
Increase in Hepatitis A Cases Linked to Imported Strains to Rio de Janeiro, Brazil: A Cross-Sectional Study

 ,
,  , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Ethics
2.2. Study Population
2.3. Socio-Epidemiological Data
2.4. Specimens
2.5. Statistical Analysis
2.6. HAV RNA Molecular Detection
2.7. Nucleotide Sequencing, Phylogenetic, and Phylogeographic Analyses
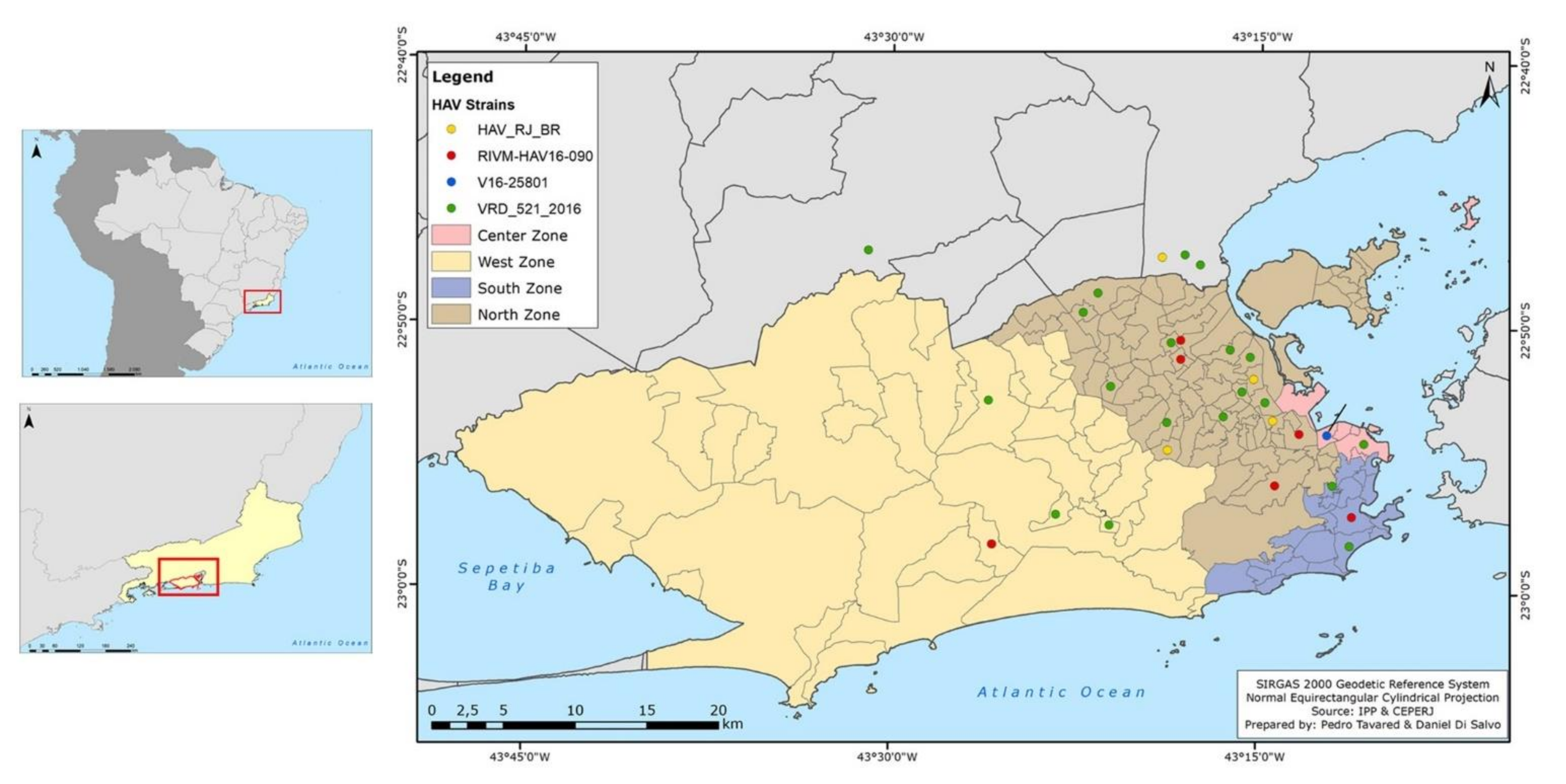
2.8. Map Construction
3. Results
3.1. Samples Description and Socio-Demographic Characteristics and Case Distribution
3.2. HAV Exposure Factors and Co-Infections
3.3. Viral Genome Detection, Genotyping, and Phylogenetic Analyses
3.4. Phylogeographic Analyses and Bayesian Inference
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Vaughan, G.; Rossi, L.M.G.; Forbi, J.C.; de Paula, V.S.; Purdy, M.A.; Xia, G.; Khudyakov, Y.E. Hepatitis A virus: Host interactions, molecular epidemiology and evolution. Infect. Genet. Evol. 2014, 21, 227–243. [Google Scholar] [CrossRef]
- Jacobsen, K.H. Globalization and the Changing Epidemiology of Hepatitis A Virus. Cold Spring Harb. Perspect. Med. 2018, 8, a031716. [Google Scholar] [CrossRef] [Green Version]
- Lima, L.R.; De Almeida, A.J.; Tourinho Rdos, S.; Hasselmann, B.; Ximenez, L.L.; De Paula, V.S. Evidence of hepatitis A virus person-to-person transmission in household outbreaks. PLoS ONE 2014, 9, e102925. [Google Scholar] [CrossRef] [Green Version]
- Corey, L.; Holmes, K.K. Sexual transmission of hepatitis A in homosexual men: Incidence and mechanism. N. Engl. J. Med. 1980, 302, 435–438. [Google Scholar] [CrossRef]
- De Paula, V.S. Laboratory diagnosis of hepatitis A. Future Virol. 2012, 7, 11. [Google Scholar] [CrossRef]
- Souto, F.J.D.; de Brito, W.I.; Fontes, C.J.F. Impact of the single-dose universal mass vaccination strategy against hepatitis A in Brazil. Vaccine 2019, 37, 771–775. [Google Scholar] [CrossRef]
- Lemon, S.M. The natural history of hepatitis A: The potential for transmission by transfusion of blood or blood products. Vox Sang. 1994, 67 (Suppl. 4), 19–23; discussion 4–6. [Google Scholar] [CrossRef]
- Da Silva, S.G.C.; Leon, L.A.A.; Alves, G.; Brito, S.M.; Sandes, V.D.S.; Lima, M.M.A.F.; Nogueira, M.C.; Tavares, R.D.C.B.D.S.; Dobbin, J.; Apa, A.; et al. A Rare Case of Transfusion Transmission of Hepatitis A Virus to Two Patients with Haematological Disease. Transfus. Med. Hemother. 2016, 43, 137–141. [Google Scholar] [CrossRef] [Green Version]
- Cuthbert, J.A. Hepatitis A: Old and new. Clin. Microbiol. Rev. 2001, 14, 38–58. [Google Scholar] [CrossRef] [Green Version]
- De Oliveira, T.M.; Vieira, N.S.G.; Sepp, T.D.S.; Souto, F.J.D. Recent trends in hepatitis A incidence in Brazil. J. Med. Virol. 2020, 92, 1343–1349. [Google Scholar] [CrossRef]
- Rivas, V.; Barrera, A.; Pino, K.; Núñez, R.; Caceres, C.J.; Lopez-Lastra, M.; Soza, A. Hepatitis A outbreak since November 2016 affecting men who have sex with men (MSM) in Chile connected to the current outbreak in MSM in Europe, situation up to October 2017. Eurosurveillance 2018, 23, 18-00060. [Google Scholar] [CrossRef] [Green Version]
- Worl Health Organization. Hepatitis A Outbreaks Mostly Affecting Men Who Have Sex with Men—European Region and the Americas. 2017. Available online: https://www.who.int/csr/don/07-june-2017-hepatitis-a/en/ (accessed on 25 January 2019).
- European Centre for Disease Prevention and Control. Epidemiological Update: Hepatitis a Outbreak in the EU/EEA Mostly Affecting Men Who Have Sex with Men; ECDC: Stockholm, Sweden, 29 September 2017. Available online: http://ecdc.europa.eu/en/news-events/epidemiological-update-hepatitis-outbreak-eueea-mostly-affecting-men-who-have-sex-men (accessed on 20 November 2018).
- Watanabe, S.; Morimoto, N.; Miura, K.; Takaoka, Y.; Nomoto, H.; Tsukui, M.; Isoda, N.; Ohnishi, H.; Nagashima, S.; Takahashi, M.; et al. Full-genome characterization of the RIVM-HAV16-090-like hepatitis A virus strains recovered from Japanese men who have sex with men, with sporadic acute hepatitis A. Hepatol. Res. 2019, 49, 521–530. [Google Scholar] [CrossRef]
- Mello, V.M.; Lago, B.V.; Sousa, P.S.F.; Mello, F.C.A.; Souza, C.B.; Pinto, L.C.M.; Ginuino, C.F.; Fernandes, C.A.S.; Aguiar, S.F.; Villar, L.M.; et al. Hepatitis A Strain Linked to the European Outbreaks During Gay Events between 2016 and 2017, Identified in a Brazilian Homosexual Couple in 2017. Viruses 2019, 11, 281. [Google Scholar] [CrossRef] [Green Version]
- Mariojoules, J.; Castro, G.; Pisano, M.B.; Barbero, P.; Fantilli, A.; Borda, M.; Canna, F.; Barbás, G.; Ré, V.; Mariojouls, J. Hepatitis A outbreak affecting men who have sex with men (MSM) in central Argentina, occurred in July 2017–April 2018, later than the European outbreak. J. Clin. Virol. 2019, 117, 49–53. [Google Scholar] [CrossRef]
- Brazilian Ministry of Health. Viral Hepatitis Epidemiological Bulletin. 2018. Available online: http://www.aids.gov.br/pt-br/pub/2018/boletim-epidemiologico-de-hepatites-virais-2018 (accessed on 15 June 2018).
- Brazilian Ministry of Health. Viral Hepatitis Epidemiological Bulletin. 2019. Available online: http://www.aids.gov.br/pt-br/pub/2019/boletim-epidemiologico-de-hepatites-virais-2019 (accessed on 27 August 2019).
- Venables, W.N.; Smith, D.M.; the R Core Team. An Introduction to R. Notes on R: A Programming Environment for Data Analysis and Graphics Version 4.0.3. 10 October 2020. Available online: https://cran.r-project.org/doc/manuals/r-release/R-intro.pdf (accessed on 15 December 2020).
- De Paula, V.S.; Baptista, M.L.; Lampe, E.; Niel, C.; Gaspar, A.M. Characterization of hepatitis A virus isolates from subgenotypes IA and IB in Rio de Janeiro, Brazil. J. Med. Virol. 2002, 66, 22–27. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular Evolutionary Genetics Analysis across Computing Platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef]
- Suchard, M.A.; Lemey, P.; Baele, G.; Ayres, D.L.; Drummond, A.J.; Rambaut, A. Bayesian phylogenetic and phylodynamic data integration using BEAST 1.10. Virus Evol. 2018, 4, vey016. [Google Scholar] [CrossRef] [Green Version]
- Rio de Janeiro Government State. Foundation State Center for Statistics, Research and Training of Public Servants in Rio de Janeiro (CEPERJ Foundation). Ter-ritory Information. Fluminense Cartography. 2019. Available online: http://www.ceperj.rj.gov.br/Conteudo.asp?ident=79 (accessed on 20 December 2020).
- Rio de Janeiro Government State; Rio de Janeiro Municipal Government. Pereira Passos Municipal Urbanism Institute (IPP). Geo Portal. 2017. Available online: http://www.rio.rj.gov.br/web/ipp/exibeconteudo?id=4477436 (accessed on 20 December 2020).
- Federal Government of Brazil; Brazilian Institute of Geography and Statistics (IBGE). Geocentric Reference System for the Americas (SIRGAS 2000). Available online: https://www.ibge.gov.br/geociencias/informacoes-sobre-posicionamento-geodesico/sirgas.html (accessed on 15 January 2021).
- Brazilian Micro and Small Business Support Service. Rio de Janeiro City Regional Panel. 2016. Available online: https://www.sebrae.com.br/Sebrae/Portal%20Sebrae/UFs/RJ/Anexos/SebraePainel_CidadeRiodeJaneiro.pdf (accessed on 5 March 2020).
- Brazilian Ministry of Health. Viral Hepatitis Epidemiological Bulletin. 2021. Available online: http://www.aids.gov.br/pt-br/pub/2021/boletim-epidemiologico-hepatites-virais-2021 (accessed on 20 December 2021).
- Lopes, G.; Amorim, V.; Cavallier, F. Favelas Cariocas: Comparação das Áreas Ocupadas 2004–2011. Rio de Janeiro Municipal Government. Pereira Passos Municipal Urbanism Institute (IPP). Geo Portal. 2012. Available online: https://www.data.rio/datasets/b99b4d29ab5344338ad6c3e7b1c1cc97 (accessed on 20 December 2020).
- Brazilian Government; Institute for Applied Economic Research; João Pimheiro Fundation. Atlas do Desenvolvimento Humano nas Regiões Metropolitanas Brasileiras. 2014. Available online: https://secid.ma.gov.br/files/2015/04/Atlas-do-Desenvolvimento-Humanonas-Regi%C3%B5es-Metropolitanas.pdf (accessed on 20 December 2020).
- Caballero, B. Índice de Desenvolvimento Humano Municipal: Análise para a Cidade do Rio de Janeiro. Rio de Janeiro Municipal Government. Pereira Passos Municipal Urbanism Institute (IPP). Geo Portal. 2015. Available online: https://www.data.rio/datasets/53d6f84a8b514efebf17e137a7d028ad (accessed on 20 December 2020).
- World Health Organization. Public Health for Mass Gatherings: Key Considerations. 2016. Available online: https://www.who.int/ihr/publications/WHO_HSE_GCR_2015.5/en/ (accessed on 20 November 2018).
- Marciano, S.; Arufe, D.; Haddad, L.; Mendizabal, M.; Gadano, A.; Gaite, L.; Anders, M.; Garrido, L.; Martinez, A.A.; Conte, D.; et al. Outbreak of hepatitis A in a post-vaccination era: High rate of co-infection with sexually transmitted diseases. Ann. Hepatol. 2020, 19, 641–644. [Google Scholar] [CrossRef]
- Ibáñez-Tomás, E.; Gasch-Gallén, À. Sexual practices and the risk of Hepatitis A in men who have sex with men in Spain. J. Nurs. Manag. 2021, 29, 32–42. [Google Scholar] [CrossRef]
- Henning, K.J.; Bell, E.; Braun, J.; Barker, N.D. A community-wide outbreak of hepatitis A: Risk factors for infection among homosexual and bisexual men. Am. J. Med. 1995, 99, 132–136. [Google Scholar] [CrossRef] [Green Version]
- Rodríguez-Tajes, S.; Perpiñán, E.; Caballol, B.; Lens, S.; Mariño, Z.; Costa, J.; Vilella, A.; Pérez-Del-Pulgar, S.; Forns, X.; Koutsoudakis, G. Hepatitis A outbreak in Barcelona among men who have sex with men (MSM), January–June 2017: A hospital perspective. Liver Int. 2018, 38, 588–593. [Google Scholar] [CrossRef]
- Bura, M.; Bura, A.; Mozer-Lisewska, I. Syphilis and HIV infection in patients with hepatitis A: A preliminary study from one centre in Poland. Postepy Dermatol. Alergol. 2020, 37, 810–811. [Google Scholar] [CrossRef] [PubMed]
- Castro, L.S.; de Rezende, G.R.; Fernandes, F.R.P.; Bandeira, L.M.; Cesar, G.A.; Lago, B.V.D.; Gouvêa, M.S.G.; Motta-Castro, A.R.C. HAV infection in Brazilian men who have sex with men: The importance of surveillance to avoid outbreaks. PLoS ONE 2021, 16, e0256818. [Google Scholar] [CrossRef] [PubMed]
- Chuffi, S.; Gomes-Gouvêa, M.S.; Casadio, L.V.B.; Nastri, A.C.S.S.; Gonzalez, M.P.; Cotia, A.L.F.; Aranda, A.G.D.; Tenore, S.B.; Ono, S.K.; Malta, F.M.; et al. The Molecular Characterization of Hepatitis A Virus Strains Circulating during Hepatitis A Outbreaks in São Paulo, Brazil, from September 2017 to May 2019. Viruses 2022, 14, 73. [Google Scholar] [CrossRef] [PubMed]
- Breyer, B.N.; Smith, J.F.; Eisenberg, M.L.; Ando, K.A.; Rowen, T.S.; Shindel, A.W. The impact of sexual orientation on sexuality and sexual practices in North American medical students. J. Sex. Med. 2010, 7, 2391–2400. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goethe, V.E.; Angerer, H.; Dinkel, A.; Arsov, C.; Hadaschik, B.; Imkamp, F.; Gschwend, J.E.; Herkommer, K. Concordance and Discordance of Sexual Identity, Sexual Experience, and Current Sexual Behavior in 45-Year-Old Men: Results from the German Male Sex-Study. Sex. Med. 2018, 6, 282–290. [Google Scholar] [CrossRef] [PubMed]
- De Paula, V.S.; Lu, L.; Niel, C.; Gaspar, A.M.; Robertson, B.H. Genetic analysis of hepatitis A virus isolates from Brazil. J. Med. Virol. 2004, 73, 378–383. [Google Scholar] [CrossRef]
- Prado, T.; Barbosa, M.R.F.; Araújo, R.S.; Garcia, S.C.; Melo, A.J.; Galvani, A.T.; Brandão, C.J.; Silva, R.L.O.; Sato, M.I.Z. Hepatitis A Outbreaks and Environmental Circulation of Genotype IA Strains in the São Paulo City, 2017–2018. Food Environ. Virol. 2021, 13, 520–527. [Google Scholar] [CrossRef]
- Cella, E.; Riva, E.; Angeletti, S.; Fogolari, M.; Blasi, A.; Bsc, V.S.; Spoto, S.; Bazzardi, R.; Lai, A.; Sagnelli, C.; et al. Genotype I hepatitis A virus introduction in Italy: Bayesian phylogenetic analysis to date different epidemics. J. Med. Virol. 2018, 90, 1493–1502. [Google Scholar] [CrossRef]
- Amon, J.; Devasia, R.; Xia, G.; Nainan, O.V.; Hall, S.; Lawson, B.; Wolthuis, J.S.; Macdonald, P.D.M.; Shepard, C.W.; Williams, I.T.; et al. Molecular epidemiology of foodborne hepatitis a outbreaks in the United States, 2003. J. Infect. Dis. 2005, 192, 1323–1330. [Google Scholar] [CrossRef] [Green Version]
- Wang, H.; Wang, X.Y.; Zheng, H.H.; Cao, J.Y.; Zhou, W.T.; Bi, S.L. Evolution and genetic characterization of hepatitis A virus isolates in China. Int. J. Infect. Dis. 2015, 33, 156–158. [Google Scholar] [CrossRef] [Green Version]
- Lanini, S.; Minosse, C.; Vairo, F.; Garbuglia, A.; Di Bari, V.; Agresta, A.; Rezza, G.; Puro, V.; Pendenza, A.; Loffredo, M.R.; et al. A large ongoing outbreak of hepatitis A predominantly affecting young males in Lazio, Italy; August 2016–March 2017. PLoS ONE 2017, 12, e0185428. [Google Scholar] [CrossRef] [Green Version]
- Beebeejaun, K.; Degala, S.; Balogun, K.; Simms, I.; Woodhall, S.C.; Heinsbroek, E.; Crook, P.; Kar-Purkayastha, I.; Treacy, J.; Wedgwood, K.; et al. Outbreak of hepatitis A associated with men who have sex with men (MSM), England, July 2016 to January 2017. Euro Surveill. 2017, 22, 30454. [Google Scholar] [CrossRef]
- Stockholm. Hepatitis A Outbreaks in the EU/EEA Mostly Affecting Men Who Have Sex with Men—Second Update, 19 May 2017; ECDC: Stockholm, Sweden, 2017. Available online: https://www.ecdc.europa.eu/sites/default/files/documents/RRA-19-May-2017_UPDATE_2-HepatitisA-in-mostly-MSM.pdf (accessed on 20 November 2018).







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variables | Total (n = 139) | Hepatitis A (Case Group) (n = 52) | Control Group (n = 87) | p-Value * | |||
|---|---|---|---|---|---|---|---|
| n | (%) | n | (%) | n | (%) | ||
| Biological Sex | 0.024 | ||||||
| Feminine | 47 | (33.81) | 11 | (21.15) | 36 | (41.38) | |
| Masculine | 92 | (66.19) | 41 | (78.85) | 51 | (58.62) | |
| Sexual Orientation | 0.002 | ||||||
| Heterosexual Women | 41 | (29.5) | 8 | (15.38) | 33 | (37.93) | |
| WSW 1 | 6 | (4.32) | 3 | (5.77) | 3 | (3.45) | |
| Heterosexual Men | 38 | (27.34) | 11 | (21.15) | 27 | (31.03) | |
| MSM 2 | 54 | (38.85) | 30 | (57.69) | 24 | (27.59) | |
| Age Groups | <0.001 | ||||||
| 20 to 29 years | 32 | (23.02) | 24 | (46.15) | 8 | (9.2) | |
| 30 to 39 years | 42 | (30.22) | 20 | (38.46) | 22 | (25.29) | |
| 40 to 59 years | 36 | (25.9) | 7 | (13.46) | 29 | (33.33) | |
| 60 years or more | 29 | (20.86) | 1 | (1.93) | 28 | (32.18) | |
| Mean age | 40.03 ± 12.97 (20 to 77 years) | - | 32.19 ± 10.54 (20 to 73 years) | - | 44.71 ± 12.03 (22 to 77 years) | - | <0.001 |
| Planning Area 3 | 0.001 | ||||||
| North † | 2.23 | (42.45) | 1.17 | (59.62) | 1.06 | (32.18) | |
| South † | 0.69 | (6.47) | 0.08 | (1.92) | 0.6 2 | (9.2) | |
| West † | 1.72 | (28.78) | 0.34 | (15.38) | 1.38 | (36.78) | |
| Central † | 0.85 | (7.91) | 0.54 | (13.46) | 0.31 | (4.6) | |
| Neighboring cities a | 0.75 | (14.39) | 0.18 | (9.62) | 0.56 | (17.24) | |
| HAV Exposure Categories | Total (n = 139) † | Hepatitis A (Case Group) (n = 52) † | Control Group (n = 87) † | p-Value * | Unadjusted | Adjusted | |||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| n | (%) | n | (%) | n | (%) | OR (95% CI) | p-Value ** | OR (95% CI) | p-Value ** | ||
| Parenteral factor | 0.056 | ||||||||||
| Yes | 63 | (45.99) | 18 | (34.62) | 45 | (52.94) | 0.42 (0.17;1.02) | 0.1254 | 0.46 (0.18; 1.17) | 0.1076 | |
| No | 74 | (54.01) | 34 | (65.38) | 40 | (47.06) | 1.00 | 1.00 | |||
| Injectable Remedies | 0.818 | ||||||||||
| Yes | 21 | (15.44) | 9 | (17.31) | 12 | (14.29) | - | - | - | - | |
| No | 115 | (84.56) | 43 | (82.69) | 72 | (85.71) | - | - | - | - | |
| Inhalable Drugs | 1.000 | ||||||||||
| Yes | 14 | (10.29) | 5 | (9.62) | 9 | (10.71) | - | - | - | - | |
| No | 122 | (89.71) | 47 | (90.38) | 75 | (89.29) | - | - | - | - | |
| Injectable Drugs | 0.698 | ||||||||||
| Yes | 2 | (1.47) | 0 | (0) | 2 | (2.38) | - | - | - | - | |
| No | 134 | (98.53) | 52 | (100) | 82 | (97.62) | - | - | - | - | |
| Tattoos | 0.751 | ||||||||||
| Yes | 13 | (9.56) | 6 | (11.54) | 7 | (8.33) | - | - | - | - | |
| No | 123 | (90.44) | 46 | (88.46) | 77 | (91.67) | - | - | - | - | |
| Piercings | 0.662 | ||||||||||
| Yes | 10 | (7.41) | 5 | (9.62) | 5 | (6.02) | - | - | - | - | |
| No | 125 | (92.59) | 47 | (90.38) | 78 | (93.98) | - | - | - | - | |
| Dental treatment | 1.000 | ||||||||||
| Yes | 24 | (17.65) | 9 | (17.31) | 15 | (17.86) | - | - | - | - | |
| No | 112 | (82.35) | 43 | (82.69) | 69 | (82.14) | - | - | - | - | |
| Blood transfusion | 0.063 | ||||||||||
| Yes | 8 | (5.93) | 0 | 0 | 8 | (9.41) | - | - | - | - | |
| No | 127 | (94.07) | 50 | (100) | 77 | (90.59) | - | - | - | - | |
| Food-borne factors | 0.101 | ||||||||||
| Yes | 38 | (34.86) | 22 | (44) | 16 | (27.12) | 1.52 (0.62–3.82) | 0.2891 | 1.71 (0.62–4.82) | 0.2992 | |
| No | 71 | (65.14) | 28 | (56) | 43 | (72.88) | 1.00 | 1.00 | |||
| Raw meat consumption | 0.632 | ||||||||||
| Yes | 29 | (27.36) | 15 | (30.61) | 14 | (24.56) | - | - | - | - | |
| No | 77 | (72.64) | 34 | (69.39) | 43 | (75.44) | - | - | - | - | |
| Raw vegetable consumption | 0.018 * | ||||||||||
| Yes | 9 | (9) | 8 | (17.39) | 1 | (1.85) | - | - | - | - | |
| No | 91 | (91) | 38 | (82.61) | 53 | (98.15) | - | - | - | - | |
| Well water and/or cistern water consumption | 0.405 | ||||||||||
| Yes | 6 | (7.23) | 4 | (11.43) | 2 | (4.17) | - | - | - | - | |
| No | 77 | (92.77) | 31 | (88.57) | 46 | (95.83) | - | - | - | - | |
| Water-borne factors | 0.410 | ||||||||||
| Yes | 40 | (37.38) | 22 | (42.31) | 18 | (32.73) | 1.53 (0.64–3.70) | 0.9041 | 0.97 (0.34–2.62) | 0.9464 | |
| No | 67 | (62.62) | 30 | (57.69) | 37 | (67.27) | 1.00 | 1.00 | |||
| Floodwater exposure | 0.786 | ||||||||||
| Yes | 12 | (11.88) | 7 | (13.73) | 5 | (10) | - | - | - | - | |
| No | 89 | (88.12) | 44 | (86.27) | 45 | (90) | - | - | - | - | |
| Beach bathing | 1.000 | ||||||||||
| Yes | 19 | (18.27) | 10 | (19.23) | 9 | (17.31) | - | - | - | - | |
| No | 85 | (81.73) | 42 | (80.77) | 43 | (82.69) | - | - | - | - | |
| River and/or lake bathing | 1.000 | ||||||||||
| Yes | 8 | (7.77) | 4 | (7.69) | 4 | (7.84) | - | - | - | - | |
| No | 95 | (92.23) | 48 | (92.31) | 47 | (92.16) | - | - | - | - | |
| Well water supply | 0.263 | ||||||||||
| Yes | 3 | (3.03) | 3 | (5.88) | 0 | (0) | - | - | - | - | |
| No | 96 | (96.97) | 48 | (94.12) | 48 | (100) | - | - | - | - | |
| Cistern water supply | 0.493 | ||||||||||
| Yes | 2 | (2.08) | 2 | (4.08) | 0 | (0) | - | - | - | - | |
| No | 94 | (97.92) | 47 | (95.92) | 47 | (100) | - | - | - | - | |
| Piped water supply | 1.000 | ||||||||||
| Yes | 89 | (92.71) | 46 | (92) | 43 | (93.48) | - | - | - | - | |
| No | 7 | (7.29) | 4 | (8) | 3 | (6.52) | - | - | - | - | |
| Sewage system | 0.752 | ||||||||||
| Yes | 90 | (93.75) | 46 | (92) | 44 | (95.65) | - | - | - | - | |
| No | 6 | (6.25) | 4 | (8) | 2 | (4.35) | - | - | - | - | |
| Sexual Factors | 0.062 | ||||||||||
| Yes | 114 | (88.37) | 48 | (96) | 66 | (83.54) | 1.81 (0.33–13.59) | 0.9178 | 0.88 (0.15–6.99) | 0.8902 | |
| No | 15 | (11.63) | 2 | (4) | 13 | (16.46) | 1.00 | 1.00 | |||
| Oral Sex | 0.167 | ||||||||||
| Yes | 72 | (81.82) | 39 | (88.64) | 33 | (75) | - | - | - | - | |
| No | 16 | (18.18) | 5 | (11.36) | 11 | (25) | - | - | - | - | |
| Oroanal sex | 0.826 | ||||||||||
| Yes | 32 | (46.38) | 19 | (44.19) | 13 | (50) | - | - | - | - | |
| No | 37 | (53.62) | 24 | (55.81) | 13 | (50) | - | - | - | - | |
| Anal Sex | 0.323 | ||||||||||
| Yes | 58 | (68.24) | 34 | (73.91) | 24 | (61.54) | - | - | - | - | |
| No | 27 | (31.76) | 12 | (26.09) | 15 | (38.46) | - | - | - | - | |
| ≥3 sexual partners | 0.004 * | ||||||||||
| Yes | 38 | (42.22) | 11 | (25.58) | 27 | (57.45) | - | - | - | - | |
| No | 52 | (57.78) | 32 | (74.42) | 20 | (42.55) | - | - | - | - | |
| Vaginal Sex | 0.082 | ||||||||||
| Yes | 60 | (82.19) | 16 | (100) | 44 | (77.19) | - | - | - | - | |
| No | 13 | (17.81) | 0 | (0) | 13 | (22.81) | - | - | - | - | |
| Unprotected sex | 0.980 | ||||||||||
| Yes | 89 | (79.46) | 36 | (78.26) | 53 | (80.3) | - | - | - | - | |
| No | 23 | (20.54) | 10 | (21.74) | 13 | (19.7) | - | - | - | - | |
| Travel to endemic region | 0.001 * | ||||||||||
| Yes | 19 | (18.81) | 15 | (34.09) | 4 | (7.02) | 6.90 (2.01–32.16) | <0.01 ** | 7.19 (1.93–36.04) | <0.01 ** | |
| No | 82 | (81.19) | 29 | (65.91) | 52 | (92.98) | 1.00 | 1.00 | |||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mello, V.M.; Bianchi, L.M.; Sousa, P.S.F.; Tavares, P.S.; Di Salvo, D.R.G.; Ginuino, C.F.; Almeida, N.A.A.; Fernandes, C.A.S.; Mello, F.C.A.; Villar, L.M.; et al. Increase in Hepatitis A Cases Linked to Imported Strains to Rio de Janeiro, Brazil: A Cross-Sectional Study. Viruses 2022, 14, 303. https://doi.org/10.3390/v14020303
Mello VM, Bianchi LM, Sousa PSF, Tavares PS, Di Salvo DRG, Ginuino CF, Almeida NAA, Fernandes CAS, Mello FCA, Villar LM, et al. Increase in Hepatitis A Cases Linked to Imported Strains to Rio de Janeiro, Brazil: A Cross-Sectional Study. Viruses. 2022; 14(2):303. https://doi.org/10.3390/v14020303
Chicago/Turabian StyleMello, Vinicius M., Lucas M. Bianchi, Paulo Sergio F. Sousa, Pedro S. Tavares, Daniel R. G. Di Salvo, Cleber F. Ginuino, Nathalia A. A. Almeida, Carlos A. S. Fernandes, Francisco C. A. Mello, Livia M. Villar, and et al. 2022. "Increase in Hepatitis A Cases Linked to Imported Strains to Rio de Janeiro, Brazil: A Cross-Sectional Study" Viruses 14, no. 2: 303. https://doi.org/10.3390/v14020303