
Co-Circulation of Chikungunya and Multiple DENV Serotypes and Genotypes, Western Indonesia 2015–2016

 ,
, 
 ,
,  , and
, and
Abstract
:1. Introduction
2. Materials and Methods
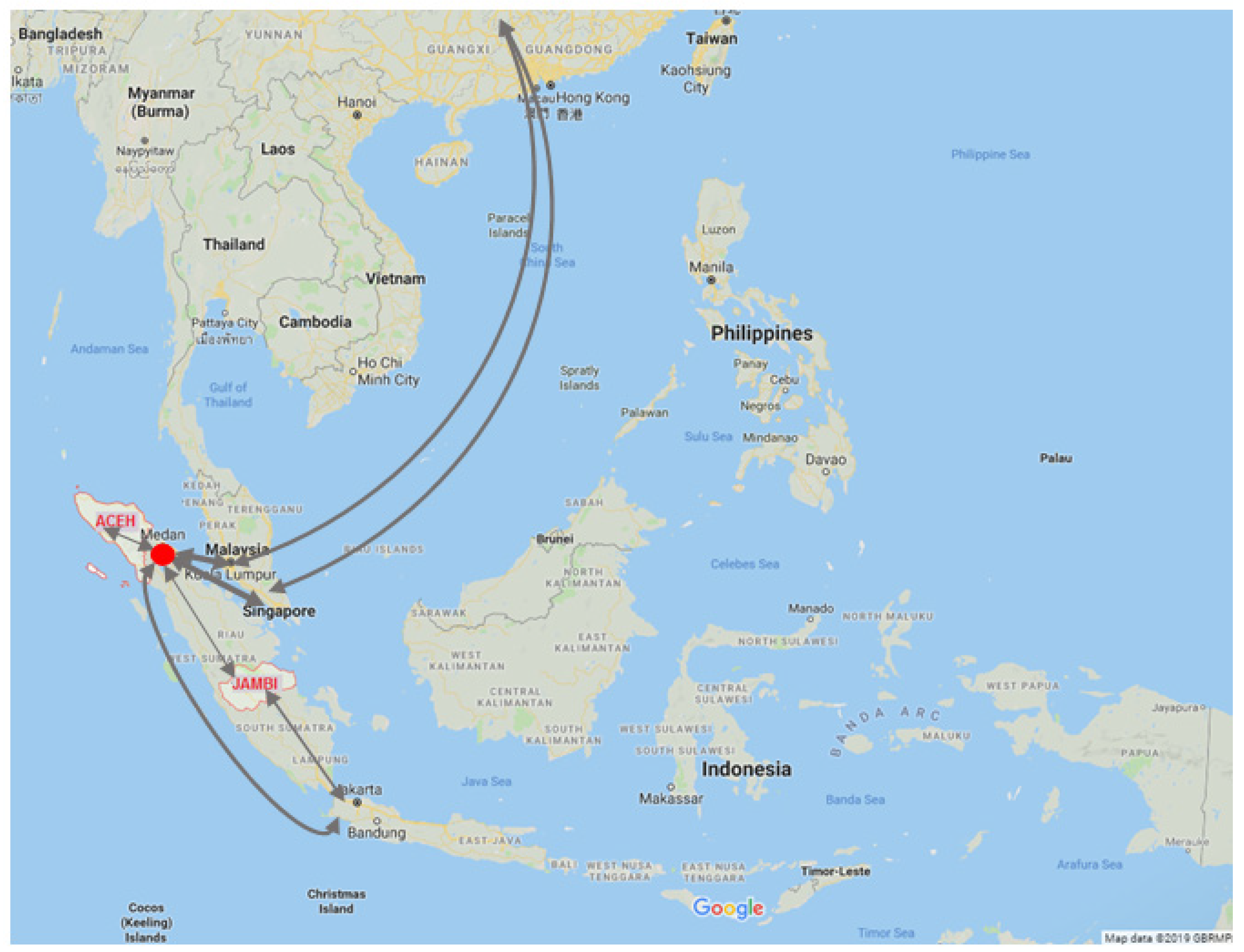
2.1. Study Setting and Patient Recruitment
2.2. NS1 EIA and DENV-Serotyping
2.3. DENV E Genes Amplification and DENV E Gene Sequencing
2.4. CHIKV Whole Genome Sequencing
2.5. Phylogenetic Analysis
3. Results
3.1. Patient Characteristics
3.2. Serotype Distributions of DENV in Aceh and Jambi
3.3. Distribution of DENV Genotypes and Lineages in Aceh and Jambi
3.3.1. DENV-1
3.3.2. DENV-2
3.3.3. DENV-3
3.4. Currently Circulating CHIKV in Aceh and Jambi
4. Discussion
4.1. Distinct DENV Lineages Circulate in Aceh and Jambi
4.2. Continuous Co-Circulation of CHIKV and DENV in Jambi
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- WHO. Global Strategy for Dengue Prevention and Control, 2012–2020; WHO Press: Geneva, Switzerland, 2012.
- Halstead, S.B. Dengue. Lancet 2007, 370, 1644–1652. [Google Scholar] [CrossRef]
- Harapan, H.; Michie, A.; Sasmono, R.T.; Imrie, A. Dengue: A minireview. Viruses 2020, 12, 829. [Google Scholar] [CrossRef]
- Prayitno, A.; Taurel, A.F.; Nealon, J.; Satari, H.I.; Karyanti, M.R.; Sekartini, R.; Soedjatmiko, S.; Gunardi, H.; Medise, B.E.; Sasmono, R.T.; et al. Dengue seroprevalence and force of primary infection in a representative population of urban dwelling Indonesian children. PLoS Negl. Trop. Dis. 2017, 11, e0005621. [Google Scholar] [CrossRef] [Green Version]
- Karyanti, M.R.; Uiterwaal, C.S.; Kusriastuti, R.; Hadinegoro, S.R.; Rovers, M.M.; Heesterbeek, H.; Hoes, A.W.; Bruijning-Verhagen, P. The changing incidence of dengue haemorrhagic fever in Indonesia: A 45-year registry-based analysis. BMC Infect. Dis. 2014, 14, 412. [Google Scholar] [CrossRef] [Green Version]
- Harapan, H.; Michie, A.; Yohan, B.; Shu, P.Y.; Mudatsir, M.; Sasmono, R.T.; Imrie, A. Dengue viruses circulating in Indonesia: A systematic review and phylogenetic analysis of data from five decades. Rev. Med. Virol. 2019, 29, e2037. [Google Scholar] [CrossRef] [PubMed]
- Harapan, H.; Michie, A.; Mudatsir, M.; Sasmono, R.T.; Imrie, A. Epidemiology of dengue hemorrhagic fever in Indonesia: Analysis of five decades data from the National Disease Surveillance. BMC Res. Notes 2019, 12, 350. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dhewantara, P.W.; Jamil, K.F.; Fajar, J.K.; Saktianggi, P.P.; Nusa, R.; Garjito, T.A.; Anwar, S.; Firzan, F.; Megawati, D.; Sasmono, R.T. Decline of notified dengue infections in Indonesia in 2017: Discussion of the possible determinants. Narra J. 2021, 1, e23. [Google Scholar] [CrossRef]
- Lestari, C.S.W.; Yohan, B.; Yunita, A.; Meutiawati, F.; Hayati, R.F.; Trimarsanto, H.; Sasmono, R.T. Phylogenetic and evolutionary analyses of dengue viruses isolated in Jakarta, Indonesia. Virus Genes 2017, 53, 778–788. [Google Scholar] [CrossRef]
- Fahri, S.; Yohan, B.; Trimarsanto, H.; Sayono, S.; Hadisaputro, S.; Dharmana, E.; Syafruddin, D.; Sasmono, R.T. Molecular surveillance of dengue in Semarang, Indonesia revealed the circulation of an old genotype of dengue virus serotype-1. PLoS Negl. Trop. Dis. 2013, 7, e2354. [Google Scholar] [CrossRef] [Green Version]
- Nusa, R.; Prasetyowati, H.; Meutiawati, F.; Yohan, B.; Trimarsanto, H.; Setianingsih, T.Y.; Sasmono, R.T. Molecular surveillance of Dengue in Sukabumi, West Java province, Indonesia. J. Infect. Dev. Ctries 2014, 8, 733–741. [Google Scholar] [CrossRef] [Green Version]
- Wardhani, P.; Aryati, A.; Yohan, B.; Trimarsanto, H.; Setianingsih, T.Y.; Puspitasari, D.; Arfijanto, M.V.; Bramantono, B.; Suharto, S.; Sasmono, R.T. Clinical and virological characteristics of dengue in Surabaya, Indonesia. PLoS ONE 2017, 12, e0178443. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yamanaka, A.; Mulyatno, K.C.; Susilowati, H.; Hendrianto, E.; Ginting, A.P.; Sary, D.D.; Rantam, F.A.; Soegijanto, S.; Konishi, E. Displacement of the predominant dengue virus from type 2 to type 1 with a subsequent genotype shift from IV to I in Surabaya, Indonesia 2008–2010. PLoS ONE 2011, 6, e27322. [Google Scholar] [CrossRef]
- Kotaki, T.; Yamanaka, A.; Mulyatno, K.C.; Labiqah, A.; Sucipto, T.H.; Churrotin, S.; Soegijanto, S.; Konishi, E.; Kameoka, M. Phylogenetic analysis of dengue virus type 3 strains primarily isolated in 2013 from Surabaya, Indonesia. Jpn. J. Infect. Dis. 2014, 67, 227–229. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kotaki, T.; Yamanaka, A.; Mulyatno, K.C.; Churrotin, S.; Sucipto, T.H.; Labiqah, A.; Ahwanah, N.L.; Soegijanto, S.; Kameoka, M.; Konishi, E. Divergence of the dengue virus type 2 Cosmopolitan genotype associated with two predominant serotype shifts between 1 and 2 in Surabaya, Indonesia, 2008–2014. Infect. Genet. Evol. 2016, 37, 88–93. [Google Scholar] [CrossRef] [PubMed]
- Kotaki, T.; Yamanaka, A.; Mulyatno, K.C.; Churrotin, S.; Labiqah, A.; Sucipto, T.H.; Soegijanto, S.; Kameoka, M.; Konishi, E. Continuous dengue type 1 virus genotype shifts followed by co-circulation, clade shifts and subsequent disappearance in Surabaya, Indonesia, 2008–2013. Infect. Genet. Evol. 2014, 28, 48–54. [Google Scholar] [CrossRef] [PubMed]
- Kusmintarsih, E.S.; Hayati, R.F.; Turnip, O.N.; Yohan, B.; Suryaningsih, S.; Pratiknyo, H.; Denis, D.; Sasmono, R.T. Molecular characterization of dengue viruses isolated from patients in Central Java, Indonesia. J. Infect. Public Health 2018, 11, 617–625. [Google Scholar] [CrossRef] [PubMed]
- Masyeni, S.; Yohan, B.; Somia, I.K.A.; Myint, K.S.A.; Sasmono, R.T. Dengue infection in international travellers visiting Bali, Indonesia. J. Travel. Med. 2018, 25, tay061. [Google Scholar] [CrossRef] [Green Version]
- Megawati, D.; Masyeni, S.; Yohan, B.; Lestarini, A.; Hayati, R.F.; Meutiawati, F.; Suryana, K.; Widarsa, T.; Budiyasa, D.G.; Budiyasa, N.; et al. Dengue in Bali: Clinical characteristics and genetic diversity of circulating dengue viruses. PLoS Negl. Trop. Dis. 2017, 11, e0005483. [Google Scholar] [CrossRef]
- Kemenkes, R.I. Profil Kesehatan Indonesia Tahun 2011; Kementerian Kesehatan RI: Jakarta, Indonesia, 2012. [Google Scholar]
- Kemenkes, R.I. Profil Kesehatan Indonesia Tahun 2013; Kementerian Kesehatan RI: Jakarta, Indonesia, 2014. [Google Scholar]
- Kemenkes, R.I. Profil Kesehatan Indonesia Tahun 2014; Kementerian Kesehatan RI: Jakarta, Indonesia, 2015. [Google Scholar]
- Haryanto, S.; Hayati, R.F.; Yohan, B.; Sijabat, L.; Sihite, I.F.; Fahri, S.; Meutiawati, F.; Halim, J.A.; Halim, S.N.; Soebandrio, A.; et al. The molecular and clinical features of dengue during outbreak in Jambi, Indonesia in 2015. Pathog. Glob. Health 2016, 110, 119–129. [Google Scholar] [CrossRef] [Green Version]
- Porter, K.R.; Tan, R.; Istary, Y.; Suharyono, W.; Sutaryo; Widjaja, S.; Ma’Roef, C.; Listiyaningsih, E.; Kosasih, H.; Hueston, L.; et al. A serological study of Chikungunya virus transmission in Yogyakarta, Indonesia: Evidence for the first outbreak since 1982. Southeast Asian J. Trop. Med. Public Health 2004, 35, 408–415. [Google Scholar]
- Harapan, H.; Michie, A.; Mudatsir, M.; Nusa, R.; Yohan, B.; Wagner, A.L.; Sasmono, R.T.; Imrie, A. Chikungunya virus infection in Indonesia: A systematic review and evolutionary analysis. BMC Infect. Dis. 2019, 19, 243. [Google Scholar] [CrossRef]
- Riswari, S.F.; Ma’roef, C.N.; Djauhari, H.; Kosasih, H.; Perkasa, A.; Yudhaputri, F.A.; Artika, I.M.; Williams, M.; van der Ven, A.; Myint, K.S.; et al. Study of viremic profile in febrile specimens of chikungunya in Bandung, Indonesia. J. Clin. Virol. 2016, 74, 61–65. [Google Scholar] [CrossRef] [Green Version]
- Mulyatno, K.C.; Susilowati, H.; Yamanaka, A.; Soegijanto, S.; Konishi, E. Primary isolation and phylogenetic studies of Chikungunya virus from Surabaya, Indonesia. Jpn. J. Infect. Dis. 2012, 65, 92–94. [Google Scholar]
- Sari, K.; Myint, K.; Andayani, A.; Adi, P.; Dhenni, R.; Perkasa, D.; Maroef, C.; Witaria, N.; Megawati, D.; Powers, A.; et al. Chikungunya fever outbreak identified in North Bali, Indonesia. Trans. R. Soc. Trop. Med. Hyg. 2017, 111, 325–327. [Google Scholar] [CrossRef] [PubMed]
- Kosasih, H.; de Mast, Q.; Widjaja, S.; Sudjana, P.; Antonjaya, U.; Ma’roef, C.; Riswari, S.F.; Porter, K.R.; Burgess, T.H.; Alisjahbana, B.; et al. Evidence for endemic chikungunya virus infections in Bandung, Indonesia. PLoS Negl. Trop. Dis. 2013, 7, e2483. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Perkasa, A.; Yudhaputri, F.; Haryanto, S.; Hayati, R.F.; Ma’roef, C.N.; Antonjaya, U.; Yohan, B.; Myint, K.S.; Ledermann, J.P.; Rosenberg, R.; et al. Isolation of Zika Virus from Febrile Patient, Indonesia. Emerg. Infect. Dis. 2016, 22, 924–925. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sasmono, R.; Perkasa, A.; Yohan, B.; Haryanto, S.; Yudhaputri, F.A.; Hayati, R.; Maaroef, C.; Ledermann, J.; Myint, K.; Powers, A. Chikungunya deterection during dengue outbreak in Sumatera, Indonesia: Clinical manifestatation and virological profile. Am. J. Trop. Med. Hyg. 2017, 97, 1393–1398. [Google Scholar] [CrossRef]
- WHO. Dengue: Guidelines for Diagnosis, Treatment, Prevention and Control—New Edition; World Health Organization (WHO) and the Special Programme for Research and Training in Tropical Diseases (TDR): Geneva, Switzerland, 2009.
- Warrilow, D.; Northill, J.A.; Pyke, A.T. Sources of dengue viruses imported into Queensland, australia, 2002–2010. Emerg. Infect. Dis. 2012, 18, 1850–1857. [Google Scholar] [CrossRef]
- Sanger, F.; Nicklen, S.; Coulson, A.R. DNA sequencing with chain-terminating inhibitors. Proc. Natl. Acad. Sci. USA 1977, 74, 5463–5467. [Google Scholar] [CrossRef] [Green Version]
- Katoh, K.; Standley, D.M. MAFFT multiple sequence alignment software version 7: Improvements in performance and usability. Mol. Biol. Evol. 2013, 30, 772–780. [Google Scholar] [CrossRef] [Green Version]
- Kearse, M.; Moir, R.; Wilson, A.; Stones-Havas, S.; Cheung, M.; Sturrock, S.; Buxton, S.; Cooper, A.; Markowitz, S.; Duran, C.; et al. Geneious Basic: An integrated and extendable desktop software platform for the organization and analysis of sequence data. Bioinformatics 2012, 28, 1647–1649. [Google Scholar] [CrossRef] [PubMed]
- Stamatakis, A.; Hoover, P.; Rougemont, J. A rapid bootstrap algorithm for the RAxML Web servers. Syst. Biol. 2008, 57, 758–771. [Google Scholar] [CrossRef]
- Stamatakis, A. RAxML-VI-HPC: Maximum likelihood-based phylogenetic analyses with thousands of taxa and mixed models. Bioinformatics 2006, 22, 2688–2690. [Google Scholar] [CrossRef] [PubMed]
- WHO. Dengue: Guideline for Diagnosis, Treatment, Prevention, and Control. Available online: http://www.who.int/tdr/publications/documents/dengue-diagnosis.pdf (accessed on 10 October 2009).
- Aryati, A.; Trimarsanto, H.; Yohan, B.; Wardhani, P.; Fahri, S.; Sasmono, R.T. Performance of commercial dengue NS1 ELISA and molecular analysis of NS1 gene of dengue viruses obtained during surveillance in Indonesia. BMC Infect. Dis. 2013, 13, 611. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yamashita, A.; Sakamoto, T.; Sekizuka, T.; Kato, K.; Takasaki, T.; Kuroda, M. DGV: Dengue Genographic Viewer. Front. Microbiol. 2016, 7, 875. [Google Scholar] [CrossRef]
- Capeding, M.R.; Chua, M.N.; Hadinegoro, S.R.; Hussain, I.I.H.M.; Nallusamy, R.; Pitisuttithum, P.; Rusmil, K.; Thisyakorn, U.; Thomas, S.J.; Huu Tran, N.; et al. Dengue and other common causes of acute febrile illness in Asia: An active surveillance study in children. PLoS Negl. Trop. Dis. 2013, 7, e2331. [Google Scholar] [CrossRef] [Green Version]
- Kemenkes, R.I. Profil Kesehatan Indonesia Tahun 2005; Kementerian Kesehatan RI: Jakarta, Indonesia, 2007. [Google Scholar]
- Kemenkes, R.I. Pedoman Pencegahan dan Pengendalian Demam Chikungunya di Indonesia; Kementerian Kesehatan RI: Jakarta, Indonesia, 2017. [Google Scholar]
- Kemenkes, R.I. Profil Kesehatan Indonesia Tahun 2016; Kementerian Kesehatan RI: Jakarta, Indonesia, 2017. [Google Scholar]
- Kemenkes, R.I. Profil Kesehatan Indonesia Tahun 2017; Kementerian Kesehatan RI: Jakarta, Indonesia, 2018. [Google Scholar]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Characteristics | Aceh (n = 29) n (%) | Jambi (n = 27) n (%) | Total n (%) |
|---|---|---|---|
| Gender | |||
| Male | 13 (44.8) | 15 (55.6) | 28 (50.0) |
| Female | 16 (55.2) | 12 (44.4) | 28 (50.0) |
| Age (year) | |||
| 0–10 | 3 (10.3) | 9 (33.3) | 12 (21.4) |
| 11–20 | 12 (41.4) | 3 (11.1) | 15 (26.8) |
| 21–30 | 12 (41.4) | 7 (25.9) | 19 (33.9) |
| 31–40 | 2 (6.9) | 5 (18.5) | 7 (12.5) |
| 41–50 | 0 (0.0) | 2 (7.4) | 2 (3.6) |
| >50 | 0 (0.0) | 1 (3.7) | 1 (1.8) |
| Clinical symptoms (n = 43) | n= 16 | n= 26 | |
| Fever | 16 (100.0) | 27 (100.0) | 43 (100.0) |
| Nausea or vomiting | 15 (93.8) | 27 (100.0) | 42 (97.7) |
| Aches and pains | 15 (93.8) | 12 (44.4) | 27 (62.8) |
| Retro-orbital pain | 10 (62.5) | 4 (14.8) | 14 (32.6) |
| Headache | 13 (81.3) | 25 (92.6) | 38 (88.4) |
| Warming signs (n = 43) | |||
| Abdominal pain or tenderness | 8 (50.5) | 1 (3.7) | 9 (20.9) |
| Persistent vomiting | 0 (0.0) | 0 (0.0) | 0 (0.0) |
| Clinical fluid accumulation | 0 (0.0) | 0 (0.0) | 0 (0.0) |
| Mucosal bleeding | 4 (25.5) | 0 (0.0) | 4 (9.3) |
| Lethargy | 0 (0.0) | 0 (0.0) | 0 (0.0) |
| Liver enlargement | 2 (12.5) | 0 (0.0) | 2 (4.7) |
| Severe dengue signs (n = 43) | |||
| Respiratory distress | 0 (0.0) | 0 (0.0) | 0 (0.0) |
| Severe bleeding | 0 (0.0) | 0 (0.0) | 0 (0.0) |
| Impaired consciousness | 0 (0.0) | 0 (0.0) | 0 (0.0) |
| Origin | Sex | Age (Year) | Collection Date (D/M/Y) | Isolate ID | Serotype | Genotype | Lineage | E-Gene Sequence |
|---|---|---|---|---|---|---|---|---|
| Aceh | Female | 53 | 27/03/2015 | D1/IDN_Aceh_006/2015 | DENV-1 | I | L2 | Complete |
| Aceh | Female | 21 | 14/09/2015 | D1/IDN_Aceh_007/2015 | DENV-1 | I | L2 | Complete |
| Aceh | Male | 19 | 12/12/2015 | D1/IDN_Aceh_014/2015 | DENV-1 | I | L1 | Partial (227 bp) |
| Aceh | Female | 11 | 21/09/2015 | D1/IDN_Aceh_021/2015 | DENV-1 | I | L4 | Complete |
| Aceh | Female | 18 | 29/09/2015 | D1/IDN_Aceh_040/2015 | DENV-1 | I | L3 | Partial (227 bp) |
| Aceh | Male | 17 | 14/09/2015 | D2/IDN_Aceh_017/2015 | DENV-2 | Cosmopolitan | L2 | Complete |
| Aceh | Female | 27 | 19/08/2015 | D2/IDN_Aceh_036/2015 | DENV-2 | Cosmopolitan | L2 | Complete |
| Aceh | Female | 20 | 16/06/2015 | D2/IDN_Aceh_046/2015 | DENV-2 | Cosmopolitan | L2 | Complete |
| Jambi | Female | 6 | 07/11/2016 | D1/IDN_Jambi_022/2016 | DENV-1 | I | L5 | Complete |
| Jambi | Male | 33 | 08/08/2016 | D1/IDN_Jambi_031/2016 | DENV-1 | I | L5 | Complete |
| Jambi | Male | 6 | 30/06/2016 | D2/IDN_Jambi_021/2016 | DENV-2 | Cosmopolitan | L2 | Complete |
| Jambi | Male | 7 | 22/07/2016 | D2/IDN_Jambi_024/2016 | DENV-2 | Cosmopolitan | L2 | Complete |
| Jambi | Female | 35 | 22/12/2016 | D2/IDN_Jambi_025/2016 | DENV-2 | Cosmopolitan | L1 | Complete |
| Jambi | Female | 43 | 08/08/2016 | D2/IDN_Jambi_029/2016 | DENV-2 | Cosmopolitan | L2 | Complete |
| Jambi | Male | 24 | 11/08/2016 | D2/IDN_Jambi_032/2016 | DENV-2 | Cosmopolitan | L1 | Complete |
| Jambi | Male | 38 | 24/08/2016 | D2/IDN_Jambi_038/2016 | DENV-2 | Cosmopolitan | L2 | Complete |
| Jambi | Male | 23 | 16/09/2016 | D2/IDN_Jambi_043/2016 | DENV-2 | Cosmopolitan | L2 | Complete |
| Jambi | Female | 30 | 21/11/2016 | D2/IDN_Jambi_102/2016 | DENV-2 | Cosmopolitan | L2 | Complete |
| Jambi | Male | 17 | 25/09/2016 | D3/IDN_Jambi_048/2016 | DENV-3 | I | L1 | Complete |
| Jambi | Female | 9 | 25/09/2016 | D3/IDN_Jambi_049/2016 | DENV-3 | I | L1 | Partial (663 bp) |
| Jambi | Female | 3 | 07/10/2016 | D3/IDN_Jambi_056/2016 | DENV-3 | I | L1 | Complete |
| Jambi | Male | 21 | 05/12/2016 | D3/IDN_Jambi_134/2016 | DENV-3 | I | L1 | Complete |
| Sex | Age (Year) | Collection Date (D/M/Y) | ID | RT-PCR CHIKV | Genotype | Sequence |
|---|---|---|---|---|---|---|
| Male | 53 | 21/11/2016 | CHIKV/IDN_Jambi_101/2016 | Positive | Asian | Full genome |
| Female | 10 | 24/11/2016 | CHIKV/IDN_Jambi_112/2016 | Positive | Asian | Full genome |
| Female | 29 | 02/12/2016 | CHIKV/IDN_Jambi_129/2016 | Positive | Asian | Partial genome |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Harapan, H.; Michie, A.; Ernst, T.; Panta, K.; Mudatsir, M.; Yohan, B.; Haryanto, S.; McCarthy, S.; Sasmono, R.T.; Imrie, A. Co-Circulation of Chikungunya and Multiple DENV Serotypes and Genotypes, Western Indonesia 2015–2016. Viruses 2022, 14, 99. https://doi.org/10.3390/v14010099
Harapan H, Michie A, Ernst T, Panta K, Mudatsir M, Yohan B, Haryanto S, McCarthy S, Sasmono RT, Imrie A. Co-Circulation of Chikungunya and Multiple DENV Serotypes and Genotypes, Western Indonesia 2015–2016. Viruses. 2022; 14(1):99. https://doi.org/10.3390/v14010099
Chicago/Turabian StyleHarapan, Harapan, Alice Michie, Timo Ernst, Kritu Panta, Mudatsir Mudatsir, Benediktus Yohan, Sotianingsih Haryanto, Suzi McCarthy, R. Tedjo Sasmono, and Allison Imrie. 2022. "Co-Circulation of Chikungunya and Multiple DENV Serotypes and Genotypes, Western Indonesia 2015–2016" Viruses 14, no. 1: 99. https://doi.org/10.3390/v14010099