
Validation of Viral Inactivation Protocols for Therapeutic Blood Products against Severe Acute Respiratory Syndrome Coronavirus-2 (SARS-CoV-2)
 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Reagents and Chemicals
2.2. Cell Culture
2.3. Normal Plasma and Convalescent Serum Preparation
2.4. Preparation of SARS-CoV-2 Pseudoparticles (SARS-CoV-2pp)
2.5. S/D Inactivation Protocols of the SARS-CoV-2pp-Spiked FFP
2.6. Cytotoxicity Assay
2.7. Infectivity Assay
2.8. Effect of S/D Inactivation Protocols on the Neutralizing Activity of CCS
2.9. Statistical Analysis
3. Results
3.1. Production of SARS-CoV-2pp
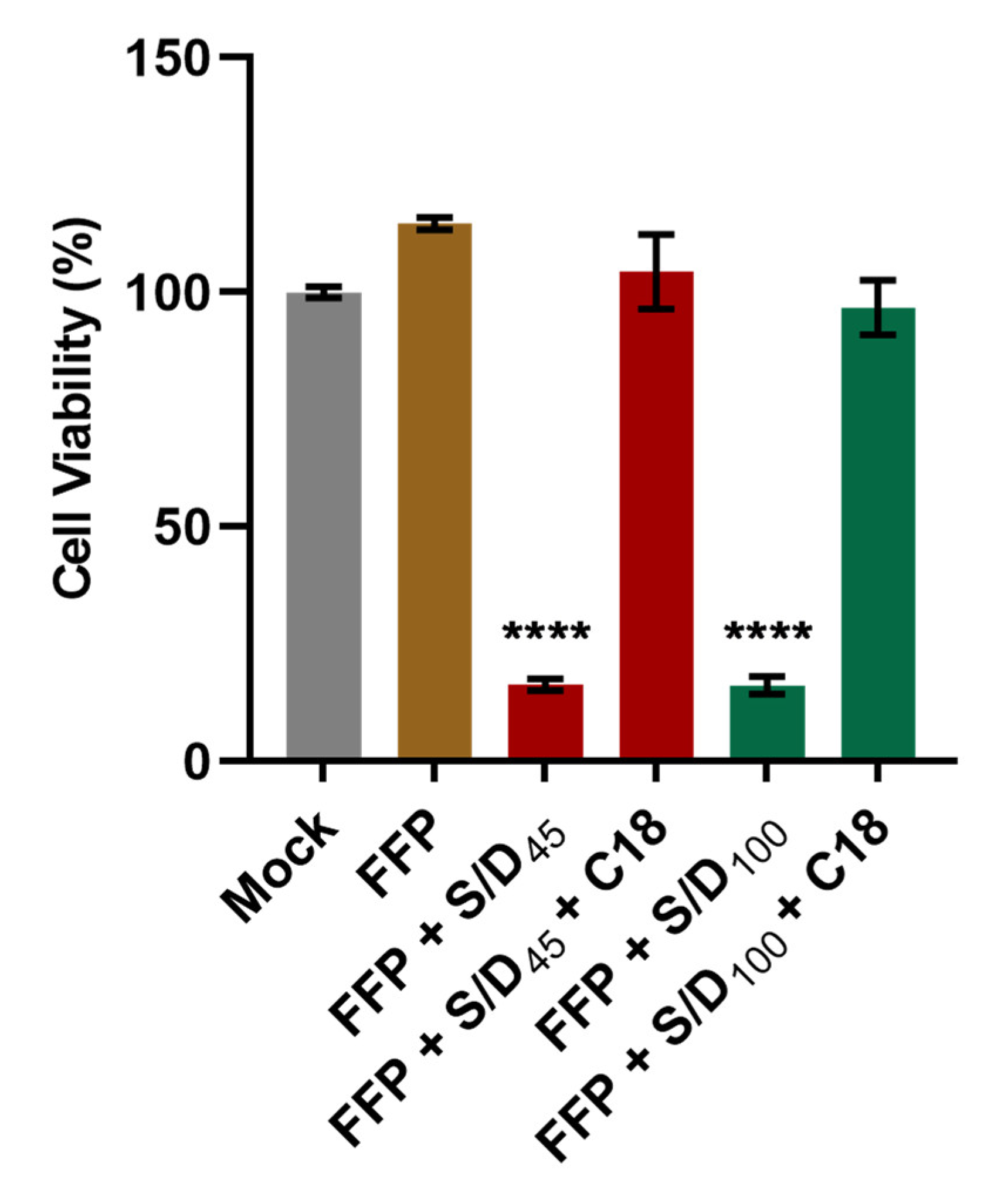
3.2. Elimination of S/D Cytotoxicity by C18 Filtration in S/D-Treated Plasma
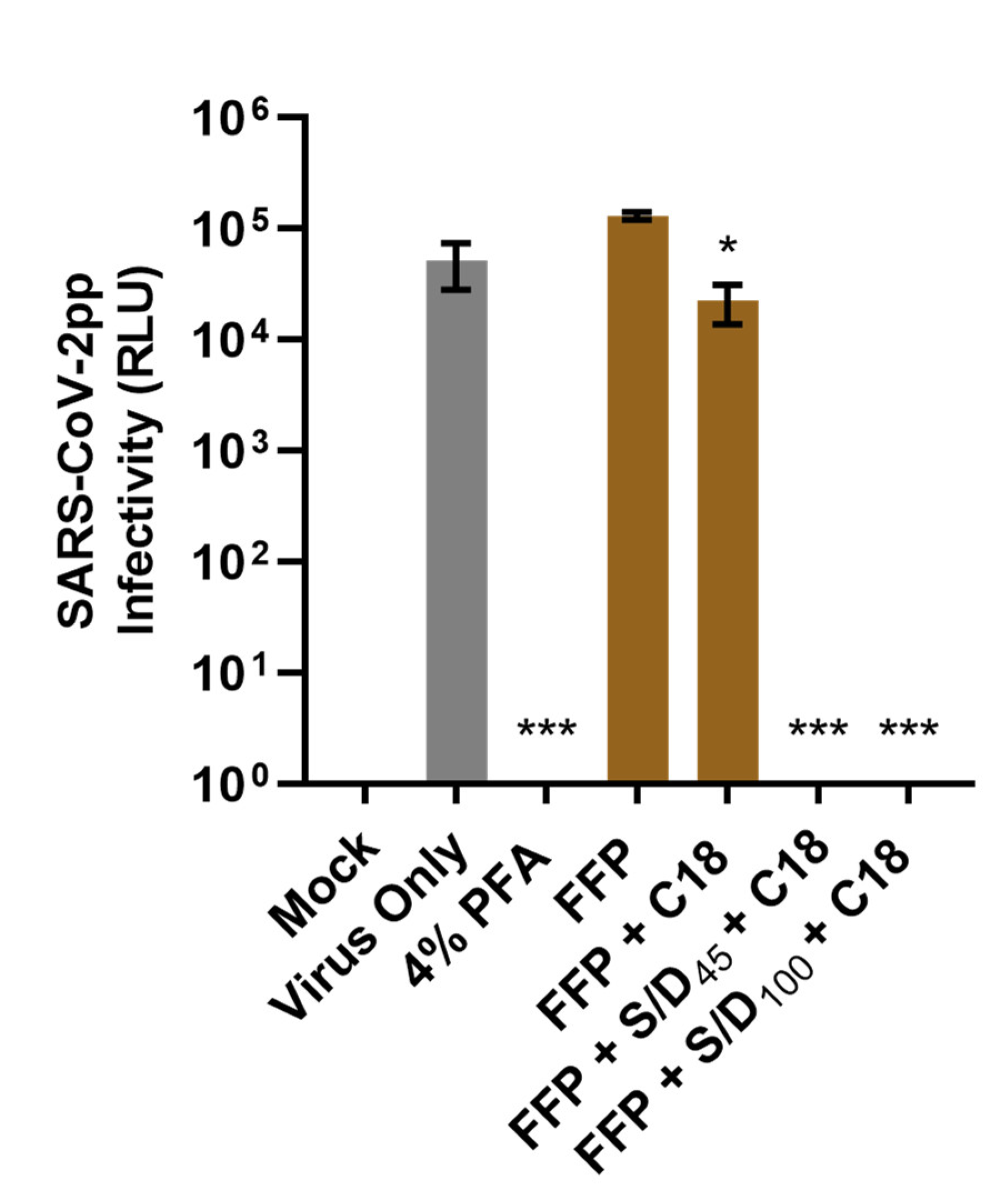
3.3. S/D Inactivation Protocols Efficiently Eliminate SARS-CoV-2pp Infectivity in Human Plasma
3.4. The Neutralizing Activity of COVID-19 Convalescent Serum Is Preserved Following the S/D Inactivation Protocols
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Astuti, I.; Ysrafil. Severe Acute Respiratory Syndrome Coronavirus 2 (SARS-CoV-2): An overview of viral structure and host response. Syndr. Clin. Res. Rev. 2020, 14, 407–412. [Google Scholar] [CrossRef] [PubMed]
- Center for Systems Science and Engineering at Johns Hopkins University. COVID-19 Dashboard. Available online: https://coronavirus.jhu.edu/map.html (accessed on 1 August 2022).
- Hadi, J.; Dunowska, M.; Wu, S.; Brightwell, G. Control Measures for SARS-CoV-2: A Review on Light-Based Inactivation of Single-Stranded RNA Viruses. Pathogens 2020, 9, 737. [Google Scholar] [CrossRef] [PubMed]
- Jiang, S.; Hillyer, C.; Du, L. Neutralizing Antibodies against SARS-CoV-2 and Other Human Coronaviruses. Trends Immunol. 2020, 41, 355–359. [Google Scholar] [CrossRef] [PubMed]
- Salazar, E.; Kuchipudi, S.V.; Christensen, P.A.; Eagar, T.; Yi, X.; Zhao, P.; Jin, Z.; Long, S.W.; Olsen, R.J.; Chen, J.; et al. Convalescent plasma anti-SARS-CoV-2 spike protein ectodomain and receptor-binding domain IgG correlate with virus neutralization. J. Clin. Investig. 2020, 130, 6728–6738. [Google Scholar] [CrossRef] [PubMed]
- Du, L.; Yang, Y.; Zhang, X. Neutralizing antibodies for the prevention and treatment of COVID-19. Cell Mol. Immunol. 2021, 18, 2293–2306. [Google Scholar] [CrossRef]
- Harvey, W.T.; Carabelli, A.M.; Jackson, B.; Gupta, R.K.; Thomson, E.C.; Harrison, E.M.; Ludden, C.; Reeve, R.; Rambaut, A.; Consortium, C.-G.U.; et al. SARS-CoV-2 variants, spike mutations and immune escape. Nat. Rev. Microbiol. 2021, 19, 409–424. [Google Scholar] [CrossRef]
- Wang, P.; Liu, L.; Nair, M.S.; Yin, M.T.; Luo, Y.; Wang, Q.; Yuan, T.; Mori, K.; Solis, A.G.; Yamashita, M.; et al. SARS-CoV-2 neutralizing antibody responses are more robust in patients with severe disease. Emerg. Microbes Infect. 2020, 9, 2091–2093. [Google Scholar] [CrossRef]
- Beaudoin-Bussières, G.; Laumaea, A.; Anand, S.P.; Prévost, J.; Gasser, R.; Goyette, G.; Medjahed, H.; Perreault, J.; Tremblay, T.; Lewin, A.; et al. Decline of humoral responses against SARS-CoV-2 Spike in convalescent individuals. mBio 2020, 11, e02590-20. [Google Scholar] [CrossRef]
- Suthar, M.S.; Zimmerman, M.; Kauffman, R.; Mantus, G.; Linderman, S.; Vanderheiden, A.; Nyhoff, L.; Davis, C.; Adekunle, S.; Affer, M.; et al. Rapid generation of neutralizing antibody responses in COVID-19 patients. Cell Rep. Med. 2020, 1, 100040. [Google Scholar] [CrossRef]
- Maor, Y.; Cohen, D.; Paran, N.; Israely, T.; Ezra, V.; Axelrod, O.; Shinar, E.; Izak, M.; Rahav, G.; Rahimi-Levene, N.; et al. Compassionate use of convalescent plasma for treatment of moderate and severe pneumonia in COVID-19 patients and association with IgG antibody levels in donated plasma. EClinicalMedicine 2020, 26, 100525. [Google Scholar] [CrossRef]
- Burnouf, T.; Gathof, B.; Bloch, E.M.; Bazin, R.; de Angelis, V.; Patidar, G.K.; Rastvorceva, R.M.G.; Oreh, A.; Goel, R.; Rahimi-Levene, N.; et al. Production and Quality Assurance of Human Polyclonal Hyperimmune Immunoglobulins Against SARS-CoV-2. Transfus. Med. Rev. 2022, 36, 125–132. [Google Scholar] [CrossRef] [PubMed]
- Perricone, C.; Triggianese, P.; Bursi, R.; Cafaro, G.; Bartoloni, E.; Chimenti, M.S.; Gerli, R.; Perricone, R. Intravenous Immunoglobulins at the Crossroad of Autoimmunity and Viral Infections. Microorganisms 2021, 9, 121. [Google Scholar] [CrossRef] [PubMed]
- Bloch, E.M.; Shoham, S.; Casadevall, A.; Sachais, B.S.; Shaz, B.; Winters, J.L.; van Buskirk, C.; Grossman, B.J.; Joyner, M.; Henderson, J.P.; et al. Deployment of convalescent plasma for the prevention and treatment of COVID-19. J. Clin. Investig. 2020, 130, 2757–2765. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Piechotta, V.; Iannizzi, C.; Chai, K.L.; Valk, S.J.; Kimber, C.; Dorando, E.; Monsef, I.; Wood, E.M.; Lamikanra, A.A.; Roberts, D.J.; et al. Convalescent plasma or hyperimmune immunoglobulin for people with COVID-19: A living systematic review. Cochrane Database Syst. Rev. 2021, 5, CD013600. [Google Scholar] [CrossRef]
- Valk, S.J.; Piechotta, V.; Chai, K.L.; Doree, C.; Monsef, I.; Wood, E.M.; Lamikanra, A.; Kimber, C.; McQuilten, Z.; So-Osman, C.; et al. Convalescent plasma or hyperimmune immunoglobulin for people with COVID-19: A rapid review. Cochrane Database Syst. Rev. 2020, 5, CD013600. [Google Scholar] [CrossRef]
- Bar, K.J.; Shaw, P.A.; Choi, G.H.; Aqui, N.; Fesnak, A.; Yang, J.B.; Soto-Calderon, H.; Grajales, L.; Starr, J.; Andronov, M.; et al. A randomized controlled study of convalescent plasma for individuals hospitalized with COVID-19 pneumonia. J. Clin. Investig. 2021, 131, e155114. [Google Scholar] [CrossRef]
- Hirsch, C.; Park, Y.S.; Piechotta, V.; Chai, K.L.; Estcourt, L.J.; Monsef, I.; Salomon, S.; Wood, E.M.; So-Osman, C.; McQuilten, Z. SARS-CoV-2-neutralising monoclonal antibodies to prevent COVID-19. Cochrane Database Syst. Rev. 2022, 6, CD014945. [Google Scholar]
- Recovery Collaborative Group. Convalescent plasma in patients admitted to hospital with COVID-19 (RECOVERY): A randomised controlled, open-label, platform trial. Lancet 2021, 397, 2049–2059. [Google Scholar] [CrossRef]
- Focosi, D.; Franchini, M.; Pirofski, L.A.; Burnouf, T.; Paneth, N.; Joyner, M.J.; Casadevall, A. COVID-19 Convalescent Plasma and Clinical Trials: Understanding Conflicting Outcomes. Clin. Microbiol. Rev. 2022, 35, e0020021. [Google Scholar] [CrossRef]
- Valdez-Cruz, N.A.; Garcia-Hernandez, E.; Espitia, C.; Cobos-Marin, L.; Altamirano, C.; Bando-Campos, C.G.; Cofas-Vargas, L.F.; Coronado-Aceves, E.W.; Gonzalez-Hernandez, R.A.; Hernandez-Peralta, P.; et al. Integrative overview of antibodies against SARS-CoV-2 and their possible applications in COVID-19 prophylaxis and treatment. Microb. Cell Fact. 2021, 20, 88. [Google Scholar] [CrossRef]
- Oliveira, F.A.; Nucci, M.P.; Rego, G.N.A.; Alves, A.D.H.; Marti, L.C.; Nucci, L.P.; Mamani, J.B.; Gamarra, L.F. Convalescent plasma therapy in COVID-19 critically ill patients during advanced phases of clinical trials and their preliminary results. Einstein 2021, 19, eRW6186. [Google Scholar] [CrossRef] [PubMed]
- Dulipsingh, L.; Ibrahim, D.; Schaefer, E.J.; Crowell, R.; Diffenderfer, M.R.; Williams, K.; Lima, C.; McKenzie, J.; Cook, L.; Puff, J.; et al. SARS-CoV-2 serology and virology trends in donors and recipients of convalescent plasma. Transfus. Apher. Sci. 2020, 59, 102922. [Google Scholar] [CrossRef] [PubMed]
- Mosnier, L.O. How much clotting is enough? Blood 2020, 136, 651–652. [Google Scholar] [CrossRef]
- Zeng, Q.L.; Yu, Z.J.; Gou, J.J.; Li, G.M.; Ma, S.H.; Zhang, G.F.; Xu, J.H.; Lin, W.B.; Cui, G.L.; Zhang, M.M.; et al. Effect of Convalescent Plasma Therapy on Viral Shedding and Survival in Patients With Coronavirus Disease 2019. J. Infect. Dis. 2020, 222, 38–43. [Google Scholar] [CrossRef] [PubMed]
- Selvi, V. Convalescent Plasma: A Challenging Tool to Treat COVID-19 Patients-A Lesson from the Past and New Perspectives. Biomed. Res. Int. 2020, 2020, 2606058. [Google Scholar] [CrossRef] [PubMed]
- Boyarsky, B.J.; Werbel, W.A.; Avery, R.K.; Tobian, A.A.R.; Massie, A.B.; Segev, D.L.; Garonzik-Wang, J.M. Antibody Response to 2-Dose SARS-CoV-2 mRNA Vaccine Series in Solid Organ Transplant Recipients. JAMA 2021, 325, 2204–2206. [Google Scholar] [CrossRef] [PubMed]
- Abdelhady, H.; Abdelrazik, M.; Abdi, Z.; Abdo, D.; Abdulle, A.; Abel, L.; Abouzeenni, S.; Abrahamson, G.; Abusamra, Y.; Adams, L.; et al. Effect of Convalescent Plasma on Organ Support-Free Days in Critically Ill Patients With COVID-19: A Randomized Clinical Trial. JAMA 2021, 326, 1690–1702. [Google Scholar] [CrossRef]
- Gharbharan, A.; Jordans, C.C.E.; GeurtsvanKessel, C.; den Hollander, J.G.; Karim, F.; Mollema, F.P.N.; Stalenhoef-Schukken, J.E.; Dofferhoff, A.; Ludwig, I.; Koster, A.; et al. Effects of potent neutralizing antibodies from convalescent plasma in patients hospitalized for severe SARS-CoV-2 infection. Nat. Commun. 2021, 12, 3189. [Google Scholar] [CrossRef]
- Expert Panel Updates Guidelines for Clinical CCP Use. Available online: https://www.aabb.org/news-resources/news/article/2022/04/13/aabb-expert-panel-updates-guidelines-for-clinical-ccp-use (accessed on 27 July 2022).
- Lacombe, K.; Hueso, T.; Porcher, R.; Mekinian, A.; Chiarabini, T.; Georgin-Lavialle, S.; Ader, F.; Saison, J.; Martin Blondel, G.; de Castro, N.; et al. COVID-19 convalescent plasma for hospitalizedCOVID-19 patients not requiring assisted ventilation. A randomized controlled trial. Vox Sang 2022, 117, 74–75. [Google Scholar]
- FDA. Regulatory Update: FDA Issues Revised CCP Guidance, AABB Updates CCP Toolkit. Available online: https://www.aabb.org/news-resources/news/article/2022/01/11/regulatory-update-fda-issues-revised-ccp-guidance-aabb-updates-ccp-toolkit (accessed on 27 July 2022).
- Larkey, N.E.; Ewaisha, R.; Lasho, M.A.; Roforth, M.M.; Granger, D.; Jerde, C.R.; Wu, L.; Gorsh, A.; Klassen, S.A.; Senefeld, J.W.; et al. Limited Correlation between SARS-CoV-2 Serologic Assays for Identification of High-Titer COVID-19 Convalescent Plasma Using FDA Thresholds. Microbiol. Spectr. 2022, 10, e0115422. [Google Scholar] [CrossRef]
- US Food and Drugs Administration. Convalescent Plasma EUA Letter of Authorization 12282021. Available online: https://www.fda.gov/media/141477/download (accessed on 25 September 2022).
- Chang, L.; Yan, Y.; Wang, L. Coronavirus Disease 2019: Coronaviruses and Blood Safety. Transfus. Med. Rev. 2020, 34, 75–80. [Google Scholar] [CrossRef] [PubMed]
- AlShehry, N.; Zaidi, S.Z.A.; AlAskar, A.; Al Odayani, A.; Alotaibi, J.M.; AlSagheir, A.; Al-Eyadhy, A.; Balelah, S.; Salam, A.; Zaidi, A.R.Z.; et al. Safety and Efficacy of Convalescent Plasma for Severe COVID-19: Interim Report of a Multicenter Phase II Study from Saudi Arabia. Saudi J. Med. Med. Sci. 2021, 9, 16–23. [Google Scholar] [CrossRef] [PubMed]
- Welch, S.R.; Davies, K.A.; Buczkowski, H.; Hettiarachchi, N.; Green, N.; Arnold, U.; Jones, M.; Hannah, M.J.; Evans, R.; Burton, C.; et al. Analysis of Inactivation of SARS-CoV-2 by Specimen Transport Media, Nucleic Acid Extraction Reagents, Detergents, and Fixatives. J. Clin. Microbiol. 2020, 58, e01713-20. [Google Scholar] [CrossRef] [PubMed]
- Patterson, E.I.; Prince, T.; Anderson, E.R.; Casas-Sanchez, A.; Smith, S.L.; Cansado-Utrilla, C.; Solomon, T.; Griffiths, M.J.; Acosta-Serrano, Á.; Turtle, L.; et al. Methods of Inactivation of SARS-CoV-2 for Downstream Biological Assays. J. Infect. Dis. 2020, 222, 1462–1467. [Google Scholar] [CrossRef]
- Liew, O.W.; Fanusi, F.; Ng, J.Y.X.; Ahidjo, B.A.; Ling, S.S.M.; Lilyanna, S.; Chong, J.P.C.; Lim, A.E.S.; Lim, W.Z.; Ravindran, S.; et al. Immunoassay-Compatible Inactivation of SARS-CoV-2 in Plasma Samples for Enhanced Handling Safety. ACS Omega 2022, 7, 25510–25520. [Google Scholar] [CrossRef] [PubMed]
- Kostin, A.I.; Lundgren, M.N.; Bulanov, A.Y.; Ladygina, E.A.; Chirkova, K.S.; Gintsburg, A.L.; Logunov, D.Y.; Dolzhikova, I.V.; Shcheblyakov, D.V.; Borovkova, N.V.; et al. Impact of pathogen reduction methods on immunological properties of the COVID-19 convalescent plasma. Vox Sang 2021, 116, 665–672. [Google Scholar] [CrossRef]
- Yonemura, S.; Hartson, L.; Dutt, T.S.; Henao-Tamayo, M.; Goodrich, R.; Marschner, S. Preservation of neutralizing antibody function in COVID-19 convalescent plasma treated using a riboflavin and ultraviolet light-based pathogen reduction technology. Vox Sang 2021, 116, 1076–1083. [Google Scholar] [CrossRef]
- Cagdas, D. Convalescent plasma and hyperimmune globulin therapy in COVID-19. Expert Rev. Clin. Immunol. 2021, 17, 309–316. [Google Scholar] [CrossRef]
- Joyner, M.J.; Wright, R.S.; Fairweather, D.; Senefeld, J.W.; Bruno, K.A.; Klassen, S.A.; Carter, R.E.; Klompas, A.M.; Wiggins, C.C.; Shepherd, J.R.; et al. Early safety indicators of COVID-19 convalescent plasma in 5000 patients. J. Clin. Investig. 2020, 130, 4791–4797. [Google Scholar] [CrossRef]
- Hellstern, P.; Solheim, B.G. The Use of Solvent/Detergent Treatment in Pathogen Reduction of Plasma. Transfus. Med. Hemother 2011, 38, 65–70. [Google Scholar] [CrossRef]
- Kuo, Y.T.; Liu, C.H.; Corona, A.; Fanunza, E.; Tramontano, E.; Lin, L.T. The Methanolic Extract of Perilla frutescens Robustly Restricts Ebola Virus Glycoprotein-Mediated Entry. Viruses 2021, 13, 1793. [Google Scholar] [CrossRef] [PubMed]
- Hung, T.C.; Jassey, A.; Liu, C.H.; Lin, C.J.; Lin, C.C.; Wong, S.H.; Wang, J.Y.; Yen, M.H.; Lin, L.T. Berberine inhibits hepatitis C virus entry by targeting the viral E2 glycoprotein. Phytomedicine 2019, 53, 62–69. [Google Scholar] [CrossRef] [PubMed]
- Connor, R.I.; Chen, B.K.; Choe, S.; Landau, N.R. Vpr is required for efficient replication of human immunodeficiency virus type-1 in mononuclear phagocytes. Virology 1995, 206, 935–944. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Giroglou, T.; Cinatl, J., Jr.; Rabenau, H.; Drosten, C.; Schwalbe, H.; Doerr, H.W.; von Laer, D. Retroviral vectors pseudotyped with severe acute respiratory syndrome coronavirus S protein. J. Virol. 2004, 78, 9007–9015. [Google Scholar] [CrossRef] [Green Version]
- Chou, M.L.; Burnouf, T.; Chang, S.P.; Hung, T.C.; Lin, C.C.; Richardson, C.D.; Lin, L.T. TnBPTriton X-45 treatment of plasma for transfusion efficiently inactivates hepatitis C virus. PLoS ONE 2015, 10, e0117800. [Google Scholar] [CrossRef] [Green Version]
- Sanclemente-Alaman, I.; Moreno-Jimenez, L.; Benito-Martin, M.S.; Canales-Aguirre, A.; Matias-Guiu, J.A.; Matias-Guiu, J.; Gomez-Pinedo, U. Experimental Models for the Study of Central Nervous System Infection by SARS-CoV-2. Front. Immunol. 2020, 11, 2163. [Google Scholar] [CrossRef]
- Sholukh, A.M.; Fiore-Gartland, A.; Ford, E.S.; Miner, M.D.; Hou, Y.J.; Tse, L.V.; Kaiser, H.; Zhu, H.; Lu, J.; Madarampalli, B.; et al. Evaluation of Cell-Based and Surrogate SARS-CoV-2 Neutralization Assays. J. Clin. Microbiol. 2021, 59, e0052721. [Google Scholar] [CrossRef]
- Zucker, I.; Lester, Y.; Alter, J.; Werbner, M.; Yecheskel, Y.; Gal-Tanamy, M.; Dessau, M. Pseudoviruses for the assessment of coronavirus disinfection by ozone. Env. Chem. Lett. 2021, 19, 1779–1785. [Google Scholar] [CrossRef]
- Hu, J.; Gao, Q.; He, C.; Huang, A.; Tang, N.; Wang, K. Development of cell-based pseudovirus entry assay to identify potential viral entry inhibitors and neutralizing antibodies against SARS-CoV-2. Genes Dis. 2020, 7, 551–557. [Google Scholar] [CrossRef]
- Darnell, M.E.R.; Taylor, D.R. Evaluation of inactivation methods for severe acute respiratory syndrome coronavirus in noncellular blood products. Transfusion 2006, 46, 1770–1777. [Google Scholar] [CrossRef] [Green Version]
- Barahona Afonso, A.F.; Joao, C.M. The Production Processes and Biological Effects of Intravenous Immunoglobulin. Biomolecules 2016, 6, 15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mould, G.P.; Sutton, J.A.; Matejtschuk, P.; Gascoigne, E.W.; Dash, C.H. Dash. Solvent detergent treatment does not alter the tolerance or uptake of human normal. Vox Sang 2001, 80, 151–158. [Google Scholar] [CrossRef] [PubMed]
- Koehler, M.; Ray, A.; Moreira, R.A.; Juniku, B.; Poma, A.B.; Alsteens, D. Molecular insights into receptor binding energetics and neutralization of SARS-CoV-2 variants. Nat. Commun. 2021, 12, 6977. [Google Scholar] [CrossRef] [PubMed]
- Buchacher, A.; Iberer, G. Purification of intravenous immunoglobulin G from human plasma-aspects of yield and virus safety. Biotechnol. J. 2006, 1, 148–163. [Google Scholar] [CrossRef] [PubMed]
- Radosevich, M.; Burnouf, T. Intravenous immunoglobulin G: Trends in production methods, quality control and quality assurance. Vox Sang 2010, 98, 12–28. [Google Scholar] [CrossRef]
- Rabenau, H.F.; Biesert, L.; Schmidt, T.; Bauer, G.; Cinatl, J.; Doerr, H.W. SARS-coronavirus (SARS-CoV) and the safety of a solvent/detergent (S/D) treated immunoglobulin preparation. Biologicals 2005, 33, 95–99. [Google Scholar] [CrossRef]
- Kang, B.; Zhang, X.; Wu, Z.; Qi, H.; Wang, Z. Solubilization capacity of nonionic surfactant micelles exhibiting strong influence on export of intracellular pigments in Monascus fermentation. Microb. Biotechnol. 2013, 6, 540–550. [Google Scholar] [CrossRef]
- Horowitz, B.; Bonomo, R.; Prince, A.M.; Chin, S.N.; Brotman, B.; Shulman, R.W. Solvent/detergent-treated plasma: A virus-inactivated substitute for fresh frozen plasma. Blood 1992, 79, 826–831. [Google Scholar] [CrossRef] [Green Version]
- Piët, M.P.; Chin, S.; Prince, A.M.; Brotman, B.; Cundell, A.M.; Horowitz, B. The use of tri(n-butyl)phosphate detergent mixtures to inactivate hepatitis viruses and human immunodeficiency virus in plasma and plasma’s subsequent fractionation. Transfusion 1990, 30, 591–598. [Google Scholar] [CrossRef]
- Toth, D. Neutralization Of SARS-CoV-2 Variants By A Human Polyclonal Antibody Therapeutic (COVID-HIG, NP-028) With High Neutralizing Titers To SARS-CoV-2. bioRxiv 2022. [Google Scholar] [CrossRef]
- Burnouf, T. An overview of plasma fractionation. Ann. Blood 2018, 3, 33. [Google Scholar] [CrossRef]
- Focosi, D.; Franchini, M. Impact of pathogen-reduction technologies on COVID-19 convalescent plasma potency. Transfus. Clin. Biol. 2021, 3188, 3. [Google Scholar] [CrossRef] [PubMed]
- Eickmann, M.; Gravemann, U.; Handke, W.; Tolksdorf, F.; Reichenberg, S.; Muller, T.H.; Seltsam, A. Inactivation of three emerging viruses-severe acute respiratory syndrome coronavirus, Crimean-Congo haemorrhagic fever virus and Nipah virus-in platelet concentrates by ultraviolet C light and in plasma by methylene blue plus visible light. Vox Sang 2020, 115, 146–151. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eickmann, M.; Gravemann, U.; Handke, W.; Tolksdorf, F.; Reichenberg, S.; Muller, T.H.; Seltsam, A. Inactivation of Ebola virus and Middle East respiratory syndrome coronavirus in platelet concentrates and plasma by ultraviolet C light and methylene blue plus visible light, respectively. Transfusion 2018, 58, 2202–2207. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Keil, S.D.; Bowen, R.; Marschner, S. Inactivation of Middle East respiratory syndrome coronavirus (MERS-CoV) in plasma products using a riboflavin-based and ultraviolet light-based photochemical treatment. Transfusion 2016, 56, 2948–2952. [Google Scholar] [CrossRef] [Green Version]
- Keil, S.D.; Ragan, I.; Yonemura, S.; Hartson, L.; Dart, N.K.; Bowen, R. Inactivation of severe acute respiratory syndrome coronavirus 2 in plasma and platelet products using a riboflavin and ultraviolet light-based photochemical treatment. Vox Sang 2020, 115, 495–501. [Google Scholar] [CrossRef]
- Azhar, E.I.; Hindawi, S.I.; El-Kafrawy, S.A.; Hassan, A.M.; Tolah, A.M.; Alandijany, T.A.; Bajrai, L.H.; Damanhouri, G.A. Amotosalen and ultraviolet A light treatment efficiently inactivates severe acute respiratory syndrome coronavirus 2 (SARS-CoV-2) in human plasma. Vox Sang 2021, 116, 673–681. [Google Scholar] [CrossRef]
- Hindawi, S.I.; Hashem, A.M.; Damanhouri, G.A.; El-Kafrawy, S.A.; Tolah, A.M.; Hassan, A.M.; Azhar, E.I. Inactivation of Middle East respiratory syndrome-coronavirus in human plasma using amotosalen and ultraviolet A light. Transfusion 2018, 58, 52–59. [Google Scholar] [CrossRef]
- Franchini, M.; Baldanti, F.; Percivalle, E.; Ferrari, A.; Biasi, P.; Liumbruno, G.M.; Glingani, C. Impact of pathogen reduction technology on neutralizing antibody titer in COVID-19 convalescent plasma. Transfus. Clin. Et Biol. 2022, 29, 101. [Google Scholar] [CrossRef]
- Raster, J.; Zimmermann, K.; Wesche, J.; Aurich, K.; Greinacher, A.; Selleng, K. Effect of Methylene Blue Pathogen Inactivation on the Integrity of Immunoglobulin M and G. Transfus. Med. Hemother 2021, 48, 148–152. [Google Scholar] [CrossRef]
- Geisen, C.; Kann, G.; Strecker, T.; Wolf, T.; Schuttfort, G.; van Kraaij, M.; MacLennan, S.; Rummler, S.; Weinigel, C.; Eickmann, M.; et al. Pathogen-reduced Ebola virus convalescent plasma: First steps towards standardization of manufacturing and quality control including assessment of Ebola-specific neutralizing antibodies. Vox Sang 2016, 110, 329–335. [Google Scholar] [CrossRef] [PubMed]
- Dean, C.L.; Hooper, J.W.; Dye, J.M.; Zak, S.E.; Koepsell, S.A.; Corash, L.; Benjamin, R.J.; Kwilas, S.; Bonds, S.; Winkler, A.M.; et al. Characterization of Ebola convalescent plasma donor immune response and psoralen treated plasma in the United States. Transfusion 2020, 60, 1024–1031. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.H.; Ngo, C.; Yeh, I.N.; Uemura, Y. Antibody Fc functional activity of intravenous immunoglobulin preparations treated with solvent-detergent for virus inactivation. Vox Sang 1994, 67, 337–344. [Google Scholar] [CrossRef] [PubMed]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Belem, W.F.; Liu, C.-H.; Hu, Y.-T.; Burnouf, T.; Lin, L.-T. Validation of Viral Inactivation Protocols for Therapeutic Blood Products against Severe Acute Respiratory Syndrome Coronavirus-2 (SARS-CoV-2). Viruses 2022, 14, 2419. https://doi.org/10.3390/v14112419
Belem WF, Liu C-H, Hu Y-T, Burnouf T, Lin L-T. Validation of Viral Inactivation Protocols for Therapeutic Blood Products against Severe Acute Respiratory Syndrome Coronavirus-2 (SARS-CoV-2). Viruses. 2022; 14(11):2419. https://doi.org/10.3390/v14112419
Chicago/Turabian StyleBelem, Wendimi Fatimata, Ching-Hsuan Liu, Yee-Tung Hu, Thierry Burnouf, and Liang-Tzung Lin. 2022. "Validation of Viral Inactivation Protocols for Therapeutic Blood Products against Severe Acute Respiratory Syndrome Coronavirus-2 (SARS-CoV-2)" Viruses 14, no. 11: 2419. https://doi.org/10.3390/v14112419