
Antiviral Immunoglobulins of Chicken Egg Yolk for Potential Prevention of SARS-CoV-2 Infection
 and
and
Abstract
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Materials and Methods
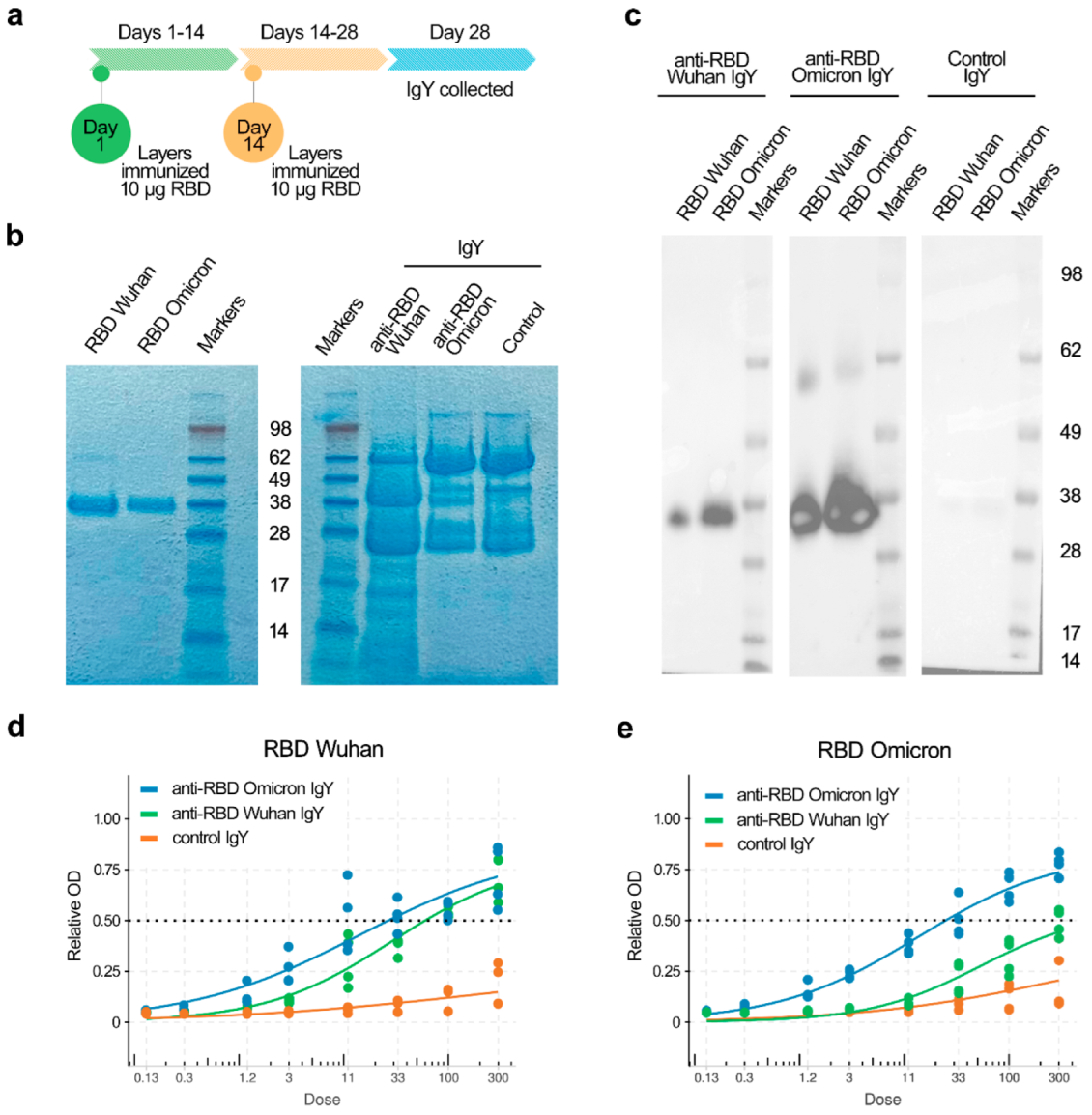
2.1. Preparation and Quantification of Anti-RBD IgY
2.2. SDS-PAGE and Immuobloting
2.3. Indirect ELISA Assay
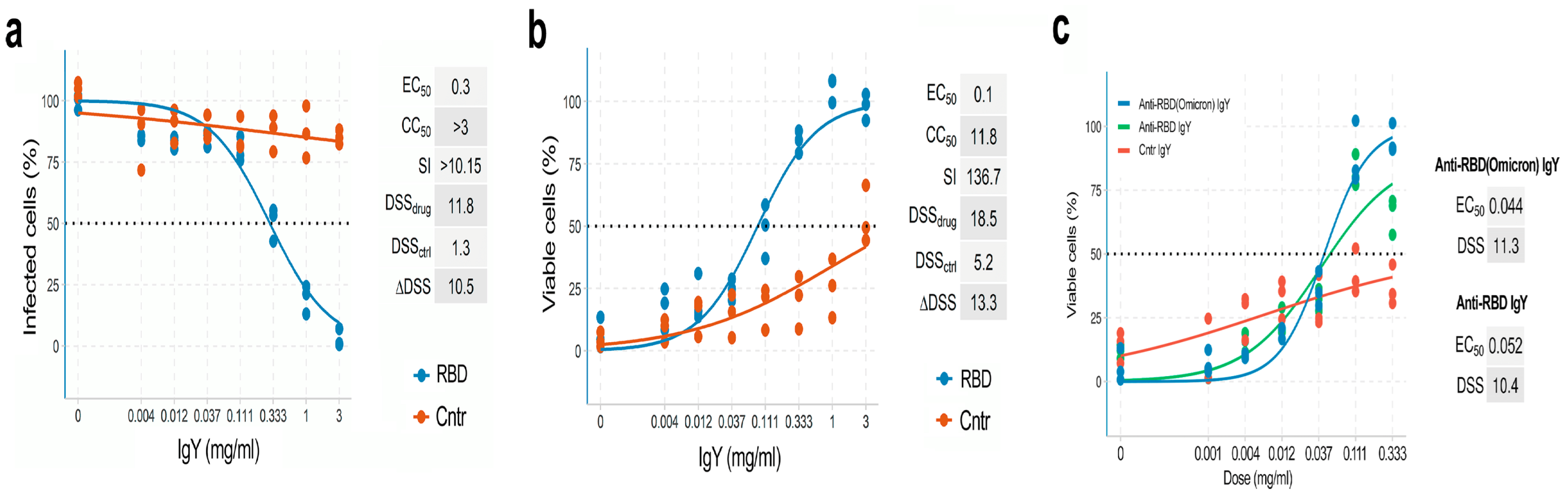
2.4. Anti-RBD IgY Testing and Data Quantification
2.5. Prophylactic Study of Anti-RBD IgY against SARS-CoV-2 Infection in Hamsters
2.6. Gene Expression Analysis
2.7. Virus TCID50 Determination
2.8. Lung Histology
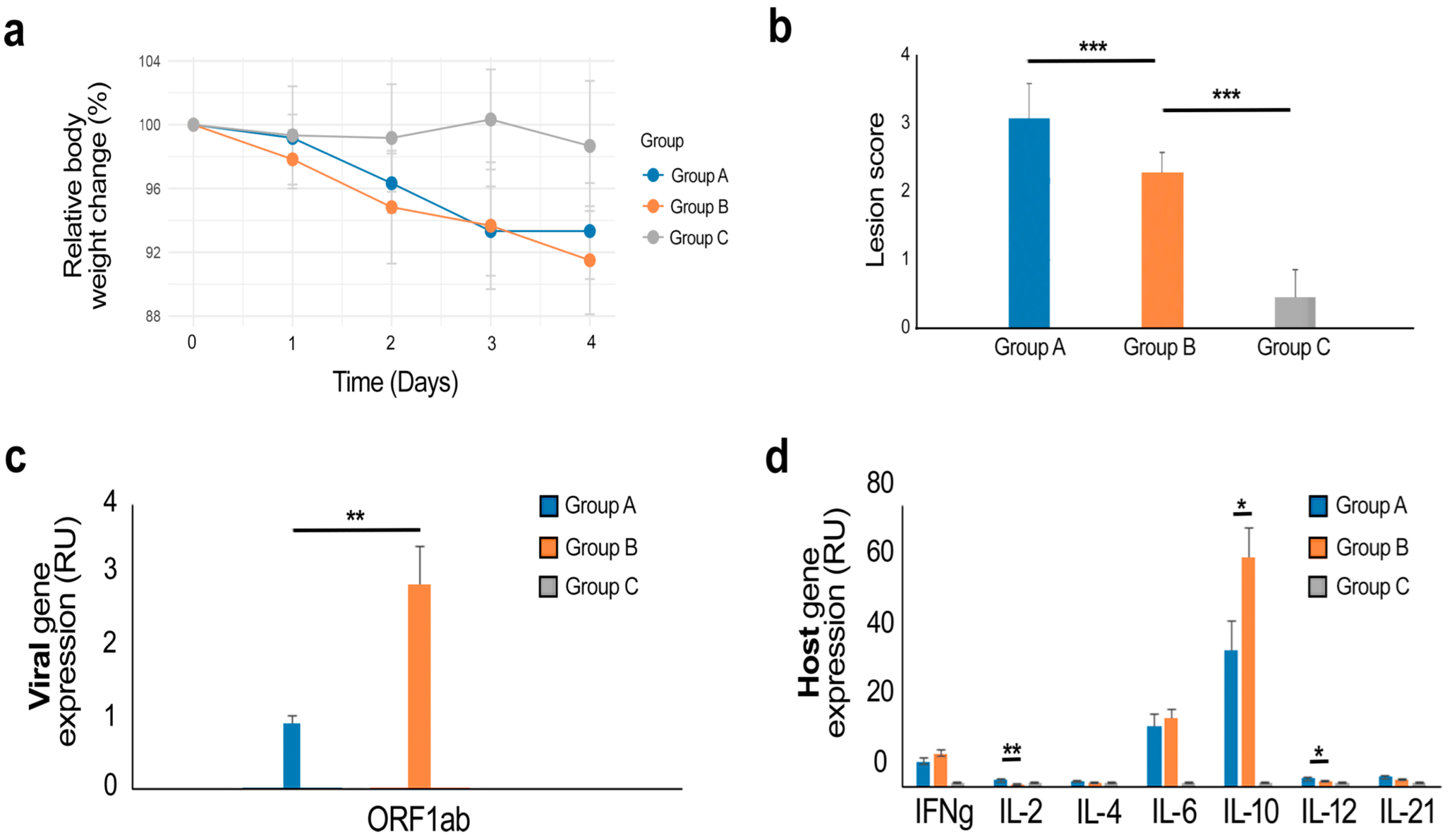
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ianevski, A.; Yao, R.; Simonsen, R.M.; Myhre, V.; Ravlo, E.; Kaynova, G.D.; Zusinaite, E.; White, J.M.; Polyak, S.J.; Oksenych, V.; et al. Mono- and combinational drug therapies for global viral pandemic preparedness. iScience 2022, 25, 104112. [Google Scholar] [CrossRef] [PubMed]
- Schmidt, F.; Weisblum, Y.; Rutkowska, M.; Poston, D.; Da Silva, J.; Zhang, F.; Bednarski, E.; Cho, A.; Schaefer-Babajew, D.J.; Gaebler, C.; et al. High genetic barrier to SARS-CoV-2 polyclonal neutralizing antibody escape. Nature 2021, 600, 512–516. [Google Scholar] [CrossRef]
- Chen, P.; Nirula, A.; Heller, B.; Gottlieb, R.L.; Boscia, J.; Morris, J.; Huhn, G.; Cardona, J.; Mocherla, B.; Stosor, V.; et al. SARS-CoV-2 Neutralizing Antibody LY-CoV555 in Outpatients with Covid-19. N. Engl. J. Med. 2021, 384, 229–237. [Google Scholar] [CrossRef]
- Weinreich, D.M.; Sivapalasingam, S.; Norton, T.; Ali, S.; Gao, H.; Bhore, R.; Musser, B.J.; Soo, Y.; Rofail, D.; Im, J.; et al. REGN-COV2, a Neutralizing Antibody Cocktail, in Outpatients with Covid-19. N. Engl. J. Med. 2021, 384, 238–251. [Google Scholar] [CrossRef] [PubMed]
- Boudewijns, R.; Thibaut, H.J.; Kaptein, S.J.F.; Li, R.; Vergote, V.; Seldeslachts, L.; Van Weyenbergh, J.; De Keyzer, C.; Bervoets, L.; Sharma, S.; et al. STAT2 signaling restricts viral dissemination but drives severe pneumonia in SARS-CoV-2 infected hamsters. Nat. Commun. 2020, 11, 5838. [Google Scholar] [CrossRef]
- Kangro, K.; Kurašin, M.; Gildemann, K.; Sankovski, E.; Žusinaite, E.; Lello, L.S.; Pert, R.; Kavak, A.; Poikalainen, V.; Lepasalu, L.; et al. Bovine colostrum-derived antibodies against SARS-CoV-2 show great potential to serve as prophylactic agents. PLoS ONE 2022, 17, e0268806. [Google Scholar] [CrossRef]
- Lee, L.; Samardzic, K.; Wallach, M.; Frumkin, L.R.; Mochly-Rosen, D. Immunoglobulin Y for Potential Diagnostic and Therapeutic Applications in Infectious Diseases. Front. Immunol. 2021, 12, 696003. [Google Scholar] [CrossRef] [PubMed]
- dda Silva, M.T.L.; Deodato, R.M.; Villar, L.M. Exploring the potential usefulness of IgY for antiviral therapy: A current review. Int. J. Biol. Macromol. 2021, 189, 785–791. [Google Scholar] [CrossRef]
- Ianevski, A.; Yao, R.; Fenstad, M.H.; Biza, S.; Zusinaite, E.; Reisberg, T.; Lysvand, H.; Løseth, K.; Landsem, V.M.; Malmring, J.F.; et al. Potential Antiviral Options against SARS-CoV-2 Infection. Viruses 2020, 12, 642. [Google Scholar] [CrossRef]
- Andersen, P.I.; Ianevski, A.; Lysvand, H.; Vitkauskiene, A.; Oksenych, V.; Bjørås, M.; Telling, K.; Lutsar, I.; Dumpis, U.; Irie, Y.; et al. Discovery and development of safe-in-man broad-spectrum antiviral agents. Int. J. Infect. Dis. 2020, 93, 268–276. [Google Scholar] [CrossRef] [PubMed]
- Andersen, P.I.; Krpina, K.; Ianevski, A.; Shtaida, N.; Jo, E.; Yang, J.; Koit, S.; Tenson, T.; Hukkanen, V.; Anthonsen, M.W.; et al. Novel Antiviral Activities of Obatoclax, Emetine, Niclosamide, Brequinar, and Homoharringtonine. Viruses 2019, 11, 964. [Google Scholar] [CrossRef] [PubMed]
- Ianevski, A.; Andersen, P.I.; Merits, A.; Bjørås, M.; Kainov, D. Expanding the activity spectrum of antiviral agents. Drug Discov. Today 2019, 24, 1224–1228. [Google Scholar] [CrossRef] [PubMed]
- Ianevski, A.; Yao, R.; Biza, S.; Zusinaite, E.; Mannik, A.; Kivi, G.; Planken, A.; Kurg, K.; Tombak, E.-M.; Ustav, M., Jr.; et al. Identification and Tracking of Antiviral Drug Combinations. Viruses 2020, 12, 1178. [Google Scholar] [CrossRef]
- Rihn, S.J.; Merits, A.; Bakshi, S.; Turnbull, M.L.; Wickenhagen, A.; Alexander, A.J.T.; Baillie, C.; Brennan, B.; Brown, F.; Brunker, K.; et al. A plasmid DNA-launched SARS-CoV-2 reverse genetics system and coronavirus toolkit for COVID-19 research. PLOS Biol. 2021, 19, e3001091. [Google Scholar] [CrossRef]
- Ianevski, A.; Yao, R.; Lysvand, H.; Grødeland, G.; Legrand, N.; Tenson, T.; Bjørås, M.; Kainov, D.E. Nafamostat-interferon-alpha combination suppresses SARS-CoV-2 infection in vitro and in vivo. bioRxiv 2021. bioRxiv:2021.06.16.448653. [Google Scholar] [CrossRef]
- Ianevski, A.; Kulesskiy, E.; Krpina, K.; Lou, G.; Aman, Y.; Bugai, A.; Aasumets, K.; Akimov, Y.; Bulanova, D.; Gildemann, K.; et al. Chemical, Physical and Biological Triggers of Evolutionary Conserved Bcl-xL-mediated Apoptosis. Cancers 2020, 12, 1694. [Google Scholar] [CrossRef] [PubMed]
- Bulanova, D.; Ianevski, A.; Bugai, A.; Akimov, Y.; Kuivanen, S.; Paavilainen, H.; Kakkola, L.; Nandania, J.; Turunen, L.; Ohman, T.; et al. Antiviral Properties of Chemical Inhibitors of Cellular Anti-Apoptotic Bcl-2 Proteins. Viruses 2017, 9, 271. [Google Scholar] [CrossRef]
- Ianevski, A.; Simonsen, R.M.; Myhre, V.; Tenson, T.; Oksenych, V.; Bjørås, M.; Kainov, D. DrugVirus.info 2.0: An integrative data portal for broad-spectrum antivirals (BSA) and BSA-containing drug combinations (BCCs). Nucleic Acids Res. 2022, 37 (Suppl. S2), W235–W239. [Google Scholar] [CrossRef]
- Potdar, S.; Ianevski, A.; Mpindi, J.-P.; Bychkov, D.; Fiere, C.; Ianevski, P.; Yadav, B.; Wennerberg, K.; Aittokallio, T.; Kallioniemi, O.; et al. Breeze: An integrated quality control and data analysis application for high-throughput drug screening. Bioinformatics 2020, 36, 3602–3604. [Google Scholar] [CrossRef]
- Yadav, B.; Pemovska, T.; Szwajda, A.; Kulesskiy, E.; Kontro, M.; Karjalainen, R.; Majumder, M.M.; Malani, D.; Murumägi, A.; Knowles, J.; et al. Quantitative scoring of differential drug sensitivity for individually optimized anticancer therapies. Sci. Rep. 2014, 4, 5193. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ianevski, A.; Yao, R.; Lysvand, H.; Grodeland, G.; Legrand, N.; Oksenych, V.; Zusinaite, E.; Tenson, T.; Bjoras, M.; Kainov, D.E. Nafamostat-Interferon-alpha Combination Suppresses SARS-CoV-2 Infection In Vitro and In Vivo by Cooperatively Targeting Host TMPRSS2. Viruses 2021, 13, 1768. [Google Scholar] [CrossRef] [PubMed]
- Frumkin, L.R.; Lucas, M.; Scribner, C.L.; Ortega-Heinly, N.; Rogers, J.; Yin, G.; Hallam, T.J.; Yam, A.; Bedard, K.; Begley, R.; et al. Egg-Derived Anti-SARS-CoV-2 Immunoglobulin Y (IgY) With Broad Variant Activity as Intranasal Prophylaxis Against COVID-19. Front. Immunol. 2022, 13, 899617. [Google Scholar] [CrossRef] [PubMed]
- Nambulli, S.; Xiang, Y.; Tilston-Lunel, N.L.; Rennick, L.J.; Sang, Z.; Klimstra, W.B.; Reed, D.S.; Crossland, N.A.; Shi, Y.; Duprex, W.P. Inhalable Nanobody (PiN-21) prevents and treats SARS-CoV-2 infections in Syrian hamsters at ultra-low doses. Sci. Adv. 2021, 7, eabh0319. [Google Scholar] [CrossRef] [PubMed]
- Agurto-Arteaga, A.; Poma-Acevedo, A.; Rios-Matos, D.; Choque-Guevara, R.; Montesinos-Millán, R.; Montalván, A.; Isasi-Rivas, G.; Cauna-Orocollo, Y.; Cauti-Mendoza, M.D.G.; Pérez-Martínez, N.; et al. Preclinical Assessment of IgY Antibodies Against Recombinant SARS-CoV-2 RBD Protein for Prophylaxis and Post-Infection Treatment of COVID-19. Front. Immunol. 2022, 13, 881604. [Google Scholar] [CrossRef] [PubMed]
- Aston, E.J.; Wallach, M.G.; Narayanan, A.; Egaña-Labrin, S.; Gallardo, R.A. Hyperimmunized Chickens Produce Neutralizing Antibodies against SARS-CoV-2. Viruses 2022, 14, 1510. [Google Scholar] [CrossRef]
- Bao, L.; Zhang, C.; Lyu, J.; Yi, P.; Shen, X.; Tang, B.; Zhao, H.; Ren, B.; Kuang, Y.; Zhou, L.; et al. Egg yolk immunoglobulin (IgY) targeting SARS-CoV-2 S1 as potential virus entry blocker. J. Appl. Microbiol. 2022, 132, 2421–2430. [Google Scholar] [CrossRef]
- Chen, C.J.; Hudson, A.F.; Jia, A.S.; Kunchur, C.R.; Song, A.J.; Tran, E.; Fisher, C.J.; Zanchi, D.; Lee, L.; Kargotich, S.; et al. Affordable IgY-based antiviral prophylaxis for resource-limited settings to address epidemic and pandemic risks. J. Glob. Health 2022, 12, 5009. [Google Scholar] [CrossRef]
- Carlander, D.; Stalberg, J.; Larsson, A. Chicken antibodies: A clinical chemistry perspective. Ups. J. Med. Sci. 1999, 104, 179–189. [Google Scholar] [CrossRef]
- Shen, H.; Cai, Y.; Zhang, H.; Wu, J.; Ye, L.; Yang, P.; Lin, X.; Jiang, S.; Liao, M. Anti-SARS-CoV-2 IgY Isolated from Egg Yolks of Hens Immunized with Inactivated SARS-CoV-2 for Immunoprophylaxis of COVID-19. Virol. Sin. 2021, 36, 1080–1082. [Google Scholar] [CrossRef]
- Wongso, H.; Mahendra, I.; Arnafia, W.; Idar, I.; Yusuf, M.; Achmad, A.; Holik, H.A.; Kurniawan, A.; Halimah, I.; Sriyani, M.E.; et al. Preclinical Evaluation of Chicken Egg Yolk Antibody (IgY) Anti-RBD Spike SARS-CoV-2—A Candidate for Passive Immunization against COVID-19. Vaccines 2022, 10, 128. [Google Scholar] [CrossRef]
- Hakalehto, E. Chicken IgY Antibodies Provide Mucosal Barrier against SARS-CoV-2 Virus and Other Pathogens. Isr. Med. Assoc. J. 2021, 23, 208–211. [Google Scholar] [PubMed]



Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ravlo, E.; Evensen, L.; Sanson, G.; Hildonen, S.; Ianevski, A.; Skjervold, P.O.; Ji, P.; Wang, W.; Kaarbø, M.; Kaynova, G.D.; et al. Antiviral Immunoglobulins of Chicken Egg Yolk for Potential Prevention of SARS-CoV-2 Infection. Viruses 2022, 14, 2121. https://doi.org/10.3390/v14102121
Ravlo E, Evensen L, Sanson G, Hildonen S, Ianevski A, Skjervold PO, Ji P, Wang W, Kaarbø M, Kaynova GD, et al. Antiviral Immunoglobulins of Chicken Egg Yolk for Potential Prevention of SARS-CoV-2 Infection. Viruses. 2022; 14(10):2121. https://doi.org/10.3390/v14102121
Chicago/Turabian StyleRavlo, Erlend, Lasse Evensen, Gorm Sanson, Siri Hildonen, Aleksandr Ianevski, Per Olav Skjervold, Ping Ji, Wei Wang, Mari Kaarbø, Gerda Dominyka Kaynova, and et al. 2022. "Antiviral Immunoglobulins of Chicken Egg Yolk for Potential Prevention of SARS-CoV-2 Infection" Viruses 14, no. 10: 2121. https://doi.org/10.3390/v14102121