
Prospective Alternate Hosts of an Emerging Polerovirus in Cotton Landscapes in the Southeastern United States

 ,
,  , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Maintenance of Plants, Virus Isolate, and the Insect Colony
2.2. Transmission of CLRDV to Test Plants
2.3. Detection and Quantitation of CLRDV
2.4. Virus Acquisition from Alternate Hosts by Aphids and CLRDV Quantitation
2.5. Back-Transmission Assays to Cotton
2.6. Alternate Plant Species as Hosts of Aphids
2.7. Statistical Analyses
3. Results
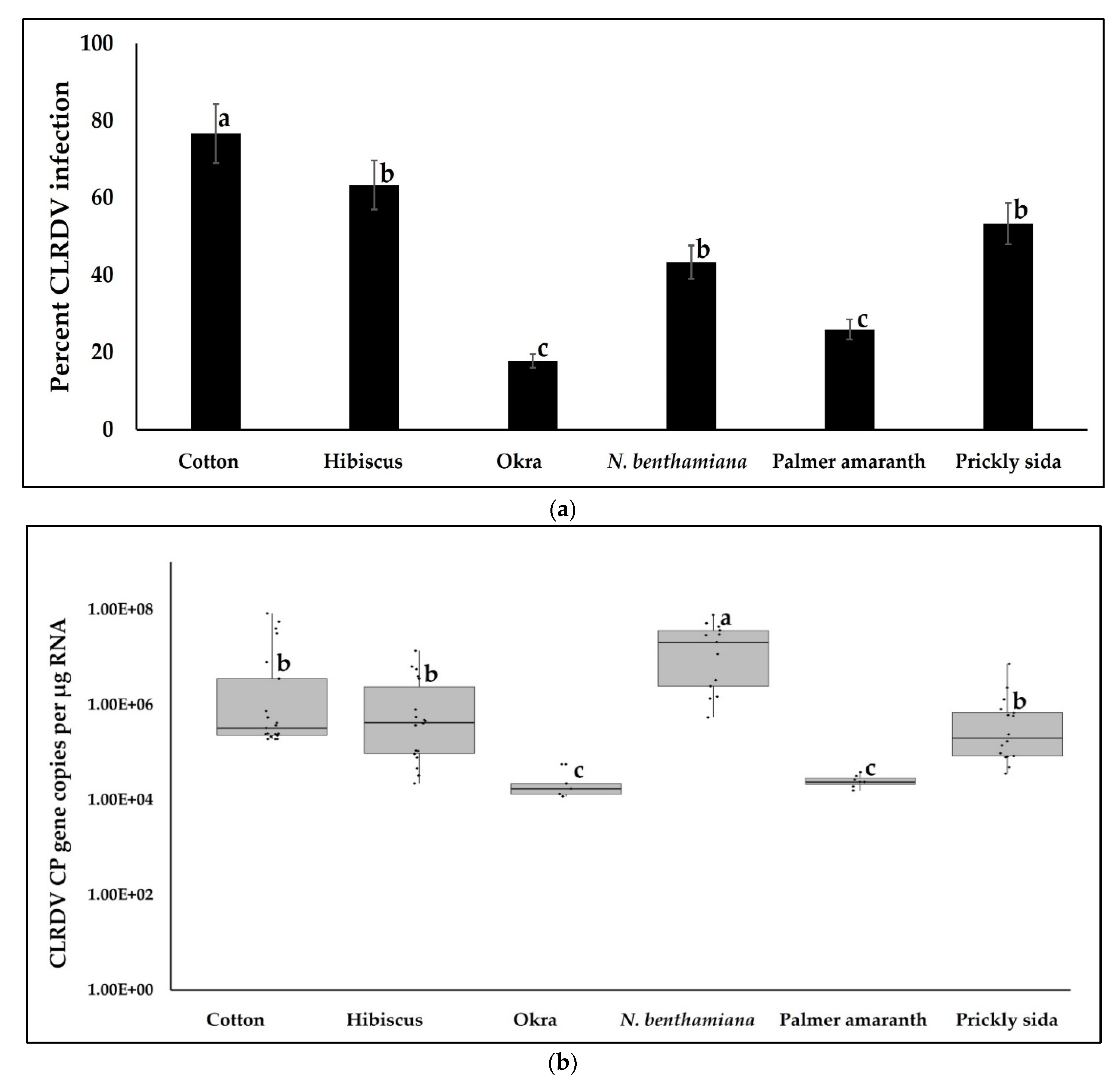
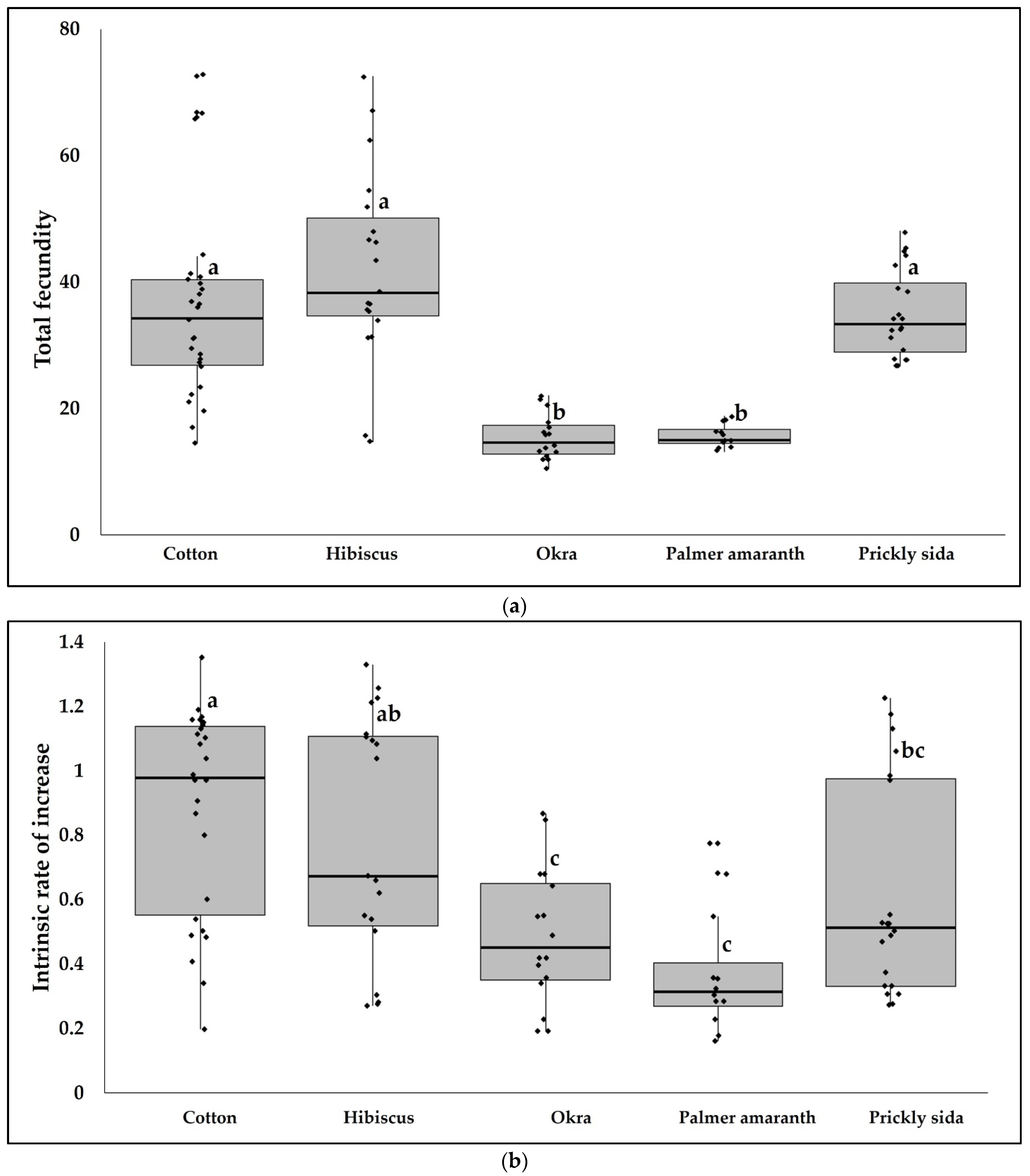
3.1. CLRDV Host Range and Virus Quantitation
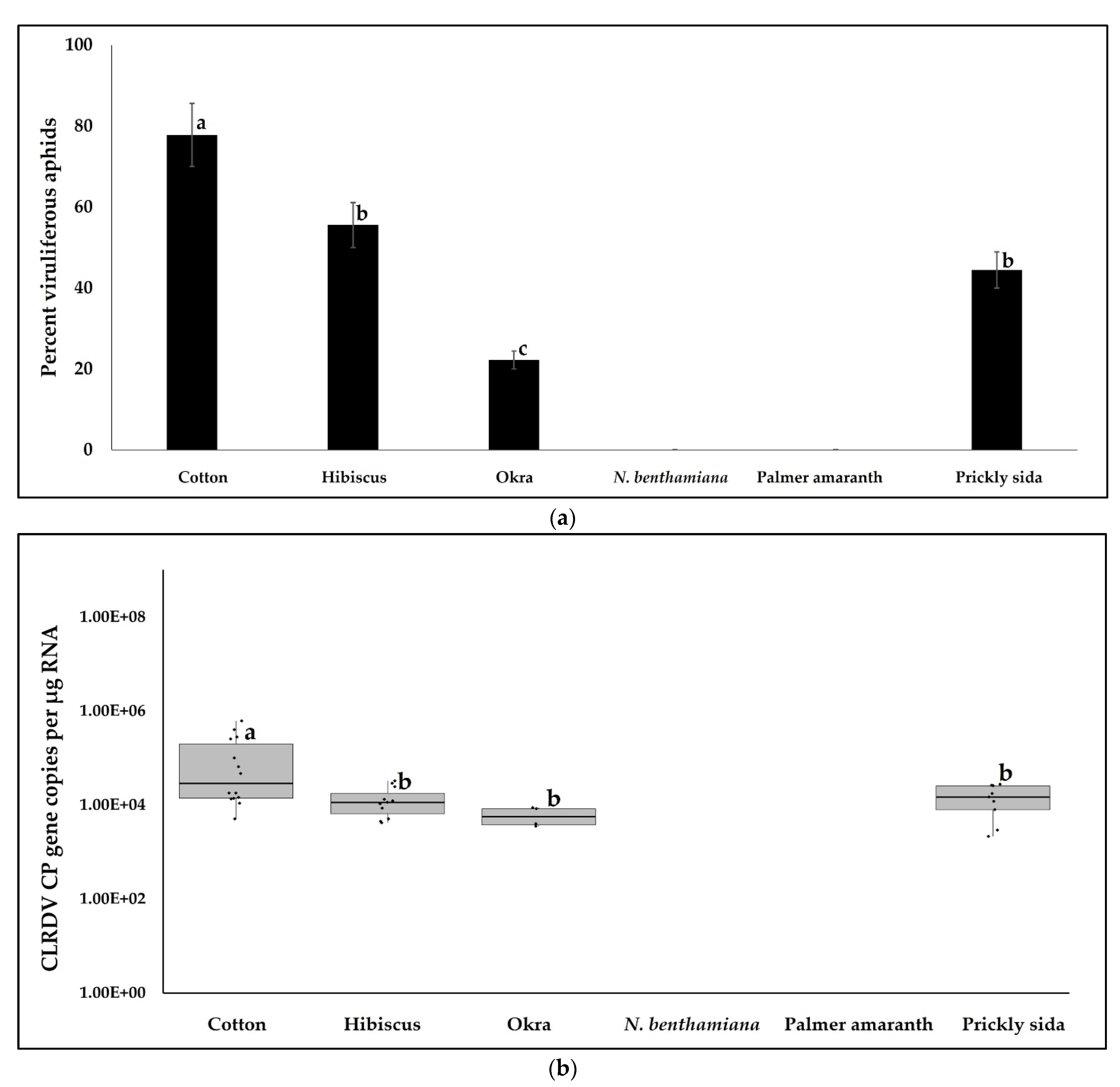
3.2. CLRDV Acquisition by Aphids from Cotton and Alternate Hosts and Virus Quantitation
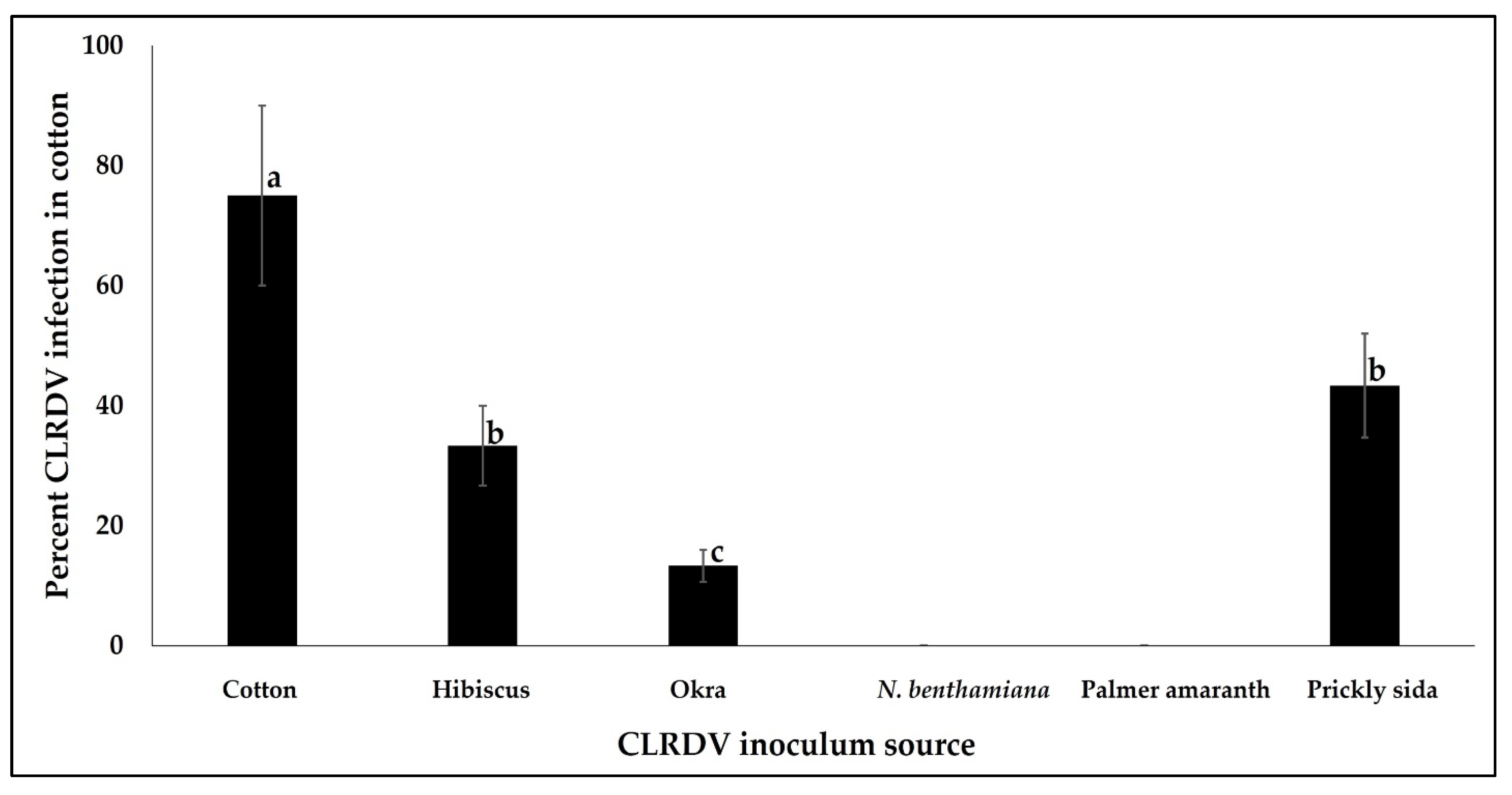
3.3. Back-Transmission of CLRDV to Cotton
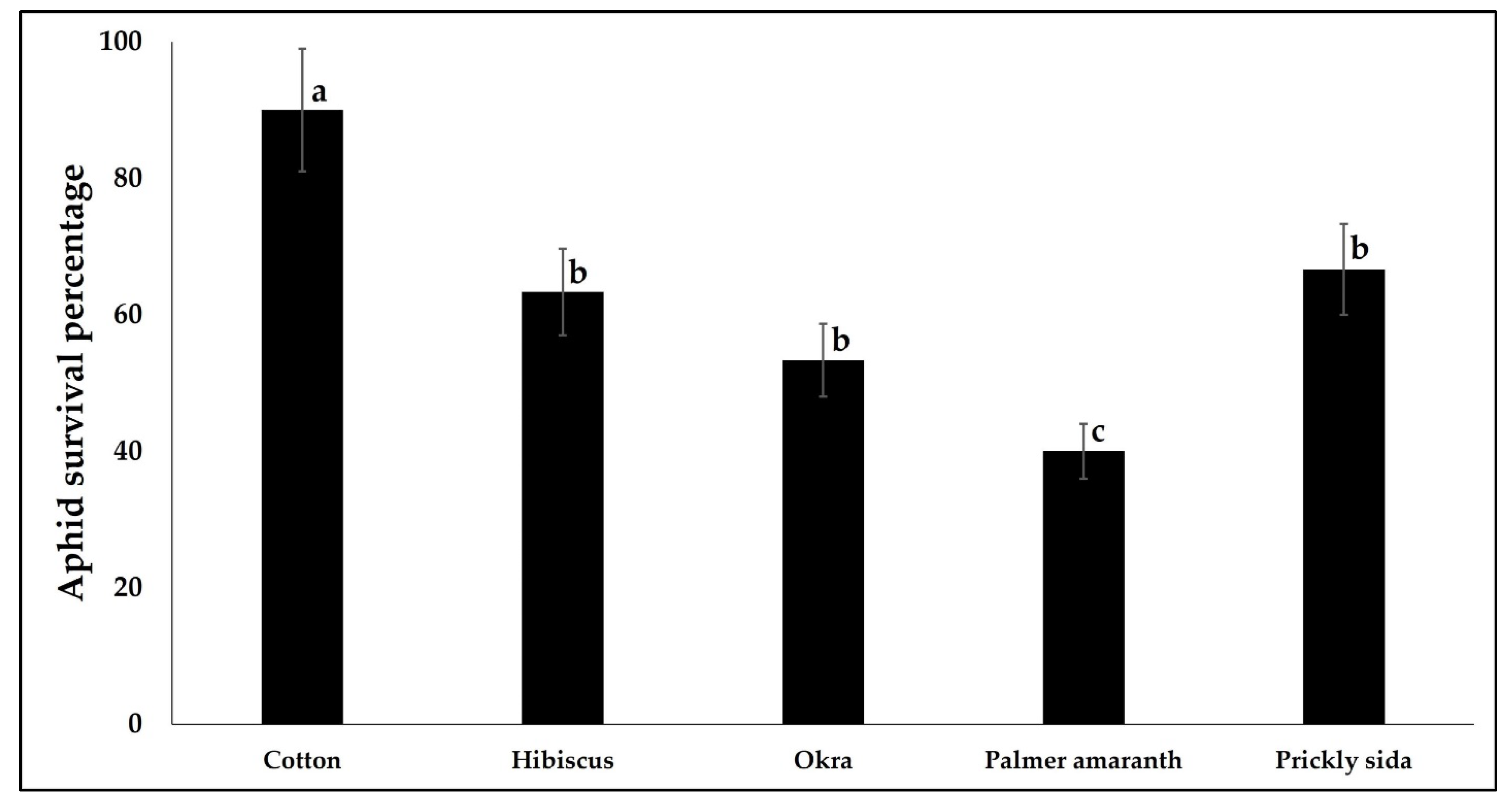
3.4. Suitability of Alternate Plant Species as Aphid Hosts
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Informed Consent Statement
Conflicts of Interest
References
- Wisler, G.C.; Norris, R.F. Interactions between Weeds and Cultivated Plants as Related to Management of Plant Pathogens. Weed Sci. 2005, 53, 914–917. [Google Scholar] [CrossRef]
- Hily, J.M.; García, A.; Moreno, A.; Plaza, M.; Wilkinson, M.D.; Fereres, A.; Fraile, A.; García-Arenal, F. The Relationship between Host Lifespan and Pathogen Reservoir Potential: An Analysis in the System Arabidopsis Thaliana-Cucumber Mosaic Virus. PLoS Pathog. 2014, 10, e1004492. [Google Scholar] [CrossRef] [PubMed]
- Brown, S.; Conner, K.; Hagan, A.; Jacobson, A.; Koebernick, J.; Lawrence, K.; Nichols, B. Report of A Research Review and Planning Meeting on Cotton Leafroll Dwarf Virus; Alabama, 2019, Cotton Incorporated: Cary, NC, USA, 2019. [Google Scholar]
- Mahas, J.W.; Hamilton, F.B.; Roberts, P.M.; Ray, C.H.; Miller, G.L.; Sharman, M.; Conner, K.; Bag, S.; Blythe, E.K.; Toews, M.D.; et al. Investigating the effects of planting date and aphis gossypii management on reducing the final incidence of cotton leafroll dwarf virus. Crop Prot. 2022, 158, 106005. [Google Scholar] [CrossRef]
- Sharman, M.; Lapbanjob, S.; Sebunruang, P.; Belot, J.L.; Galbieri, R.; Giband, M.; Suassuna, N. First Report of Cotton Leafroll Dwarf Virus in Thailand Using a Species-Specific PCR Validated with Isolates from Brazil. Australas. Plant Dis. Notes 2015, 10, 24. [Google Scholar] [CrossRef] [Green Version]
- Corrêa, R.L.; Silva, T.F.; Simões-Araújo, J.L.; Barroso, P.A.V.; Vidal, M.S.; Vaslin, M.F.S. Molecular Characterization of a Virus from the Family Luteoviridae Associated with Cotton Blue Disease. Arch. Virol. 2005, 150, 1357–1367. [Google Scholar] [CrossRef]
- Cauquil, J. Etudes Sur Une Maladie d’ Origine Virale Du Cotonnier: La Maladie Bleue. Cot. Fibres Trop. 1977, 32, 259–278. [Google Scholar]
- Avelar, S.; Schrimsher, D.W.; Lawrence, K.; Brown, J.K. First Report of Cotton Leafroll Dwarf Virus Associated with Cotton Blue Disease Symptoms in Alabama. Plant Dis. Notes 2019, 103, 592. [Google Scholar] [CrossRef]
- Iriarte, F.B.; Dey, K.K.; Small, I.M.; Conner, K.N.; O’Brien, G.K.; Johnson, L.; Savery, C.; Carter, E.; Sprague, D.; Nichols, R.L.; et al. First Report of Cotton Leafroll Dwarf Virus in Florida. Plant Dis. Notes 2020, 104, 2744. [Google Scholar] [CrossRef] [Green Version]
- Tabassum, A.; Bag, S.; Roberts, P.; Suassuna, N.; Chee, P.; Whitaker, J.R.; Conner, K.N.; Brown, J.; Nichols, R.L.; Kemerait, R.C. First Report of Cotton Leafroll Dwarf Virus Infecting Cotton in Georgia, U.S.A. Plant Dis. Notes 2019, 103, 1803. [Google Scholar] [CrossRef]
- Aboughanem-Sabanadzovic, N.; Allen, T.W.; Wilkerson, T.H.; Conner, K.N.; Sikora, E.J.; Nichols, R.L.; Sabanadzovic, S. First Report of Cotton Leafroll Dwarf Virus in Upland Cotton (Gossypium Hirsutum) in Mississippi. Plant Dis. 2019, 103, 1798. [Google Scholar] [CrossRef]
- Alabi, O.J.; Isakeit, T.; Vaughn, R.; Stelly, D.; Conner, K.N.; Gaytan, B.C.; Villegas, C.; Hitzelberger, C.; De Santiago, L.; Monclova-Santana, C.; et al. First Report of Cotton Leafroll Dwarf Virus Infecting Upland Cotton (Gossypium Hirsutum L.) in Texas. Plant Dis. Notes 2020, 104, 10–11. [Google Scholar] [CrossRef]
- Thiessen, L.D.; Schappe, T.; Zaccaron, M.; Conner, K.; Koebernick, J.; Jacobson, A.; Huseth, A. First Report of Cotton Leafroll Dwarf Virus in Cotton Plants Affected by Cotton Leafroll Dwarf Disease in North Carolina. Plant Dis. Notes 2020, 104, 3275. [Google Scholar] [CrossRef]
- Price, T.; Valverde, R.; Singh, R.; Davis, J.; Brown, S.; Jones, H. First Report of Cotton Leafroll Dwarf Virus in Louisiana. Plant Heal. Prog. 2020, 21, 142–143. [Google Scholar] [CrossRef]
- Wang, H.; Greene, J.; Mueller, J.; Conner, K.; Jacobson, A. First Report of Cotton Leafroll Dwarf Virus in Cotton Fields of South Carolina. Plant Dis. Notes 2020, 104, 2532. [Google Scholar] [CrossRef] [Green Version]
- Faske, T.R.; Stainton, D.; Aboughanem-Sabanadzovic, N.; Allen, T.W. First Report of Cotton Leafroll Dwarf Virus from Upland Cotton (Gossypium Hirsutum) in Arkansas. Plant Dis. Notes 2020, 104, 2742. [Google Scholar] [CrossRef]
- Ali, A.; Mokhtari, S.; Ferguson, C. First Report of Cotton Leafroll Dwarf Virus from Cotton (Gossypium Hirsutum) in Oklahoma. Plant Dis. Notes 2020, 104, 2531. [Google Scholar] [CrossRef] [Green Version]
- Ali, A.; Mokhtari, S. First Report of Cotton Leafroll Dwarf Virus Infecting Cotton (Gossypium Hirsutum) in Kansas. Plant Dis. 2020, 104, 1880. [Google Scholar] [CrossRef]
- Cascardo, R.S.; Arantes, I.L.G.; Silva, T.F.; Sachetto-Martins, G.; Vaslin, M.F.S.; Corrêa, R.L. Function and Diversity of P0 Proteins among Cotton Leafroll Dwarf Virus Isolates. Virol. J. 2015, 12, 123. [Google Scholar] [CrossRef]
- Silva, A.K.F.; Romanel, E.; Silva, T.F.; Castilhos, Y.; Schrago, C.G.; Galbieri, R.; Bélot, J.L.; Vaslin, M.F.S. Complete Genome Sequences of Two New Virus Isolates Associated with Cotton Blue Disease Resistance Breaking in Brazil. Arch. Virol. 2015, 160, 1371–1374. [Google Scholar] [CrossRef]
- Parkash, V.; Sharma, D.B.; Snider, J.; Bag, S.; Roberts, P.; Tabassum, A.; West, D.; Khanal, S.; Suassuna, N.; Chee, P. Effect of Cotton Leafroll Dwarf Virus on Physiological Processes and Yield of Individual Cotton Plants. Front. Plant Sci. 2021, 12, 734386. [Google Scholar] [CrossRef]
- Sõmera, M.; Fargette, D.; Hébrard, E.; Sarmiento, C. ICTV Virus Taxonomy Profile: Solemoviridae 2021. J. Gen. Virol. 2021, 102, 001707. [Google Scholar] [CrossRef] [PubMed]
- Delfosse, V.C.; Agrofoglio, Y.C.; Casse, M.F.; Kresic, I.B.; Hopp, H.E.; Ziegler-Graff, V.; Distéfano, A.J. The P0 Protein Encoded by Cotton Leafroll Dwarf Virus (CLRDV) Inhibits Local but Not Systemic RNA Silencing. Virus Res. 2014, 180, 70–75. [Google Scholar] [CrossRef] [PubMed]
- Distéfano, A.J.; Kresic, I.B.; Hopp, H.E. The Complete Genome Sequence of a Virus Associated with Cotton Blue Disease, Cotton Leafroll Dwarf Virus, Confirms That It Is a New Member of the Genus Polerovirus. Arch. Virol. 2010, 155, 1849–1854. [Google Scholar] [CrossRef] [PubMed]
- Tabassum, A.; Bag, S.; Suassuna, N.D.; Conner, K.N.; Chee, P.; Kemerait, R.C.; Roberts, P. Genome Analysis of Cotton Leafroll Dwarf Virus Reveals Variability in the Silencing Suppressor Protein, Genotypes and Genomic Recombinants in the USA. PLoS ONE 2021, 16, e0252523. [Google Scholar] [CrossRef]
- Ali, A.; Mokhtari, S. Complete Genome Sequence of Cotton Leafroll Dwarf Virus Isolated from Cotton in Texas, USA. Microbiol. Resour. Announc. 2020, 9, e01587-19. [Google Scholar] [CrossRef] [Green Version]
- Fereres, A.; Moreno, A. Behavioural Aspects Influencing Plant Virus Transmission by Homopteran Insects. Virus Res. 2009, 141, 158–168. [Google Scholar] [CrossRef]
- Heilsnis, B.; McLaughlin, A.; Conner, K.; Koebernick, J.; Jacobson, A.L. Vector Competency of Aphis Gossypii and Bemisia Tabaci to Transmit Cotton Leafroll Dwarf Virus. J. Cotton Sci. 2022, 26, 23–30. [Google Scholar] [CrossRef]
- Michelotto, M.D.; Busoli, A.C. Eficiência de Ninfas e Adultos de Aphis Gossypii Glov. Na Transmissão Do Vírus Do Mosaico Das Nervuras Do Algodoeiro. Bragantia 2003, 62, 255–259. [Google Scholar] [CrossRef] [Green Version]
- Gray, S.; Gildow, F.E. Luteovirus-Aphid Interactions. Annu. Rev. Phytopathol. 2003, 41, 539–566. [Google Scholar] [CrossRef]
- Catto, M.A.; Mugerwa, H.; Myers, B.K.; Pandey, S.; Dutta, B.; Srinivasan, R. A Review on Transcriptional Responses of Interactions between Insect Vectors and Plant Viruses. Cells 2022, 11, 693. [Google Scholar] [CrossRef]
- Michelotto, M.D.; Antonio, E.; Busoli, C. Efeito Da Época de Inoculação Do Vírus Do Mosaico Das Nervuras Por Aphis Gossypii Glover (Hemiptera: Aphididae) No Desenvolvimento e Na Produção Do Algodoeiro. Neotrop. Entomol. 2006, 35, 251–256. [Google Scholar]
- Michelotto, M.D.; Busoli, A.C. Biology of Cotton Aphid on Plants Infected by Cotton Vein Mosaic Virus. Bragantia 2009, 68, 1017–1024. [Google Scholar] [CrossRef]
- Feng, Y.; Krueger, E.N.; Liu, S.; Dorman, K.; Bonning, B.C.; Miller, W.A. Discovery of Known and Novel Viral Genomes in Soybean Aphid by Deep Sequencing. Phytobiomes J. 2017, 1, 36–45. [Google Scholar] [CrossRef] [Green Version]
- Sedhain, N.P.; Bag, S.; Morgan, K.; Carter, R.; Triana, P.; Whitaker, J.; Kemerait, R.C.; Roberts, P.M. Natural Host Range, Incidence on Overwintering Cotton and Diversity of Cotton Leafroll Dwarf Virus in Georgia USA. Crop Prot. 2021, 144, 105604. [Google Scholar] [CrossRef]
- Legarrea, S.; Barman, A.; Marchant, W.; Diffie, S.; Srinivasan, R. Temporal Effects of a Begomovirus Infection and Host Plant Resistance on the Preference and Development of an Insect Vector, Bemisia Tabaci, and Implications for Epidemics. PLoS ONE 2015, 10, e0142114. [Google Scholar] [CrossRef] [Green Version]
- Barman, A.K.; Gadhave, K.R.; Dutta, B.; Srinivasan, R. Plasticity in Host Utilization by Two Host-Associated Populations of Aphis Gossypii Glover. Bull. Entomol. Res. 2018, 108, 360–369. [Google Scholar] [CrossRef]
- Srinivasan, R.; Alvarez, J.M.; Bosque-Pérez, N.A.; Eigenbrode, S.D.; Novy, R.G. Effect of an Alternate Weed Host, Hairy Nightshade, Sofonum Sarrachoides, on the Biology of the Two Most Important Potato Leafroll Virus (Luteoviridae: Polerovirus) Vectors, Myzus Persicae and Macrosiphum Euphorbiae (Aphididae: Homoptera). Environ. Entomol. 2008, 37, 592–600. [Google Scholar] [CrossRef]
- Wyatt, I.J.; White, P.F. Simple Estimation of Intrinsic Increase Rates for Aphids and Tetranychid Mites. J. Appl. Ecol. 1977, 14, 757–766. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2021. [Google Scholar]
- Kumari, S.G.; Sharman, M.; Moukahel, A.; Ziyaev, Z.; Ahmed, S. First Report of Cotton Leafroll Dwarf Virus Affecting Chickpea (Cicer Arietinum) in Uzbekistan. Plant Dis. 2020, 104, 2532. [Google Scholar] [CrossRef]
- Mukherjee, A.K.; Mukherjee, P.K.; Kranthi, S. Genetic Similarity between Cotton Leafroll Dwarf Virus and Chickpea Stunt Disease Associated Virus in India. Plant Pathol. J. 2016, 32, 580–583. [Google Scholar] [CrossRef]
- Ingwell, L.L.; Bosque-Pérez, N.A. The Invasive Weed Ventenata Dubia Is a Host of Barley Yellow Dwarf Virus with Implications for an Endangered Grassland Habitat. Weed Res. 2014, 55, 62–70. [Google Scholar] [CrossRef]
- Alvarez, J.M.; Srinivasan, R. Evaluation of Hairy Nightshade as an Inoculum Source for Aphid-Mediated Transmission of Potato Leafroll Virus. J. Econ. Entomol. 2005, 98, 1101–1108. [Google Scholar] [CrossRef]
- Griesbach, J.A.; Steffenson, B.J.; Brown, M.P.; Falk, B.W.; Webster, R.K. Infection of Grasses by Barley Yellow Dwarf Viruses in California. Crop Sci. 1990, 30, 1173–1177. [Google Scholar] [CrossRef]
- Irwin, M.E.; Thresh, J.M. Epidemiology of Barley Yellow Dwarf: A Study in Ecological Complexity. Annu. Rev. Phytopathol. 1990, 28, 393–424. [Google Scholar] [CrossRef]
- Fingu-Mabola, J.C.; Martin, C.; Bawin, T.; Verheggen, F.J.; Francis, F. Does the Infectious Status of Aphids Influence Their Preference towards Healthy, Virus-Infected and Endophytically Colonized Plants? Insects 2020, 11, 435. [Google Scholar] [CrossRef] [PubMed]
- Eigenbrode, S.D.; Ding, H.; Shiel, P.; Berger, P.H. Volatiles from Potato Plants Infected with Potato Leafroll Virus Attract and Arrest the Virus Vector, Myzus Persicae (Homoptera: Aphididae). Proc. R. Soc. Lond. B 2002, 269, 455–460. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ingwell, L.L.; Zemetra, R.; Mallory-Smith, C.; Bosque-Pérez, N.A. Arundo Donax Infection with Barley Yellow Dwarf Virus Has Implications for Biofuel Production and Non-Managed Habitats. Biomass Bioenergy 2014, 66, 426–433. [Google Scholar] [CrossRef]
- Liu, X.D.; Zhai, B.P.; Zhang, X.X. Specialized Host-Plant Performance of the Cotton Aphid Is Altered by Experience. Ecol. Res. 2008, 23, 919–925. [Google Scholar] [CrossRef]
- De Barro, P.; Sherratt, T.; David, O.; Maclean, N. An Investigation of the Differential Performance of Clones of the Aphid Sitobion Avenae on Two Host Species. Oecologia 1995, 104, 379–385. [Google Scholar] [CrossRef]
- Caillaud, M.C.; Via, S. Specialized Feeding Behavior Influences Both Ecological Specialization and Assortative Mating in Sympatric Host Races of Pea Aphids. Am. Nat. 2000, 156, 606–621. [Google Scholar] [CrossRef]
- Komazaki, S. Variation in the Hatch Timing of the Overwintering Egg Amoung Populations of Aphis Spiraecola Patch (Homoptera: Aphididae) Collected from Different Host Plants and Localities in Japan. Appl. Entomol. Zool. 1990, 25, 27–34. [Google Scholar] [CrossRef]
- Caballero, P.P.; Ramirez, C.C.; Niemeyer, H.M. Specialisation Pattern of the Aphid Rhopalosiphum Maidis Is Not Modified by Experience on a Novel Host. Entomol. Exp. Appl. 2001, 100, 43–52. [Google Scholar] [CrossRef]
- Mackenzie, A. A Trade-off for Host Plant Utilization in the Black Bean Aphid, Aphis Fabae. Evolution 1996, 50, 155–162. [Google Scholar] [CrossRef] [PubMed]
- Stilmant, D.; Van Bellinghen, C.; Hance, T.; Boivin, G. Host Specialization in Habitat Specialists and Generalists. Oecologia 2008, 156, 905–912. [Google Scholar] [CrossRef] [PubMed]
- Gorur, G.; Lomonaco, C.; Mackenzie, A. Phenotypic Plasticity in Host-Plant Specialisation in Aphis Fabae. Ecol. Entomol. 2005, 30, 657–664. [Google Scholar] [CrossRef]
- Wu, W.; Liang, X.L.; Zhao, H.Y.; Xu, T.T.; Liu, X.D. Special Plant Species Determines Diet Breadth of Phytophagous Insects: A Study on Host Plant Expansion of the Host-Specialized Aphis Gossypii Glover. PLoS ONE 2013, 8, e60832. [Google Scholar] [CrossRef] [Green Version]
- Alkhedir, H.; Karlovsky, P.; Mashaly, A.M.A.; Vidal, S. Specialization and Host Plant Use of the Common Clones of Sitobion Avenae (Homoptera: Aphididae). Appl. Entomol. Zool. 2016, 51, 289–295. [Google Scholar] [CrossRef]
- Legarrea, S.; Barman, A.; Diffie, S.; Srinivasan, R. Virus Accumulation and Whitefly Performance Modulate the Role of Alternate Host Species as Inoculum Sources of Tomato Yellow Leaf Curl Virus. Plant Dis. 2020, 104, 2958–2966. [Google Scholar] [CrossRef]
- Artico, S.; Nardeli, S.M.; Brilhante, O.; Grossi-de-Sa, M.F.; Alves-Ferreira, M. Identification and evaluation of new reference genes in Gossypium hirsutum for accurate normalization of real-time quantitative RT-PCR data. BMC Plant Biol. 2010, 10, 49. [Google Scholar] [CrossRef] [Green Version]
- Liu, D.; Shi, L.; Han, C.; Yu, J.; Li, D.; Zhang, Y. Validation of Reference Genes for Gene Expression Studies in Virus-Infected Nicotiana benthamiana Using Quantitative Real-Time PCR. PLoS ONE 2012, 7, e46451. [Google Scholar] [CrossRef]
- Niu, X.; Chen, M.; Huang, X.; Chen, H.; Tao, A.; Xu, J.; Qi, J. Reference gene selection for qRT-PCR normalization analysis in kenaf (Hibiscus cannabinus L.) under abiotic stress and hormonal stimuli. Front. Plant Sci. 2017, 8, 771. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pupim Junior, O.; Schuster, I.; Pinto, R.B.; Pires, E.; Belot, J.L.; Silvie, P.; Chitarra, L.G.; Hoffmann, L.V.; Barroso, P. Inheritance of resistance to cotton blue disease. Pesqui. Agropecu. Bras. 2008, 43, 661–665. [Google Scholar] [CrossRef]
- Zhang, J.R.; Feng, Y.Y.; Yang, M.J.; Xiao, Y.; Liu, Y.S.; Yuan, Y.; Li, Z.; Zhang, Y.; Zhuo, M.; Zhang, J.; et al. Systematic screening and validation of reliable reference genes for qRT-PCR analysis in Okra (Abelmoschus esculentus L.). Sci. Rep. 2022, 121, 12913. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| S.No. | Primer Pair | Sequence | Amplicon Size | Purpose |
|---|---|---|---|---|
| 1 | CLRDV3675F | CCACGTAGRCGCAACAGGCGT | * CP = 310 bp [5,10] | Endpoint PCR |
| Pol3982R | CGAGGCCTCGGAGATGAACT | |||
| 2 | CLRDV-CP5L | TGGAGGACCAGGAGCTTCAA | 109 bp (This study) | qPCR |
| CLRDV-CP5R | TGCCGGGCAATCTGATAAAG | |||
| CLRDV-CP5probe | TCTCTCGGGAAGTTCCTCAG |
| Host | N | NP | PrRP | RP | PoRP | AP | LS |
|---|---|---|---|---|---|---|---|
| Cotton | 27 | 5 (2–7) a | 1 (1–6) a | 15 (6–23) a | 2 (1–11) a | 17 (10–35) a | 23 (14–39) a |
| Hibiscus | 19 | 4 (2–6) ab | 2 (1–4) a | 11 (2–15) a | 1 (1–6) a | 15 (9–20) a | 19 (13–22) b |
| Okra | 16 | 4 (3–7) ab | 1 (1–2) a | 5 (3–9) b | 2 (0–3) a | 8 (6–12) b | 13 (9–16) c |
| Palmer amaranth | 12 | 4 (2–6) ab | 2 (1–3) a | 6 (3–9) b | 2 (0–4) a | 8 (7–15) b | 13 (10–18) c |
| Prickly sida | 20 | 3 (2–6) b | 2 (1–4) a | 12 (7–17) a | 2 (1–4) a | 17 (11–25) a | 21 (13–27) b |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pandey, S.; Bag, S.; Roberts, P.; Conner, K.; Balkcom, K.S.; Price, A.J.; Jacobson, A.L.; Srinivasan, R. Prospective Alternate Hosts of an Emerging Polerovirus in Cotton Landscapes in the Southeastern United States. Viruses 2022, 14, 2249. https://doi.org/10.3390/v14102249
Pandey S, Bag S, Roberts P, Conner K, Balkcom KS, Price AJ, Jacobson AL, Srinivasan R. Prospective Alternate Hosts of an Emerging Polerovirus in Cotton Landscapes in the Southeastern United States. Viruses. 2022; 14(10):2249. https://doi.org/10.3390/v14102249
Chicago/Turabian StylePandey, Sudeep, Sudeep Bag, Phillip Roberts, Kassie Conner, Kipling S. Balkcom, Andrew J. Price, Alana L. Jacobson, and Rajagopalbabu Srinivasan. 2022. "Prospective Alternate Hosts of an Emerging Polerovirus in Cotton Landscapes in the Southeastern United States" Viruses 14, no. 10: 2249. https://doi.org/10.3390/v14102249