
Garlic Potyviruses Are Translocated to the True Seeds through the Vegetative and Reproductive Systems of the Mother Plant

Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material and Sampling
2.2. RNA Fluorescence In Situ Hybridization
2.3. 4′,6-Diamidino-2-phenylindole (DAPI) Staining
2.4. Histology of the Reproductive Organs
2.5. Viral RNA Extraction, cDNA Synthesis, and PCR Analysis
3. Results
3.1. Calibration of FISH Technique for Virus Detection in Garlic
3.2. Identification of Viral Infection by PCR Analysis
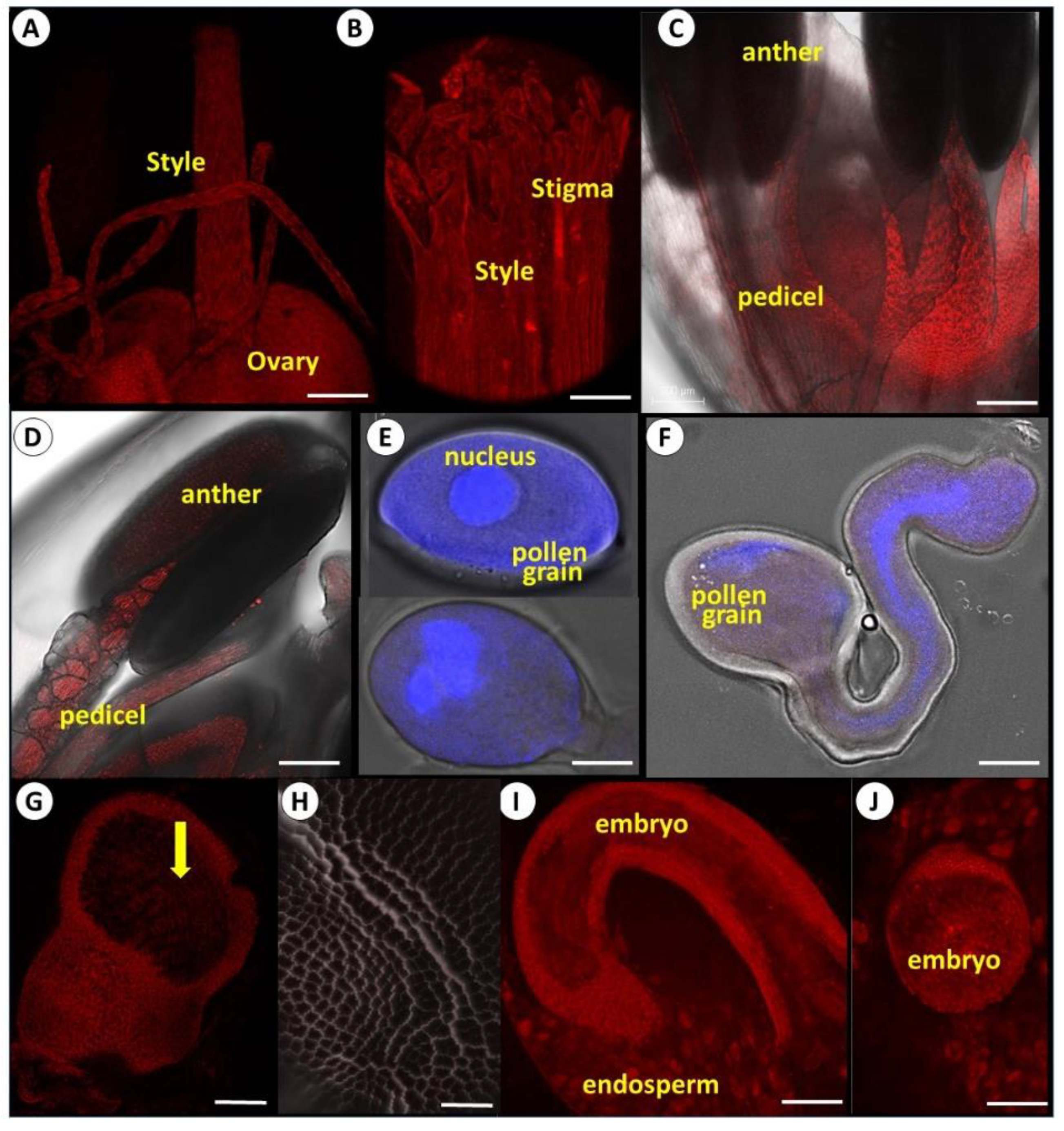
3.3. Anatomy of Reproductive Organs
3.4. FISH Identification of Potyviruses in Vegetative and Reproductive Tissues
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Fritsch, R.M.; Friesen, N. Evolution, domestication and taxonomy. In Allium Crop Sciences: Recent Advances; Rabinowitch, H.D., Currah, L., Eds.; CABI: Wallingford, UK, 2002; pp. 5–30. [Google Scholar]
- FAOStat. Food and Agriculture Organization of the United Nations. 2022. Available online: https://www.fao.org (accessed on 30 June 2022).
- Etoh, T.; Simon, P.W. Diversity, fertility and seed production of garlic. In Allium Crop Sciences: Recent Advances; Rabinowitch, H.D., Currah, L., Eds.; CABI: Wallingford, UK, 2002; pp. 101–117. [Google Scholar]
- Kamenetsky, R. Garlic: Botany and horticulture. In Horticultural Reviews; Corelli-Grapadelli, L., Kamenetsky, R., Prange, R., Eds.; John Wiley & Sons: Hoboken, NJ, USA, 2007; Volume 33, pp. 123–172. ISSN 0163-7851. [Google Scholar]
- Messiaen, C.M.; Lot, H.; Delecolle, B. Thirty years of France’experience in the production of disease-free garlic and shallot mother bulbs. In Proceedings of the International Symposium on Alliums for the Tropics, Chiang Mai, Thailand, 15–19 February 1993; Volume 358, pp. 275–280. [Google Scholar]
- Wylie, S.J.; Li, H.; Saqib, M.; Jones, M.G.K. The Global Trade in Fresh Produce and the Vagility of Plant Viruses: A Case Study in Garlic. PLoS One 2014, 9, e105044. [Google Scholar] [CrossRef] [PubMed]
- Kreuze, J.F.; Souza-Dias, J.A.C.; Jeevalatha, A.; Figueira, A.R.; Valkonen, J.P.T.; Jones, R.A.C. Viral Diseases in Potato. In The Potato Crop; Campos, H., Ortiz, O., Eds.; Springer: Cham, Switzerland, 2020; pp. 389–430. ISBN 978-3-030-28682-8. [Google Scholar]
- Salomon, R. Virus diseases in garlic and propagation of virus-free plants. In Allium Crop Sciences: Recent Advances; Rabinowitch, H.D., Currah, L., Eds.; CABI: Wallingford, UK, 2002; pp. 311–328. [Google Scholar]
- Bereda, M.; Paduch-Cichal, E. Allexiviruses–pathogens of garlic plants Allexiwirusy–patogeny czosnku pospolitego. Prog. Plant Prot. 2016, 56, 302–311. [Google Scholar] [CrossRef]
- Da Silva, L.A.; Oliveira, A.S.; Melo, F.L.; Ardisson-Araujo, D.M.; Resende, F.V.; Resende, R.O.; Ribeiro, B.M. A new virus found in garlic virus complex is a member of possible novel genus of the family Betaflexiviridae (order Tymovirales). Peer J. 2019, 7, e6285. [Google Scholar] [CrossRef] [PubMed]
- Jo, Y.; Kim, K.H.; Cho, W.K. First report of cucumber mosaic virus infecting garlic (Allium sativum L.) in Korea. Plant Pathol. J. 2021, 103, 1063–1064. [Google Scholar] [CrossRef]
- Manjunathagowda, D.C.; Gopal, J.; Archana, R.; Asiya, K.R. Virus–free seed production of garlic (Allium sativum L.): Status and prospects. Int. J. Curr. Microbial. App. Sci. 2017, 6, 2446–2456. [Google Scholar] [CrossRef]
- Desta, B.; Woldetsadik, K.; Ali, W.M. Effect of Harvesting Time, Curing and Storage Methods on Storability of Garlic Bulbs. Open Biotechnol. J. 2021, 15, 36–45. [Google Scholar] [CrossRef]
- Shemesh-Mayer, E.; Kamenetsky-Goldstein, R. Traditional and novel approaches in garlic (Allium sativum L.) breeding. In Advances in Plant Breeding Strategies: Vegetable Crops; Al-Khayri, J.M., Jain, S.M., Johnson, D.V., Eds.; Springer: Cham, Switzerland, 2021; pp. 3–49. [Google Scholar]
- Zawistowski, J.; Kopec, A.; Jędrszczyk, E.; Francik, R.; Bystrowska, B. Garlic grown from air bulbils and its potential health benefits. In Advances in Plant Phenolics: From Chemistry to Human Health; Jayaprakasha, G.K., Patil, B.S., Gattuso, G., Eds.; American Chemical Society: Washington, DC, USA, 2018; Volume 1286, pp. 315–328. [Google Scholar]
- Sulastiningsih, N.W.H.; Ashwani, N.; Hermanto, C. Agronomic characters evaluation of garlic (Allium sativum L.) bulbils. In Proceedings of the 4th International Conference on Green Agro-Industry, Grand Inna Malioboro, Indonesia, 22–23 October 2019; Volume 4, pp. 197–203. [Google Scholar]
- Ebi, M.; Kasai, N.; Masuda, K. Small inflorescence bulbils are best for micropropagation and virus elimination in garlic. HortScience 2000, 35, 735–737. [Google Scholar] [CrossRef]
- Fan, B.; He, R.; Shang, Y.; Xu, L.; Wang, N.; Gao, H.; Liu, X.; Wang, Z. System construction of virus-free and rapid-propagation technology of Baodi garlic (Allium sativum L.). Sci. Hortic. 2017, 225, 498–504. [Google Scholar] [CrossRef]
- Pappu, H.R.; Hellier, B.C.; Dugan, F.M. First report of onion yellow dwarf virus, leek yellow stripe virus, and garlic common latent virus in garlic in Washington State. Plant Dis. 2007, 89, 205. [Google Scholar] [CrossRef]
- Parrano, L.; Afunian, M.; Pagliaccia, D.; Douhan, G.; Vidalakis, G. Characterization of viruses associated with garlic plants propagated from different reproductive tissues from Italy and other geographic regions. Phytopathol. Mediterr. 2012, 51, 549–565. [Google Scholar]
- Bhusal, H.; Shemesh-Mayer, E.; Forer, I.; Kryukov, L.; Peters, R.; Kamenetsky-Goldstein, R. Bulbils in garlic inflorescence: Development and virus translocation. Sci. Hortic. 2021, 285, 110146. [Google Scholar] [CrossRef]
- Gaur, A.; Kumar, A.; Kiran, R.; Kumari, P. Importance of seed-borne diseases of agricultural crops: Economic losses and impact on society. In Seed-Borne Diseases of Agricultural Crops: Detection, Diagnosis & Management; Kumar, R., Gupta, A., Eds.; Springer: Singapore, 2020; pp. 3–23. [Google Scholar]
- Bhat, A.I.; Rao, G.P. Transmission through seeds. In Characterization of Plant Viruses; Humana: New York, NY, USA, 2020; pp. 65–67. [Google Scholar]
- Holkar, S.K.; Sandra, N.; Jain, R.K. Genetic diversity, transmission and understanding of seed-borne viruses and phytoplasma. In Seed-Borne Diseases of Agricultural Crops: Detection, Diagnosis & Management; Kumar, R., Gupta, A., Eds.; Springer: Singapore, 2020; pp. 329–353. [Google Scholar]
- Procter, C.H.; Fry, P.R. Seed transmission of tobacco mosaic virus in tomato. N. Z. J. Agric. Res. 1965, 8, 367–369. [Google Scholar] [CrossRef]
- Bennett, C.W. Seed transmission of plant viruses. Adv. Virus Res. 1969, 14, 221–261. [Google Scholar] [CrossRef] [PubMed]
- Shepherd, R.J. Transmission of viruses through seed and pollen. In Principles and Techniques in Plant Virology; Kado, C.I., Agrawal, H.O., Eds.; Van Nostrand Reinhold Co.: Washington, DC, USA, 1972; pp. 267–292. [Google Scholar]
- Hull, R. Virus plant interactions: RNA silencing. In Matthew’s Plant Virology; Academic Press: San Diego, CA, USA, 2013; pp. 477–530. [Google Scholar]
- Kumar, P.; Dhawan, P.; Mehra, R. Symptoms and losses caused by Onion yellow dwarf virus and Iris yellow spot virus diseases of onion crop in Northern India. J. Mycol. Plant Pathol. 2012, 42, 153–160. [Google Scholar]
- Ahmed, M.M.S.; Elhassan, S.M. Epidemiology and management of onion yellow dwarf disease in Sudan. Plant Pathol. J. 2013, 12, 162–168. [Google Scholar] [CrossRef]
- Louie, R.; Lorbeer, J.W. Mechanical transmission of onion yellow dwarf virus. Phytopathology 1966, 56, 1020–1023. [Google Scholar]
- Ibrahim, L.M.; Awad, M.A.E.; Abou-Zeid, A.A.; Gamal-Elin, A.S. Isolation and identification of Onion yellow dwarf virus in Egypt. J. Appl. Sci. Res. 1996, 11, 184–196. [Google Scholar]
- Abd El-Wahab, A.S. Aphid-transmission efficiency of two main viruses on garlic in Egypt, Onion yellow dwarf virus (OYDV-G) and Leek yellow stripe virus (LYSV-G). Acad. J. Entomol. 2009, 2, 40–42. [Google Scholar]
- Kritzman, A.; Lampel, M.; Raccah, B.; Gera, A. Distribution and transmission of Iris yellow spot virus. Plant Dis. 2001, 85, 838–842. [Google Scholar] [CrossRef]
- Bulajić, A.; Djekić, I.; Jović, J.; Krnjajić, S.; Vučurović, A.; Krstić, B. Incidence and distribution of Iris yellow spot virus on onion in Serbia. Plant Dis. 2009, 93, 976–982. [Google Scholar] [CrossRef]
- Shargil, D.; Zemach, H.; Belausov, E.; Lachman, O.; Luria, N.; Molad, O.; Smith, E.; Kamenetsky, R.; Dombrovsky, A. Insights into the maternal pathway for Cucumber green mottle mosaic virus infection of cucurbit seeds. Protoplasma 2019, 256, 1109–1118. [Google Scholar] [CrossRef] [PubMed]
- Gottlieb, Y.; Ghanim, M.; Chiel, E.; Gerling, D.; Portnoy, V.; Steinberg, S.; Tzuri, G.; Horowitz, A.R.; Belausov, E.; Mozes-Daube, N.; et al. Identification and localization of a Rickettsia sp. in Bemisia tabaci (Homoptera: Aleyrodidae). Appl. Environ. Microbiol. 2006, 72, 3646–3652. [Google Scholar] [CrossRef]
- Rotem, N.; David-Schwartz, R.; Peretz, Y.; Sela, I.; Rabinowitch, H.D.; Flaishman, M.; Kamenetsky, R. Flower development in garlic: The ups and downs of gaLFY expression. Planta 2011, 233, 1063–1072. [Google Scholar] [CrossRef]
- Zavriev, S.K. Allexivirus. In Encyclopedia of Virology, 3rd ed.; Mahy, B.W.J., Van Regenmortel, M.H.V., Eds.; Elsevier Academic Press Ltd.: Amsterdam, The Netherlands, 2008; pp. 96–98. [Google Scholar] [CrossRef]
- Mansouri, F.; Ryšánek, P. Allexivirus: Review and perspectives. Phytopathol. Mediterr. 2021, 60, 389–402. [Google Scholar] [CrossRef]
- Kamenetsky, R.; Faigenboim, A.; Shemesh Mayer, E.; Ben Michael, T.; Gershberg, C.; Kimhi, S.; Esquira, I.; Rohkin Shalom, S.; Eshel, D.; Rabinowitch, H.D.; et al. Integrated transcriptome catalogue and organ-specific profiling of gene expression in fertile garlic (Allium sativum L.). BMC Genom. 2015, 16, 12. [Google Scholar] [CrossRef] [PubMed]
- Navarro, J.A.; Sanchez-Navarro, J.A.; Pallas, V. Key checkpoints in the movement of plant viruses through the host. Adv. Virus Res. 2019, 104, 1–64. [Google Scholar] [CrossRef]
- Wang, A. Cell-to-cell movement of plant viruses via plasmodesmata: A current perspective on potyviruses. Curr. Opin. Virol. 2021, 48, 10–16. [Google Scholar] [CrossRef]
- Tilsner, J.; Linnik, O.; Louveaux, M.; Roberts, I.M.; Chapman, S.N.; Oparka, K.J. Replication and trafficking of a plant virus are coupled at the entrances of plasmodesmata. J. Cell Biol. 2013, 201, 981–995. [Google Scholar] [CrossRef]
- Albrechtsen, S.E. Testing Methods for Seed-transmitted Viruses: Principles and Protocols; CABI: Wallingford, UK, 2006; p. 268. ISBN 0851990169. [Google Scholar]
- Roberts, I.M.; Wang, D.; Thomas, C.L.; Maule, A.J. Pea seed-borne mosaic virus seed transmission exploits novel symplastic pathways to infect the pea embryo and is, in part, dependent upon chance. Protoplasma 2003, 222, 31–43. [Google Scholar] [CrossRef]
- Manglli, A.; Tomassoli, L.; Tiberini, A.; Agosteo, G.E.; Fontana, A.; Pappu, H.R.; Albanese, G. A survey on the infection of Onion yellow dwarf virus and Iris yellow spot tospovirus in seed and bulb productions systems of onion in Calabria, Italy. Eur. J. Plant Pathol. 2020, 156, 767–778. [Google Scholar] [CrossRef]
- Bhat, A.I.; Rao, G.P. Virus transmission through pollen. In Characterization of Plant Viruses; Bhat, A.I., Rao, G.P., Eds.; Humana: New York, NY, USA, 2020; pp. 61–64. ISBN 978-1-0716-0333-8. [Google Scholar]
- Ashurmetov, O.A.; Yengalycheva, S.S.; Fritsch, R.M. Morphological and embryological characters of three middle Asian allium L. species (Alliaceae). Bot. J. Linn. 2001, 137, 51–64. [Google Scholar] [CrossRef]
- Lucas, W.J. Plant viral movement proteins: Agents for cell-to-cell trafficking of viral genomes. Virology 2006, 344, 169–184. [Google Scholar] [CrossRef] [PubMed]
- Schoelz, J.E.; Harries, P.A.; Nelson, R.S. Intracellular transport of plant viruses: Finding the door out of the cell. Mol. Plant 2011, 4, 813–831. [Google Scholar] [CrossRef] [PubMed]
- Card, S.D.; Pearson, M.N.; Clover, G.R.G. Plant pathogens transmitted by pollen. Australas. Plant Pathol. 2007, 36, 455–461. [Google Scholar] [CrossRef]
- Hunter, D.G.; Bowyer, J.W. Cytopathology of anthers and pollen from lettuce plants infected by lettuce mosaic virus. J. Phytopathol. 1994, 142, 107–114. [Google Scholar] [CrossRef]
- Bashir, M.; Ahmad, Z.; Ghafoor, A. Cowpea aphid-borne mosaic potyvirus: A review. Int. J. Pest Manag. 2002, 48, 155–168. [Google Scholar] [CrossRef]
- Bashar, T.; Belfrage, R.; Macneil, J.; Bernard, M.; Wang, A.M. A preliminary study on soybean mosaic virus transmission via soybean seeds. In Proceedings of the Annual Meeting of Canadian Phytopathological Society, Southwestern Ontario Region Association, London, ON, Canada, 2 November 2013. [Google Scholar]
- Avni, B. Seed and Pollen Transmission of Tomato Brown Rugose Fruit Virus (ToBRFV) in Tomato Plant. Master’s Thesis, The Hebrew University of Jerusalem, Rehovot, Israel, 2019. [Google Scholar]
- Salem, N.M.; Sulaiman, A.; Samarah, N.; Turina, M.; Vallino, M. Localization and mechanical transmission of tomato brown rugose fruit virus in tomato seeds. Plant Dis. 2022, 106, 275–281. [Google Scholar] [CrossRef]
- Da Silva, W.; Kutnjak, D.; Xu, Y.; Xu, Y.; Giovannoni, J.; Elena, S.F.; Gray, S. Transmission modes affect the population structure of potato virus Y in potato. PLoS Pathog. 2020, 16, e1008608. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Virus | Forward Primer | Reverse Primer | Amplicon |
|---|---|---|---|
| Potyvirus LYSV | 5′-GAG GAA AGT CAA TAC TTA AC-3′ | 5′-TGC TGT GTG CCT CTC CGT GTC CTC-3′ | 578 nt |
| Potyvirus OYDV | 5′-GAG GAT GCA CAA TCA AG-3′ | 5′-TGC TGT GTG CCT CTC CGT GTC CTC-3′ | 714 nt |
| Allexiviruses GarV-A, B, C, D, E, X | 5′-GAT CAC ATT CTG ATG CAT CCA CAC-3′ | 5′-CGT GAG GTC TTT GTT CAC GTC-3′ | 460 nt |
| Carlaviruses GLV, GCLV | 5′-CTG AAT CAG ATT ATG AAG CTT TTG ATG C-3′ | 5′-CAA TCA CCC AGC TGG TAT TCG TC-3′ | 949 nt |
| Stage | Genotype | ALLEXI * | CARLA ** | LYSV | OYDV |
|---|---|---|---|---|---|
| Foliage leaves | 3376 | + | - | + | + |
| 3379 | + | - | - | + | |
| Young inflorescences | 3376 | + | - | + | + |
| 3379 | + | + | + | + | |
| Mature flowers | 3376 | + | - | + | + |
| 3379 | + | - | + | + | |
| Seeds | 87 (2015) | + | + | + | + |
| 87 (2016) | + | - | - | - | |
| 87 (2017) | + | - | + | + | |
| Seedlings | 87 (2017) | + | - | + | + |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shemesh-Mayer, E.; Gelbart, D.; Belausov, E.; Sher, N.; Daus, A.; Rabinowitch, H.D.; Kamenetsky-Goldstein, R. Garlic Potyviruses Are Translocated to the True Seeds through the Vegetative and Reproductive Systems of the Mother Plant. Viruses 2022, 14, 2092. https://doi.org/10.3390/v14102092
Shemesh-Mayer E, Gelbart D, Belausov E, Sher N, Daus A, Rabinowitch HD, Kamenetsky-Goldstein R. Garlic Potyviruses Are Translocated to the True Seeds through the Vegetative and Reproductive Systems of the Mother Plant. Viruses. 2022; 14(10):2092. https://doi.org/10.3390/v14102092
Chicago/Turabian StyleShemesh-Mayer, Einat, Dana Gelbart, Eduard Belausov, Nisan Sher, Ahuva Daus, Haim D. Rabinowitch, and Rina Kamenetsky-Goldstein. 2022. "Garlic Potyviruses Are Translocated to the True Seeds through the Vegetative and Reproductive Systems of the Mother Plant" Viruses 14, no. 10: 2092. https://doi.org/10.3390/v14102092