar] [CrossRef] [PubMed]
- Baker, T.; Carpenter, S.; Gubbins, S.; Newton, R.; Lo Iacono, G.; Wood, J.; Harrup, L.E. Can insecticide-treated netting provide protection for equids from Culicoides biting midges in the United Kingdom? Parasites Vectors 2015, 8, 604. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Iacono, G.L.; Robin, C.A.; Newton, J.R.; Gubbins, S.; Wood, J.L.N. Where are the horses? With the sheep or cows? Uncertain host location, vector-feeding preferences and the risk of African horse sickness transmission in Great Britain. J. R. Soc. Interface 2013, 10, 20130194. [Google Scholar] [CrossRef]
- Sellers, R.F.; Pedgley, D.E.; Tucker, M.R. Possible spread of African horse sickness on the wind. J. Hyg. 1977, 79, 279–298. [Google Scholar] [CrossRef] [Green Version]
- Allison, K.; Taylor, N.; Upton, M.; Wilsmore, T. African Horse Sickness: Impact on the UK Horse Industry; University of Reading: Reading, UK, 2009. [Google Scholar]
- England, M.E. Understanding the Risks and Factors Associated with the Introduction of Crimean-Congo Haemorrhagic Fever Virus into Great Britain; University of Southampton: Southampton, UK, 2013. [Google Scholar]
- Updated Outbreak Assessment #5: Bluetongue Virus Serotype 8 in West Europe; Department for Environment, Food and Rural Affairs: London, UK, 2020.
- Bréard, E.; Schulz, C.; Sailleau, C.; Bernelin-Cottet, C.; Viarouge, C.; Vitour, D.; Guillaume, B.; Caignard, G.; Gorlier, A.; Attoui, H.; et al. Bluetongue virus serotype 27: Experimental infection of goats, sheep and cattle with three BTV-27 variants reveal atypical characteristics and likely direct contact transmission BTV-27 between goats. Transbound. Emerg. Dis. 2018, 65, 251–263. [Google Scholar] [CrossRef] [PubMed]
- Chaignat, V.; Worwa, G.; Scherrer, N.; Hilbe, M.; Ehrensperger, F.; Batten, C.; Cortyen, M.; Hofmann, M.; Thuer, B. Toggenburg Orbivirus, a new bluetongue virus: Initial detection, first observations in field and experimental infection of goats and sheep. Vet. Microbiol. 2009, 138, 11–19. [Google Scholar] [CrossRef] [Green Version]
- Batten, C.; Darpel, K.; Henstock, M.; Fay, P.; Veronesi, E.; Gubbins, S.; Graves, S.; Frost, L.; Oura, C. Evidence for Transmission of Bluetongue Virus Serotype 26 through Direct Contact. PLoS ONE 2014, 9, e96049. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zientara, S.; Weyer, C.T.; Lecollinet, S. African horse sickness. Rev. Sci. Tech. Off. Int. Epiz. 2015, 34, 315–327. [Google Scholar] [CrossRef] [Green Version]
- Mellor, P.S.; Boorman, J.; Baylis, M. Culicoides biting midges: Their role as arbovirus vectors. Annu. Rev. Entomol. 2000, 45, 307–340. [Google Scholar] [CrossRef]
- Linden, S. MPS Stables—Supply & Delivery Of Feed, Forage & Bedding. Available online: https://www.london.gov.uk/what-we-do/mayors-office-policing-and-crime-mopac/governance-and-decision-making/mopac-decisions-0/mps-stables-supply-delivery-feed-forage-bedding (accessed on 13 October 2021).
- Bevan, E. Hidden in the heart of London: Visit the mounted police’s Narnia-style stables. Horse Hound 2017. [Google Scholar]
- Silver, M. Double-Decker stables and a hidden horse walker: Is this Britain’s most compact yard? Horse Hound 2017. [Google Scholar]
- The Royal Mews, Buckingham Palace. Available online: https://www.rct.uk/visit/the-royal-mews-buckingham-palace (accessed on 13 October 2021).
- Saegerman, C.; Mellor, P.; Uyttenhoef, A.; Hanon, J.-B.; Kirschvink, N.; Haubruge, E.; Delcroix, P.; Houtain, J.-Y.; Pourquier, P.; Vandenbussche, F.; et al. The most likely time and place of introduction of BTV8 into Belgian ruminants. PLoS ONE 2010, 5, e9405. [Google Scholar] [CrossRef]
- Nie, W.-Z.; Li, J.-C.; Li, D.-X.; Gratz, N. A preliminary report of introduced living biting midges (Diptera: Ceratopogonidae) found on entry ships arriving at Qinhuangdao Port, China. Med. Entomol. Zool. 2005, 56, 359–361. [Google Scholar] [CrossRef] [Green Version]
- Hornberger, K.; Ndiritu, N.; Ponce-Brito, L.; Tashu, M.; Watt, T. Kenya’s Cut-Flower Cluster; Harvard University: Cambridge, MA, USA, 2007. [Google Scholar]
- Pascall, D.J.; Nomikou, K.; Bréard, E.; Zientara, S.; Filipe, A.d.S.; Hoffmann, B.; Jacquot, M.; Singer, J.B.; De Clercq, K.; Bøtner, A.; et al. “Frozen evolution” of an RNA virus suggests accidental release as a potential cause of arbovirus re-emergence. PLoS Biol. 2020, 18, e3000673. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Risk Probability | Definition |
|---|---|
| Negligible | Event is so rare that it does not merit consideration |
| Very low | Event is very rare but cannot be excluded |
| Low | Event is rare but does occur |
| Medium | Event occurs regularly |
| High | Event occurs very often |
| Very high | Event occurs almost certainly |
| Scientific Name | Common Name | Total No. of Animals |
|---|---|---|
| Camelus bactrianus | Bactrian camel | 2 |
| Muntiacus reevesi | Chinese muntjac | 2 |
| Giraffa camelopardalis | Giraffe | 3 |
| Okapia johnstoni | Okapi | 3 |
| Capra hircus | Nigerian goat | 4 |
| Capra hircus | West African pygmy goat | 3 |
| Cephalophus natalensis | Red forest duiker | 2 |
| Lama glama | Llama | 2 |
| Vicugna pacos | Alpaca | 1 |
| Total | 22 | |
| Scientific Name | Common Name | Total |
|---|---|---|
| Lycaon pictus | African hunting dog | 7 |
| Equus asinus | Donkey | 2 |
| Equus quagga burchelli | Burchell’s zebra | 2 |
| Equus quagga chapmani | Chapman’s zebra | 2 |
| Camelus bactrianus domestic | Bactrian camel | 2 |
| Total | 15 | |
| Probability | Qualitative Probability |
|---|---|
| BTV-infected animal enters the zoo (P1) | Very low |
| BTV enters zoo population of Culicoides (P2) | Medium |
| Resident zoo animal infected with BTV (P3) | High to very high |
| BTV-infected livestock enters the UK (P4) | Very low |
| BTV enters local population of Culicoides (P5) | Medium |
| BTV spread to London populations of Culicoides (P6) | Low |
| Windborne BTV-infected Culicoides enter the UK (P7) | Low |
| BTV infection in UK livestock (P8) | Medium |
| Risk pathway #1, BTVR1 (P1, P2, P3) | Low |
| Risk pathway #2, BTVR2 (P4, P5, P6, P3) | Low to medium |
| Risk pathway #3, BTVR3 (P7, P8, P5, P6, P3) | Medium |
| Probability | Qualitative Probability |
|---|---|
| AHSV-infected animal enters the zoo (P9) | Very low |
| AHSV enters zoo population of Culicoides (P10) | Medium |
| Resident zoo animal infected with AHSV (P11) | Medium |
| AHSV-infected equid enters the UK (P12) | Very low |
| AHSV enters local population of Culicoides (P13) | Low to medium |
| AHSV spread to London populations of Culicoides (P14) | Very low |
| Risk pathway #1, AHSVR1 (P9, P10, P11) | Very low |
| Risk pathway #2, AHSVR2 (P12, P13, P14, P11) | Very low to low |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nelson, E.; Thurston, W.; Pearce-Kelly, P.; Jenkins, H.; Cameron, M.; Carpenter, S.; Guthrie, A.; England, M. A Qualitative Risk Assessment for Bluetongue Disease and African Horse Sickness: The Risk of Entry and Exposure at a UK Zoo. Viruses 2022, 14, 502. https://doi.org/10.3390/v14030502
Nelson E, Thurston W, Pearce-Kelly P, Jenkins H, Cameron M, Carpenter S, Guthrie A, England M. A Qualitative Risk Assessment for Bluetongue Disease and African Horse Sickness: The Risk of Entry and Exposure at a UK Zoo. Viruses. 2022; 14(3):502. https://doi.org/10.3390/v14030502
Chicago/Turabian StyleNelson, Elisabeth, William Thurston, Paul Pearce-Kelly, Hannah Jenkins, Mary Cameron, Simon Carpenter, Amanda Guthrie, and Marion England. 2022. "A Qualitative Risk Assessment for Bluetongue Disease and African Horse Sickness: The Risk of Entry and Exposure at a UK Zoo" Viruses 14, no. 3: 502. https://doi.org/10.3390/v14030502