
Cross-Reactive T Cell Response Exists in Chronic Lymphocytic Choriomeningitis Virus Infection upon Pichinde Virus Challenge

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Ethics Statement
2.2. Mice, Viral Infection and Treatment
2.3. Synthetic Peptides and Dextramers
2.4. Cell Surface, Intracellular and Dextramer Staining for Flow Cytometry
2.5. Intracellular Cytokine Measurement
2.6. Viral Titer Determination
2.7. Next-Generation Sequencing
2.8. Sequencing Analysis
2.9. Statistics
3. Results
3.1. Diminished T Cell Responses during Sequential PICV Infection in a Setting of Chronic LCMV Infection
3.2. Cross-Reactive Epitope-Specific T Cell Responses Existed in Chronically LCMV Infected Mice upon PICV Infection
3.3. Checkpoint-Inhibitor Treatment Had No Effects on the Cross-Reactive NP205-Specific T Cells in LCMVcl13+PICV Infected Mice
3.4. Distinct Phenotype of Cross-Reactive and Non-Cross-Reactive T Cells in LCMVcl13+PICV
3.5. No Major Differences in the NP205-Specific TCR Repertoire between LCMV+PICV and LCMVcl13+PICV Infected Mice
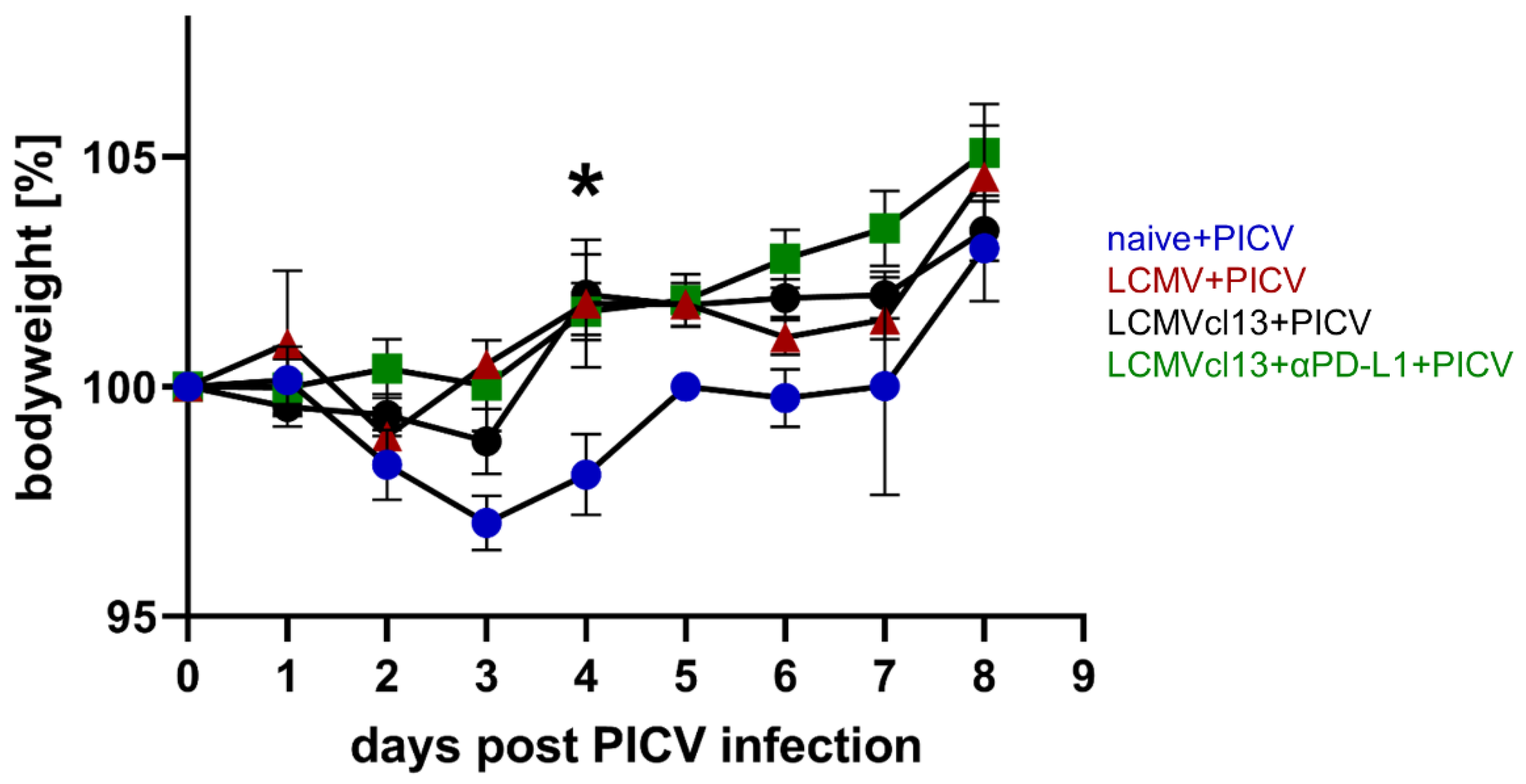
3.6. Chronically LCMVcl13 Infected Mice Are Protected against PICV Induced Weight Loss
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
References
- Cornberg, M.; Chen, A.T.; Wilkinson, L.A.; Brehm, M.A.; Kim, S.; Calcagno, C.; Ghersi, D.; Puzone, R.; Celada, F.; Welsh, R.M.; et al. Narrowed TCR Repertoire and Viral Escape as a Consequence of Heterologous Immunity. J. Clin. Investig. 2006, 116, 1443–1450. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Welsh, R.M.; Che, J.W.; Brehm, M.A.; Selin, L.K. Heterologous Immunity between Viruses. Immunol. Rev. 2010, 235, 244–266. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barton, E.S.; White, D.W.; Cathelyn, J.S.; Brett-McClellan, K.A.; Engle, M.; Diamond, M.S.; Miller, V.L.; Virgin, H.W. Herpesvirus Latency Confers Symbiotic Protection from Bacterial Infection. Nature 2007, 447, 326–329. [Google Scholar] [CrossRef] [PubMed]
- Sharma, S.; Thomas, P.G. The Two Faces of Heterologous Immunity: Protection Or Immunopathology. J. Leukoc. Biol. 2014, 95, 405–416. [Google Scholar] [CrossRef] [Green Version]
- Brehm, M.A.; Pinto, A.K.; Daniels, K.A.; Schneck, J.P.; Welsh, R.M.; Selin, L.K. T Cell Immunodominance and Maintenance of Memory Regulated by Unexpectedly Cross-Reactive Pathogens. Nat. Immunol. 2002, 3, 627–634. [Google Scholar] [CrossRef]
- Wlodarczyk, M.F.; Kraft, A.R.; Chen, H.D.; Kenney, L.L.; Selin, L.K. Anti–IFN-Γ and Peptide-Tolerization Therapies Inhibit Acute Lung Injury Induced by Cross-Reactive Influenza A–Specific Memory T Cells. J. Immunol. 2013, 190, 1201936. [Google Scholar] [CrossRef] [Green Version]
- Kraft, A.R.M.; Wlodarczyk, M.F.; Kenney, L.L.; Selin, L.K. PC61 (Anti-CD25) Treatment Inhibits Influenza A Virus-Expanded Regulatory T Cells and Severe Lung Pathology during a Subsequent Heterologous Lymphocytic Choriomeningitis Virus Infection. J. Virol. 2013, 87, 12636–12647. [Google Scholar] [CrossRef] [Green Version]
- Selin, L.K.; Wlodarczyk, M.F.; Kraft, A.R.; Nie, S.; Kenney, L.L.; Puzone, R.; Celada, F. Heterologous Immunity: Immunopathology, Autoimmunity and Protection during Viral Infections. Autoimmunity 2011, 44, 328–347. [Google Scholar] [CrossRef] [Green Version]
- Selin, L.K.; Varga, S.M.; Wong, I.C.; Welsh, R.M. Protective Heterologous Antiviral Immunity and Enhanced Immunopathogenesis Mediated by Memory T Cell Populations. J. Exp. Med. 1998, 188, 1705–1715. [Google Scholar] [CrossRef] [Green Version]
- Lipsitch, M.; Grad, Y.H.; Sette, A.; Crotty, S. Cross-Reactive Memory T Cells and Herd Immunity to SARS-CoV-2. Nat. Rev. Immunol. 2020, 20, 709–713. [Google Scholar] [CrossRef]
- Grifoni, A.; Weiskopf, D.; Ramirez, S.I.; Mateus, J.; Dan, J.M.; Moderbacher, C.R.; Rawlings, S.A.; Sutherland, A.; Premkumar, L.; Jadi, R.S.; et al. Targets of T Cell Responses to SARS-CoV-2 Coronavirus in Humans with COVID-19 Disease and Unexposed Individuals. Cell 2020, 181, 1489–1501.e15. [Google Scholar] [CrossRef]
- Kundu, R.; Narean, J.S.; Wang, L.; Fenn, J.; Pillay, T.; Fernandez, N.D.; Conibear, E.; Koycheva, A.; Davies, M.; Tolosa-Wright, M.; et al. Cross-Reactive Memory T Cells Associate with Protection Against SARS-CoV-2 Infection in COVID-19 Contacts. Nat. Commun. 2022, 13, 1–8. [Google Scholar] [CrossRef]
- Loyal, L.; Braun, J.; Henze, L.; Kruse, B.; Dingeldey, M.; Reimer, U.; Kern, F.; Schwarz, T.; Mangold, M.; Unger, C.; et al. Cross-Reactive CD4+ T Cells Enhance SARS-CoV-2 Immune Responses upon Infection and Vaccination. Science 2021, 374, eabh1823. [Google Scholar] [CrossRef]
- Selin, L.K.; Brehm, M.A.; Naumov, Y.N.; Cornberg, M.; Kim, S.; Clute, S.C.; Welsh, R.M. Memory of Mice and Men: CD8+ T-Cell Cross-Reactivity and Heterologous Immunity. Immunol. Rev. 2006, 211, 164–181. [Google Scholar] [CrossRef]
- Chen, A.T.; Cornberg, M.; Gras, S.; Guillonneau, C.; Rossjohn, J.; Trees, A.; Emonet, S.; de la Torre, J.C.; Welsh, R.M.; Selin, L.K. Loss of Anti-Viral Immunity by Infection with a Virus Encoding a Cross-Reactive Pathogenic Epitope. PLoS Pathog. 2012, 8, e1002633. [Google Scholar] [CrossRef] [Green Version]
- Welsh, R.M.; Selin, L.K. No One is Naive: The Significance of Heterologous T-Cell Immunity. Nat. Rev. Immunol. 2002, 2, 417–426. [Google Scholar] [CrossRef]
- Che, J.W.; Daniels, K.A.; Selin, L.K.; Welsh, R.M. Heterologous Immunity and Persistent Murine Cytomegalovirus Infection. J. Virol. 2017, 91, e01386-16. [Google Scholar] [CrossRef] [Green Version]
- Honke, N.; Shaabani, N.; Merches, K.; Gassa, A.; Kraft, A.; Ehrhardt, K.; Häussinger, D.; Löhning, M.; Dittmer, U.; Hengel, H.; et al. Immunoactivation Induced by Chronic Viral Infection Inhibits Viral Replication and Drives Immunosuppression through Sustained IFN-I Responses. Eur. J. Immunol. 2016, 46, 372–380. [Google Scholar] [CrossRef] [Green Version]
- Bergthaler, A.; Flatz, L.; Hegazy, A.N.; Johnson, S.; Horvath, E.; Löhning, M.; Pinschewer, D.D. Viral Replicative Capacity is the Primary Determinant of Lymphocytic Choriomeningitis Virus Persistence and Immunosuppression. Proc. Natl. Acad. Sci. USA 2010, 107, 21641–21646. [Google Scholar] [CrossRef] [Green Version]
- Souquette, A.; Thomas, P.G. Past Life and Future Effects—How Heterologous Infections Alter Immunity to Influenza Viruses. Front. Immunol. 2018, 9, 1071. [Google Scholar] [CrossRef]
- Stelekati, E.; Wherry, E.J. Chronic Bystander Infections and Immunity to Unrelated Antigens. Cell Host Microbe 2012, 12, 458–469. [Google Scholar] [CrossRef] [Green Version]
- Kahan, S.M.; Wherry, E.J.; Zajac, A.J. T Cell Exhaustion during Persistent Viral Infections. Virology 2015, 479–480, 180–193. [Google Scholar] [CrossRef]
- Wherry, J.E.; Blattman, J.N.; Murali-Krishna, K.; van der Most, R.; Ahmed, R. Viral Persistence Alters CD8 T-Cell Immunodominance and Tissue Distribution and Results in Distinct Stages of Functional Impairment. J. Virol. 2003, 77, 4911–4927. [Google Scholar] [CrossRef] [Green Version]
- Wherry, J.E. T Cell Exhaustion. Nat. Immunol. 2011, 12, 492–499. [Google Scholar] [CrossRef]
- Wherry, J.E.; Kurachi, M. Molecular and Cellular Insights into T Cell Exhaustion. Nat. Rev. Immunol. 2015, 15, 486–499. [Google Scholar] [CrossRef]
- Barber, D.L.; Wherry, E.J.; Masopust, D.; Zhu, B.; Allison, J.P.; Sharpe, A.H.; Freeman, G.J.; Ahmed, R. Restoring Function in Exhausted CD8 T Cells during Chronic Viral Infection. Nature 2006, 439, 682–687. [Google Scholar] [CrossRef]
- Akinleye, A.; Rasool, Z. Immune Checkpoint Inhibitors of PD-L1 as Cancer Therapeutics. J. Hematol. Oncol. 2019, 12, 92. [Google Scholar] [CrossRef] [Green Version]
- Finn, R.S.; Qin, S.; Ikeda, M.; Galle, P.R.; Ducreux, M.; Kim, T.; Kudo, M.; Breder, V.; Merle, P.; Kaseb, A.O.; et al. Atezolizumab Plus Bevacizumab in Unresectable Hepatocellular Carcinoma. N. Engl. J. Med. 2020, 382, 1894–1905. [Google Scholar] [CrossRef]
- Gane, E.; Verdon, D.J.; Brooks, A.E.; Gaggar, A.; Nguyen, A.H.; Subramanian, G.M.; Schwabe, C.; Dunbar, P.R. Anti-PD-1 Blockade with Nivolumab with and without Therapeutic Vaccination for Virally Suppressed Chronic Hepatitis B: A Pilot Study. J. Hepatol. 2019, 71, 900–907. [Google Scholar] [CrossRef]
- A Phase 2a, Open-Label Study to Evaluate the Safety and Efficacy of Selgantolimod (SLGN)-Containing Combination Therapies for the Treatment of Chronic Hepatitis B (CHB). Available online: https://clinicaltrials.gov/ct2/show/NCT04891770 (accessed on 9 August 2022).
- Matloubian, M.; Kolhekar, S.R.; Somasundaram, T.; Ahmed, R. Molecular Determinants of Macrophage Tropism and Viral Persistence: Importance of Single Amino Acid Changes in the Polymerase and Glycoprotein of Lymphocytic Choriomeningitis Virus. J. Virol. 1993, 67, 7340–7349. [Google Scholar] [CrossRef]
- Salvato, M.; Borrow, P.; Shimomaye, E.; Oldstone, M.B. Molecular Basis of Viral Persistence: A Single Amino Acid Change in the Glycoprotein of Lymphocytic Choriomeningitis Virus is Associated with Suppression of the Antiviral Cytotoxic T-Lymphocyte Response and Establishment of Persistence. J. Virol. 1991, 65, 1863–1869. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Welsh, R.M.; Seedhom, M.O. Lymphocytic Choriomeningitis Virus (LCMV): Propagation, Quantitation, and Storage. Curr. Protoc. Microbiol. 2008, 15, 15A.1.1–15A.1.11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Klein, S.; Ghersi, D.; Manns, M.P.; Prinz, I.; Cornberg, M.; Kraft, A.R.M. PD-L1 Checkpoint Inhibition Narrows the Antigen-Specific T Cell Receptor Repertoire in Chronic Lymphocytic Choriomeningitis Virus Infection. J. Virol. 2020, 94, e00795-20. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.; Zhou, Y.; Chen, Y.; Gu, J. Fastp: An Ultra-Fast all-in-One FASTQ Preprocessor. Bioinformatics 2018, 34, i884–i890. [Google Scholar] [CrossRef] [PubMed]
- Bolotin, D.A.; Poslavsky, S.; Mitrophanov, I.; Shugay, M.; Mamedov, I.Z.; Putintseva, E.V.; Chudakov, D.M. MiXCR: Software for Comprehensive Adaptive Immunity Profiling. Nat. Methods 2015, 12, 380–381. [Google Scholar] [CrossRef] [PubMed]
- Warren, R.L.; Freeman, J.D.; Zeng, T.; Choe, G.; Munro, S.; Moore, R.; Webb, J.R.; Holt, R.A. Exhaustive T-Cell Repertoire Sequencing of Human Peripheral Blood Samples Reveals Signatures of Antigen Selection and a Directly Measured Repertoire Size of at Least 1 Million Clonotypes. Genome Res. 2011, 21, 790–797. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shugay, M.; Bagaev, D.V.; Turchaninova, M.A.; Bolotin, D.A.; Britanova, O.V.; Putintseva, E.V.; Pogorelyy, M.V.; Nazarov, V.I.; Zvyagin, I.V.; Kirgizova, V.I.; et al. VDJtools: Unifying Post-Analysis of T Cell Receptor Repertoires. PLoS Comput. Biol. 2015, 11, e1004503. [Google Scholar] [CrossRef] [Green Version]
- Dash, P.; Fiore-Gartland, A.J.; Hertz, T.; Wang, G.C.; Sharma, S.; Souquette, A.; Crawford, J.C.; Clemens, E.B.; Nguyen, T.H.O.; Kedzierska, K.; et al. Quantifiable Predictive Features Define Epitope-Specific T Cell Receptor Repertoires. Nature 2017, 547, 89–93. [Google Scholar] [CrossRef] [Green Version]
- Gil, A.; Kamga, L.; Chirravuri-Venkata, R.; Aslan, N.; Clark, F.; Ghersi, D.; Luzuriaga, K.; Selin, L.K. Epstein-Barr Virus Epitope-Major Histocompatibility Complex Interaction Combined with Convergent Recombination Drives Selection of Diverse T Cell Receptor A and Β Repertoires. MBio 2020, 11, e00250-20. [Google Scholar] [CrossRef] [Green Version]
- Agrawal, B. Heterologous Immunity: Role in Natural and Vaccine-Induced Resistance to Infections. Front. Immunol. 2019, 10, 2631. [Google Scholar] [CrossRef]
- Kim, S.; Brehm, M.A.; Welsh, R.M.; Selin, L.K. Dynamics of Memory T Cell Proliferation Under Conditions of Heterologous Immunity and Bystander Stimulation. J. Immunol. 2002, 169, 90–98. [Google Scholar] [CrossRef] [Green Version]
- Wedemeyer, H.; Mizukoshi, E.; Davis, A.R.; Bennink, J.R.; Rehermann, B. Cross-Reactivity between Hepatitis C Virus and Influenza A Virus Determinant-Specific Cytotoxic T Cells. J. Virol. 2001, 75, 11392–11400. [Google Scholar] [CrossRef] [Green Version]
- Saito, F.; Ito, T.; Connett, J.M.; Schaller, M.A.; Carson, W.F.; Hogaboam, C.M.; Rochford, R.; Kunkel, S.L. MHV68 Latency Modulates the Host Immune Response to Influenza A Virus. Inflammation 2013, 36, 1295–1303. [Google Scholar] [CrossRef] [Green Version]
- Furman, D.; Jojic, V.; Sharma, S.; Shen-Orr, S.S.; Angel, C.J.L.; Onengut-Gumuscu, S.; Kidd, B.A.; Maecker, H.T.; Concannon, P.; Dekker, C.L.; et al. Cytomegalovirus Infection Enhances the Immune Response to Influenza. Sci. Transl. Med. 2015, 7, 281. [Google Scholar] [CrossRef] [Green Version]
- Aliabadi, E.; Urbanek-Quaing, M.; Maasoumy, B.; Bremer, B.; Grasshoff, M.; Li, Y.; Niehaus, C.E.; Wedemeyer, H.; Kraft, A.R.M.; Cornberg, M. Impact of HBsAg and HBcrAg Levels on Phenotype and Function of HBV-Specific T Cells in Patients with Chronic Hepatitis B Virus Infection. Gut 2021, 71, 2300–2312. [Google Scholar] [CrossRef]
- Aregay, A.; Owusu Sekyere, S.; Deterding, K.; Port, K.; Dietz, J.; Berkowski, C.; Sarrazin, C.; Manns, M.P.; Cornberg, M.; Wedemeyer, H. Elimination of Hepatitis C Virus has Limited Impact on the Functional and Mitochondrial Impairment of HCV-Specific CD8+ T Cell Responses. J. Hepatol. 2019, 71, 889–899. [Google Scholar] [CrossRef]
- Owusu Sekyere, S.; Suneetha, P.V.; Hardtke, S.; Falk, C.S.; Hengst, J.; Manns, M.P.; Cornberg, M.; Wedemeyer, H.; Schlaphoff, V. Type I Interferon Elevates Co-Regulatory Receptor Expression on CMV- and EBV-Specific CD8 T Cells in Chronic Hepatitis C. Front. Immunol. 2015, 6, 270. [Google Scholar] [CrossRef] [Green Version]
- McLane, L.M.; Abdel-Hakeem, M.S.; Wherry, E.J. CD8 T Cell Exhaustion during Chronic Viral Infection and Cancer. Annu. Rev. Immunol. 2019, 37, 457–495. [Google Scholar] [CrossRef] [Green Version]
- Himmel, M.E.; Saibil, S.D.; Saltman, A.P. Immune Checkpoint Inhibitors in Cancer Immunotherapy. CMAJ 2020, 192, E651. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Splenocyte Number | α-CD3 Response [% IFNγ+ CD8+ T Cells] | |||||
|---|---|---|---|---|---|---|
| Naive | LCMV | LCMVcl13 | Naive | LCMV | LCMVcl13 | |
| (I) no sequential infection (mean ± SEM) (n = 2–13) | 6.5 × 107 ± 1.1 × 107 | 3.7 × 107 ± 6.2 × 106 | 2.7 × 107 ± 2.7 × 106 | 4.4 ± 0.2 | 47.9 ± 3.6 | 28.0 ± 3.5 |
| (II) plus sequential infection (mean ± SEM) (n = 6–14) | 8.1 × 107 ± 8.3 × 106 | 7.2 × 107 ± 7.1 × 106 | 4.3 × 107 ± 6.8 × 106 | 45.9 ± 4.2 | 51.3 ± 3.3 | 35.1 ± 2.5 |
| p-value (I) vs. (II) | n.s. | 0.0029 | 0.026 | 0.0019 | n.s. | n.s. |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mischke, J.; Klein, S.; Seamann, A.; Prinz, I.; Selin, L.; Ghersi, D.; Cornberg, M.; Kraft, A.R.M. Cross-Reactive T Cell Response Exists in Chronic Lymphocytic Choriomeningitis Virus Infection upon Pichinde Virus Challenge. Viruses 2022, 14, 2293. https://doi.org/10.3390/v14102293
Mischke J, Klein S, Seamann A, Prinz I, Selin L, Ghersi D, Cornberg M, Kraft ARM. Cross-Reactive T Cell Response Exists in Chronic Lymphocytic Choriomeningitis Virus Infection upon Pichinde Virus Challenge. Viruses. 2022; 14(10):2293. https://doi.org/10.3390/v14102293
Chicago/Turabian StyleMischke, Jasmin, Sebastian Klein, Austin Seamann, Immo Prinz, Liisa Selin, Dario Ghersi, Markus Cornberg, and Anke R.M. Kraft. 2022. "Cross-Reactive T Cell Response Exists in Chronic Lymphocytic Choriomeningitis Virus Infection upon Pichinde Virus Challenge" Viruses 14, no. 10: 2293. https://doi.org/10.3390/v14102293