
C Proteins: Controllers of Orderly Paramyxovirus Replication and of the Innate Immune Response


Abstract
:1. Introduction
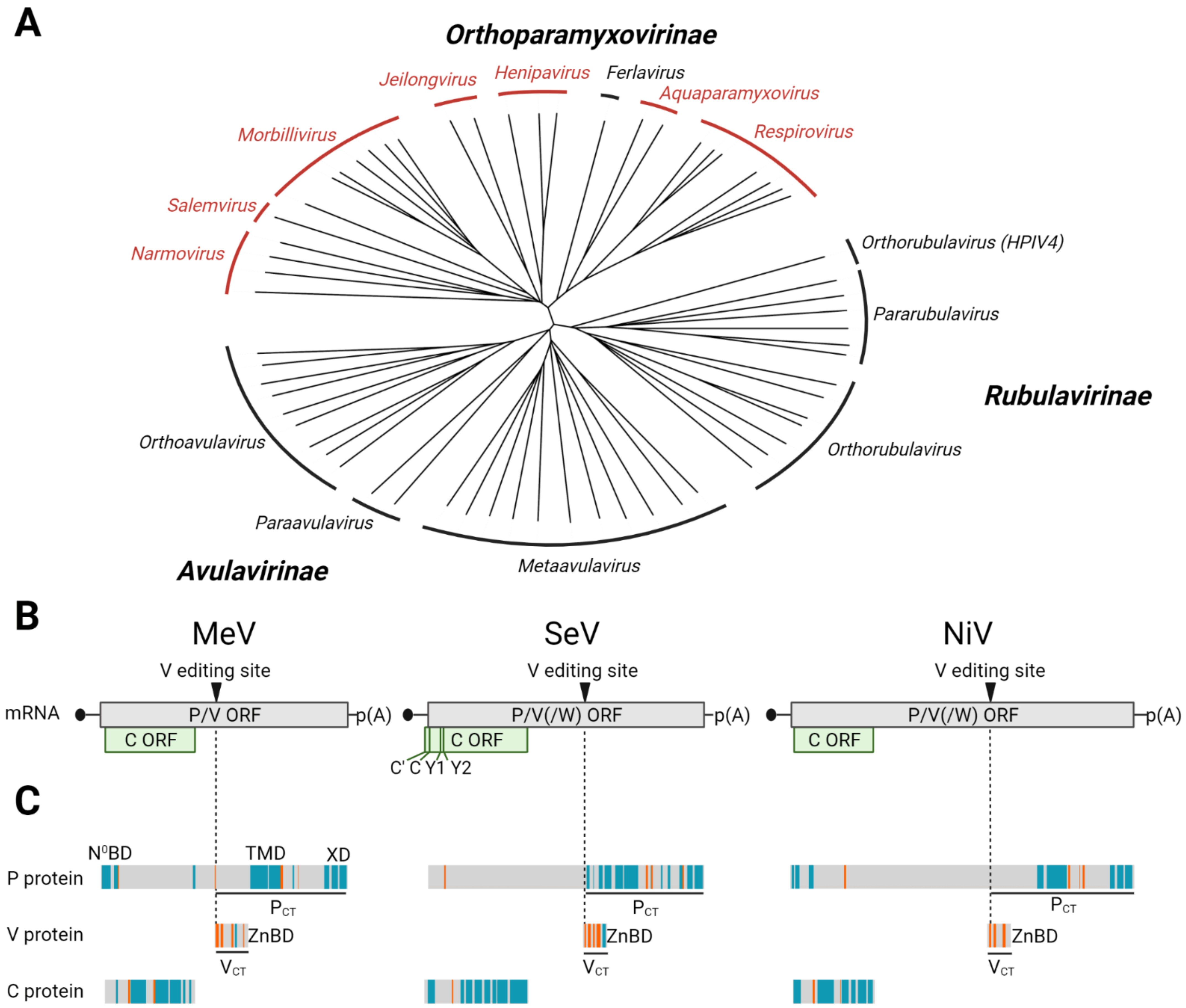
2. C Proteins Are Expressed from Alternative Open Reading Frames in the P/V/C Genes
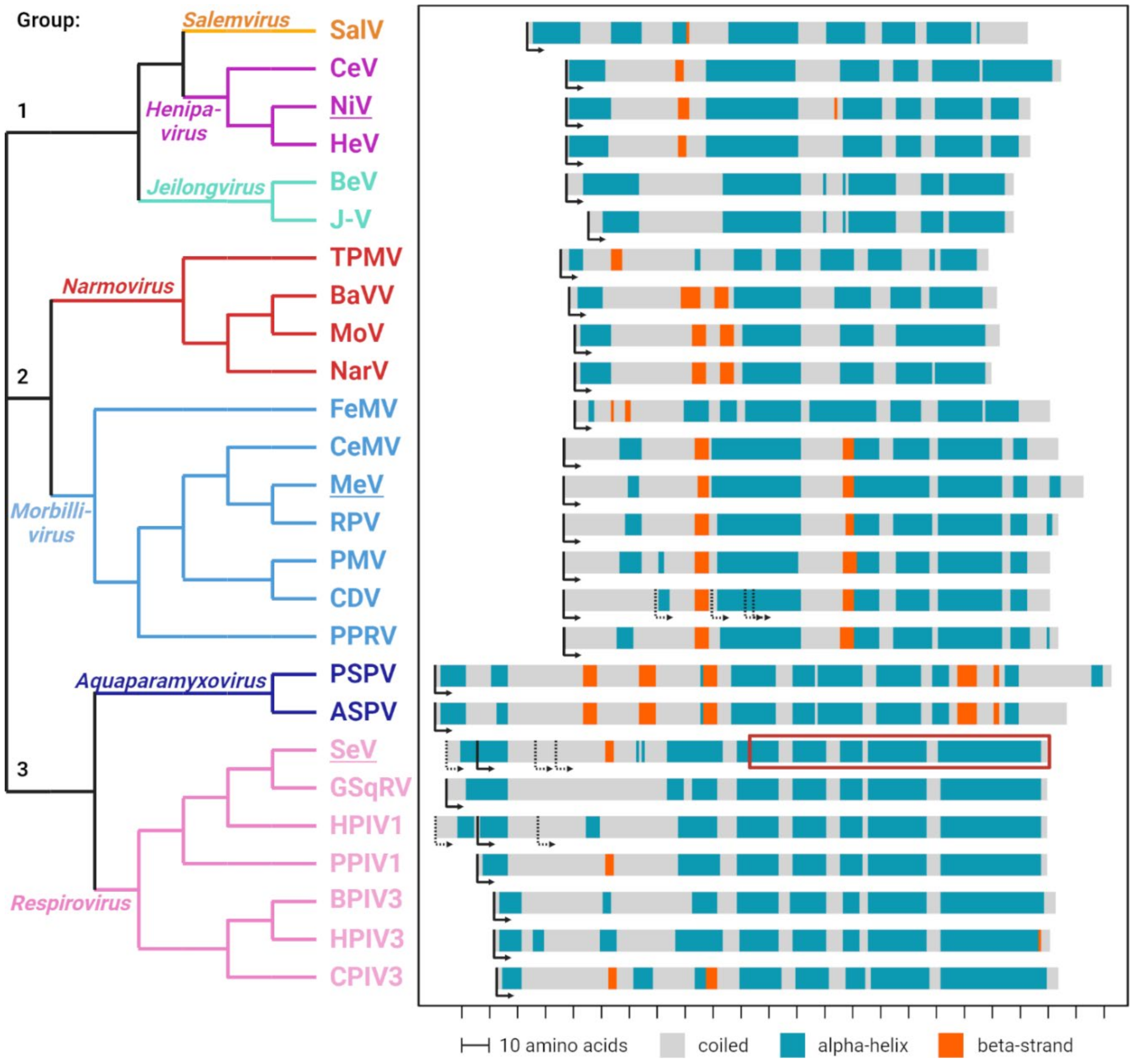
3. All Three Types of C Proteins Include an Intrinsically Disordered Part
4. Functional Insights from the SeV C-STAT1 Complex Structure
5. C Proteins Are Basic and Shuttle between Cytoplasm and Nucleus
6. C Proteins Enhance Virulence through Multiple Mechanisms
7. C Proteins Regulate Viral Transcription and Replication
8. C Proteins Minimize Production of Immunostimulatory DI RNA
9. C proteins Interact with the Polymerase Complex
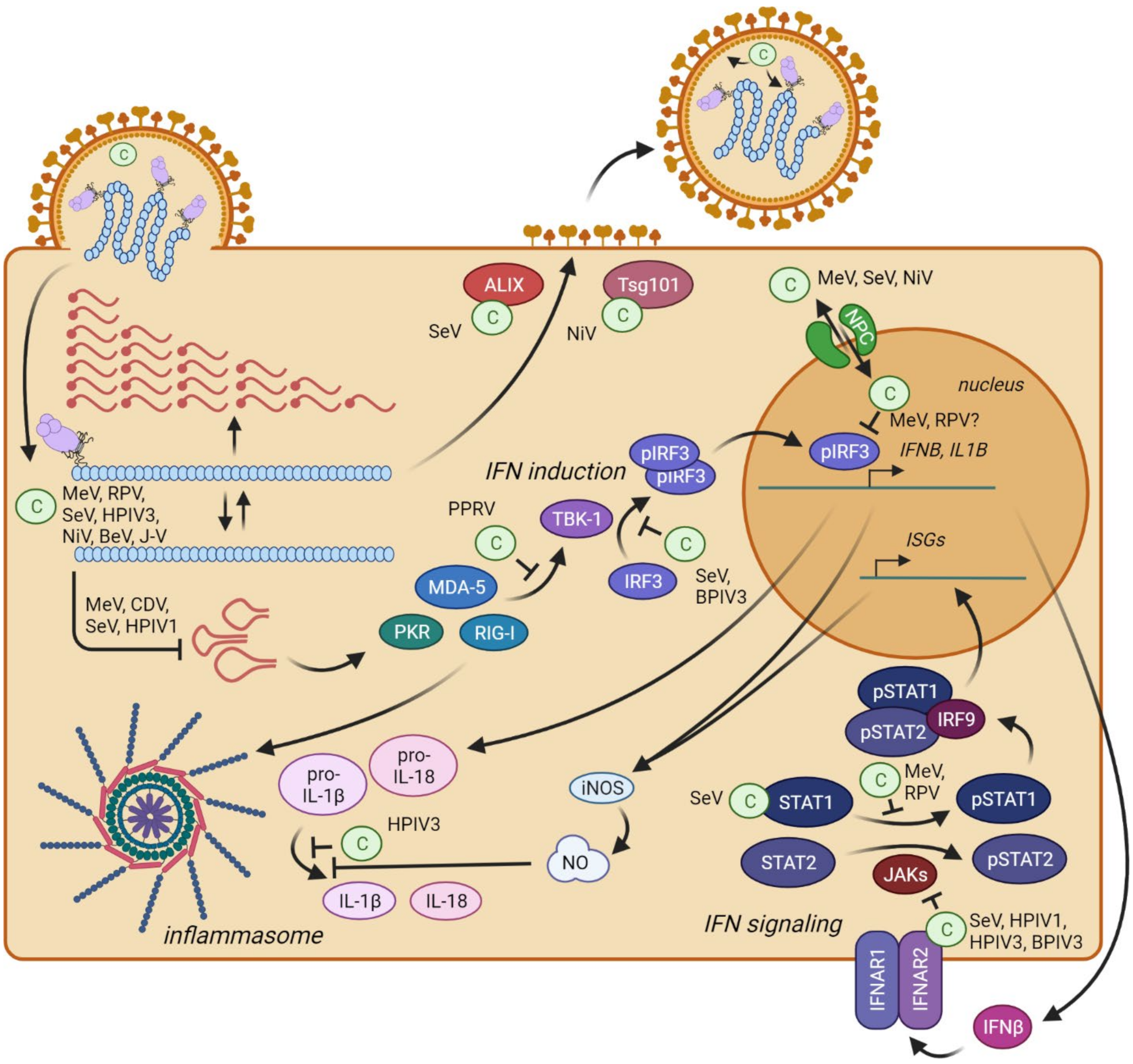
10. C Proteins Directly Interfere with Innate Immunity Activation
10.1. Interferon Induction
10.2. Interferon Signaling
10.3. Inflammation
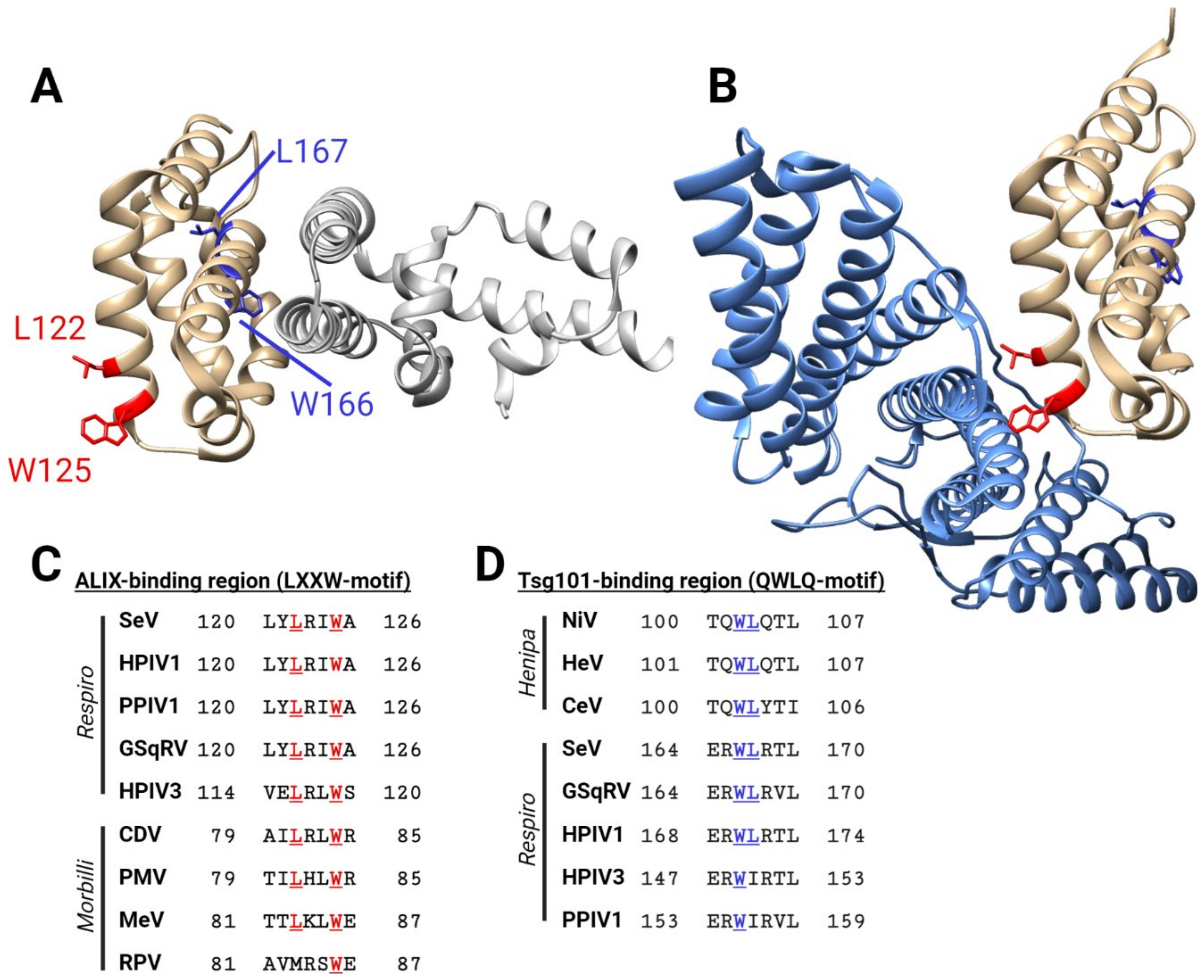
11. C Proteins Support Viral Particle Assembly and Budding
12. Concluding Remarks
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Mountcastle, W.E.; Compans, R.W.; Caliguiri, L.A.; Choppin, P.W. Nucleocapsid protein subunits of simian virus 5, Newcastle disease virus, and Sendai virus. J. Virol. 1970, 6, 677–684. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mountcastle, W.E.; Compans, R.W.; Choppin, P.W. Proteins and glycoproteins of paramyxoviruses: A comparison of simian virus 5, Newcastle disease virus, and Sendai virus. J. Virol. 1971, 7, 47–52. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Content, J.; Duesberg, P.H. Electrophoretic distribution of the proteins and glycoproteins of influenza virus and Sendai virus. J. Virol. 1970, 6, 707–716. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shimizu, K.; Hosaka, Y.; Shimizu, Y.K. Solubilization of envelopes of HVJ (Sendai virus) with alkali-emasol treatment and reassembly of envelope particles with removal of the detergent. J. Virol. 1972, 9, 842–850. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stone, H.O.; Kingsbury, D.W.; Darlington, R.W. Sendai virus-induced transcriptase from infected cells: Polypeptides in the transcriptive complex. J. Virol. 1972, 10, 1037–1043. [Google Scholar] [CrossRef] [Green Version]
- Lamb, R.A.; Parks, G. Paramyxoviridae. In Fields Virology, 6th ed.; Knipe, D.M., Howley, P.M., Eds.; Lippincott Williams & Wilkins: Philadelphia, PA, USA, 2013; pp. 957–995. [Google Scholar]
- Lamb, R.A.; Mahy, B.W.; Choppin, P.W. The synthesis of sendai virus polypeptides in infected cells. Virology 1976, 69, 116–131. [Google Scholar] [CrossRef]
- Cadd, T.; Garcin, D.; Tapparel, C.; Itoh, M.; Homma, M.; Roux, L.; Curran, J.; Kolakofsky, D. The Sendai paramyxovirus accessory C proteins inhibit viral genome amplification in a promoter-specific fashion. J. Virol. 1996, 70, 5067–5074. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Radecke, F.; Billeter, M.A. The nonstructural C protein is not essential for multiplication of Edmonston B strain measles virus in cultured cells. Virology 1996, 217, 418–421. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schneider, H.; Kaelin, K.; Billeter, M.A. Recombinant measles viruses defective for RNA editing and V protein synthesis are viable in cultured cells. Virology 1997, 227, 314–322. [Google Scholar] [CrossRef] [PubMed]
- Nagai, Y.; Kato, A. Accessory genes of the paramyxoviridae, a large family of nonsegmented negative-strand RNA viruses, as a focus of active investigation by reverse genetics. Curr. Top. Microbiol. Immunol. 2004, 283, 197–248. [Google Scholar] [CrossRef] [PubMed]
- Kato, A.; Kiyotani, K.; Sakai, Y.; Yoshida, T.; Nagai, Y. The paramyxovirus, Sendai virus, V protein encodes a luxury function required for viral pathogenesis. EMBO J. 1997, 16, 578–587. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takeuchi, K.; Takeda, M.; Miyajima, N.; Ami, Y.; Nagata, N.; Suzaki, Y.; Shahnewaz, J.; Kadota, S.-I.; Nagata, K. Stringent requirement for the C protein of wild-type measles virus for growth both in vitro and in macaques. J. Virol. 2005, 79, 7838–7844. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Von Messling, V.; Svitek, N.; Cattaneo, R. Receptor (SLAM CD150) recognition and the V protein sustain swift lymphocyte-based invasion of mucosal tissue and lymphatic organs by a morbillivirus. J. Virol. 2006, 80, 6084–6092. [Google Scholar] [CrossRef] [Green Version]
- Devaux, P.; Hodge, G.; McChesney, M.B.; Cattaneo, R. Attenuation of V- or C-defective measles viruses: Infection control by the inflammatory and interferon responses of rhesus monkeys. J. Virol. 2008, 82, 5359–5367. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dowling, P.C.; Giorgi, C.; Roux, L.; Dethlefsen, L.A.; Galantowicz, M.E.; Blumberg, B.M.; Kolakofsky, D. Molecular cloning of the 3’-proximal third of Sendai virus genome. Proc. Natl. Acad. Sci. USA 1983, 80, 5213–5216. [Google Scholar] [CrossRef] [Green Version]
- Giorgi, C.; Blumberg, B.M.; Kolakofsky, D. Sendai virus contains overlapping genes expressed from a single mRNA. Cell 1983, 35, 829–836. [Google Scholar] [CrossRef]
- Thompson, J.D.; Higgins, D.G.; Gibson, T.J. CLUSTAL W: Improving the sensitivity of progressive multiple sequence alignment through sequence weighting, position-specific gap penalties and weight matrix choice. Nucleic Acids Res.Res. 1994, 22, 4673–4680. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Letunic, I.; Bork, P. Interactive Tree Of Life (iTOL) v5: An online tool for phylogenetic tree display and annotation. Nucleic Acids Res. 2021, 49, W293–W296. [Google Scholar] [CrossRef]
- Bellini, W.J.; Englund, G.; Rozenblatt, S.; Arnheiter, H.; Richardson, C.D. Measles virus P gene codes for two proteins. J. Virol. 1985, 53, 908–919. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Harcourt, B.H.; Tamin, A.; Ksiazek, T.G.; Rollin, P.E.; Anderson, L.J.; Bellini, W.J.; Rota, P.A. Molecular characterization of Nipah virus, a newly emergent paramyxovirus. Virology 2000, 271, 334–349. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thomas, S.M.; Lamb, R.A.; Paterson, R.G. Two mRNAs that differ by two nontemplated nucleotides encode the amino coterminal proteins P and V of the paramyxovirus SV5. Cell 1988, 54, 891–902. [Google Scholar] [CrossRef]
- Cattaneo, R.; Kaelin, K.; Baczko, K.; Billeter, M.A. Measles virus editing provides an additional cysteine-rich protein. Cell 1989, 56, 759–764. [Google Scholar] [CrossRef]
- Goodbourn, S.; Randall, R.E. The regulation of type I interferon production by paramyxoviruses. J. Interferon Cytokine Res. 2009, 29, 539–547. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ramachandran, A.; Horvath, C.M. Paramyxovirus disruption of interferon signal transduction: STATus report. J. Interferon Cytokine Res. 2009, 29, 531–537. [Google Scholar] [CrossRef] [PubMed]
- Parks, G.D.; Alexander-Miller, M.A. Paramyxovirus activation and inhibition of innate immune responses. J. Mol. Biol. 2013, 425, 4872–4892. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Audsley, M.D.; Moseley, G.W. Paramyxovirus evasion of innate immunity: Diverse strategies for common targets. World J. Virol. 2013, 2, 57–70. [Google Scholar] [CrossRef] [PubMed]
- Horvath, C.M. Silencing STATs: Lessons from paramyxovirus interferon evasion. Cytokine Growth Factor Rev. 2004, 15, 117–127. [Google Scholar] [CrossRef]
- Winton, J.R.; Batts, W.N.; Powers, R.L.; Purcell, M.K. Complete Genome Sequences of the Index Isolates of Two Genotypes of Pacific Salmon Paramyxovirus. Microbiol. Resour. Announc. 2019, 8, e01521-18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Falk, K.; Batts, W.N.; Kvellestad, A.; Kurath, G.; Wiik-Nielsen, J.; Winton, J.R. Molecular characterisation of Atlantic salmon paramyxovirus (ASPV): A novel paramyxovirus associated with proliferative gill inflammation. Virus Res. 2008, 133, 218–227. [Google Scholar] [CrossRef] [PubMed]
- Marsh, G.A.; de Jong, C.; Barr, J.A.; Tachedjian, M.; Smith, C.; Middleton, D.; Yu, M.; Todd, S.; Foord, A.J.; Haring, V.; et al. Cedar virus: A novel Henipavirus isolated from Australian bats. PLoS Pathog. 2012, 8, e1002836. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.F.; Yu, M.; Hansson, E.; Pritchard, L.I.; Shiell, B.; Michalski, W.P.; Eaton, B.T. The exceptionally large genome of Hendra virus: Support for creation of a new genus within the family Paramyxoviridae. J. Virol. 2000, 74, 9972–9979. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chua, K.B.; Bellini, W.J.; Rota, P.A.; Harcourt, B.H.; Tamin, A.; Lam, S.K.; Ksiazek, T.G.; Rollin, P.E.; Zaki, S.R.; Shieh, W.; et al. Nipah virus: A recently emergent deadly paramyxovirus. Science 2000, 288, 1432–1435. [Google Scholar] [CrossRef]
- Li, Z.; Yu, M.; Zhang, H.; Magoffin, D.E.; Jack, P.J.M.; Hyatt, A.; Wang, H.-Y.; Wang, L.-F. Beilong virus, a novel paramyxovirus with the largest genome of non-segmented negative-stranded RNA viruses. Virology 2006, 346, 219–228. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jack, P.J.M.; Boyle, D.B.; Eaton, B.T.; Wang, L.-F. The complete genome sequence of J virus reveals a unique genome structure in the family Paramyxoviridae. J. Virol. 2005, 79, 10690–10700. [Google Scholar] [CrossRef] [Green Version]
- Sidhu, M.S.; Husar, W.; Cook, S.D.; Dowling, P.C.; Udem, S.A. Canine distemper terminal and intergenic non-protein coding nucleotide sequences: Completion of the entire CDV genome sequence. Virology 1993, 193, 66–72. [Google Scholar] [CrossRef] [PubMed]
- Rima, B.K.; Collin, A.M.J.; Earle, J.A.P. Completion of the sequence of a cetacean morbillivirus and comparative analysis of the complete genome sequences of four morbilliviruses. Virus Genes 2005, 30, 113–119. [Google Scholar] [CrossRef] [PubMed]
- Woo, P.C.Y.; Lau, S.K.P.; Wong, B.H.L.; Fan, R.Y.Y.; Wong, A.Y.P.; Zhang, A.J.X.; Wu, Y.; Choi, G.K.Y.; Li, K.S.M.; Hui, J.; et al. Feline morbillivirus, a previously undescribed paramyxovirus associated with tubulointerstitial nephritis in domestic cats. Proc. Natl. Acad. Sci. USA 2012, 109, 5435–5440. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takeuchi, K.; Miyajima, N.; Kobune, F.; Tashiro, M. Comparative nucleotide sequence analyses of the entire genomes of B95a cell-isolated and vero cell-isolated measles viruses from the same patient. Virus Genes 2000, 20, 253–257. [Google Scholar] [CrossRef] [PubMed]
- De Vries, R.D.; Verburgh, R.J.; van de Bildt, M.W.G.; Osterhaus, A.D.M.E.; de Swart, R.L. Complete Genome Sequence of Phocine Distemper Virus Isolated from a Harbor Seal (Phoca vitulina) during the 1988 North Sea Epidemic. Genome Announc 2013, 1, e00291-13. [Google Scholar] [CrossRef] [Green Version]
- Baron, M.D.; Kamata, Y.; Barras, V.; Goatley, L.; Barrett, T. The genome sequence of the virulent Kabete ‘O’ strain of rinderpest virus: Comparison with the derived vaccine. J. Gen. Virol. 1996, 77 Pt 12, 3041–3046. [Google Scholar] [CrossRef] [PubMed]
- Bailey, D.; Banyard, A.; Dash, P.; Ozkul, A.; Barrett, T. Full genome sequence of peste des petits ruminants virus, a member of the Morbillivirus genus. Virus Res. 2005, 110, 119–124. [Google Scholar] [CrossRef] [PubMed]
- Miller, P.J.; Boyle, D.B.; Eaton, B.T.; Wang, L.-F. Full-length genome sequence of Mossman virus, a novel paramyxovirus isolated from rodents in Australia. Virology 2003, 317, 330–344. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alkhovsky, S.; Butenko, A.; Eremyan, A.; Shchetinin, A. Genetic characterization of bank vole virus (BaVV), a new paramyxovirus isolated from kidneys of bank voles in Russia. Arch. Virol. 2018, 163, 755–759. [Google Scholar] [CrossRef] [PubMed]
- Lambeth, L.S.; Yu, M.; Anderson, D.E.; Crameri, G.; Eaton, B.T.; Wang, L.-F. Complete genome sequence of Nariva virus, a rodent paramyxovirus. Arch. Virol. 2009, 154, 199–207. [Google Scholar] [CrossRef]
- Tidona, C.A.; Kurz, H.W.; Gelderblom, H.R.; Darai, G. Isolation and molecular characterization of a novel cytopathogenic paramyxovirus from tree shrews. Virology 1999, 258, 425–434. [Google Scholar] [CrossRef] [Green Version]
- Bailly, J.E.; McAuliffe, J.M.; Skiadopoulos, M.H.; Collins, P.L.; Murphy, B.R. Sequence determination and molecular analysis of two strains of bovine parainfluenza virus type 3 that are attenuated for primates. Virus Genes 2000, 20, 173–182. [Google Scholar] [CrossRef]
- Yang, L.; Li, W.; Mao, L.; Hao, F.; Wang, Z.; Zhang, W.; Deng, J.; Jiang, J. Analysis on the complete genome of a novel caprine parainfluenza virus 3. Infect. Genet. Evol. 2016, 38, 29–34. [Google Scholar] [CrossRef] [PubMed]
- Newman, J.T.; Surman, S.R.; Riggs, J.M.; Hansen, C.T.; Collins, P.L.; Murphy, B.R.; Skiadopoulos, M.H. Sequence analysis of the Washington/1964 strain of human parainfluenza virus type 1 (HPIV1) and recovery and characterization of wild-type recombinant HPIV1 produced by reverse genetics. Virus Genes 2002, 24, 77–92. [Google Scholar] [CrossRef] [PubMed]
- Durbin, A.P.; McAuliffe, J.M.; Collins, P.L.; Murphy, B.R. Mutations in the C, D, and V open reading frames of human parainfluenza virus type 3 attenuate replication in rodents and primates. Virology 1999, 261, 319–330. [Google Scholar] [CrossRef] [Green Version]
- Itoh, M.; Isegawa, Y.; Hotta, H.; Homma, M. Isolation of an avirulent mutant of Sendai virus with two amino acid mutations from a highly virulent field strain through adaptation to LLC-MK2 cells. J. Gen. Virol. 1997, 78 Pt 12, 3207–3215. [Google Scholar] [CrossRef] [Green Version]
- Lau, S.K.P.; Woo, P.C.Y.; Wu, Y.; Wong, A.Y.P.; Wong, B.H.L.; Lau, C.C.Y.; Fan, R.Y.Y.; Cai, J.-P.; Tsoi, H.-W.; Chan, K.-H.; et al. Identification and characterization of a novel paramyxovirus, porcine parainfluenza virus 1, from deceased pigs. J. Gen. Virol. 2013, 94, 2184–2190. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Forth, L.F.; Konrath, A.; Klose, K.; Schlottau, K.; Hoffmann, K.; Ulrich, R.G.; Höper, D.; Pohlmann, A.; Beer, M. A Novel Squirrel Respirovirus with Putative Zoonotic Potential. Viruses 2018, 10, 373. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Anderson, D.E.; Dubovi, E.J.; Yu, M.; Wang, L.-F.; Renshaw, R.W. Genome characterization of Salem virus reveals its evolutionary intermediate status in the subfamily Paramyxovirinae. Arch. Virol. 2012, 157, 1989–1993. [Google Scholar] [CrossRef] [PubMed]
- Lo, M.K.; Søgaard, T.M.; Karlin, D.G. Evolution and structural organization of the C proteins of paramyxovirinae. PLoS ONE 2014, 9, e90003. [Google Scholar] [CrossRef] [PubMed]
- Pei, J.; Kim, B.-H.; Grishin, N.V. PROMALS3D: A tool for multiple protein sequence and structure alignments. Nucleic Acids Res. 2008, 36, 2295–2300. [Google Scholar] [CrossRef] [PubMed]
- Madeira, F.; Park, Y.m.; Lee, J.; Buso, N.; Gur, T.; Madhusoodanan, N.; Basutkar, P.; Tivey, A.R.N.; Potter, S.C.; Finn, R.D.; et al. The EMBL-EBI search and sequence analysis tools APIs in 2019. Nucleic Acids Res. 2019, 47, W636–W641. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Drozdetskiy, A.; Cole, C.; Procter, J.; Barton, G.J. JPred4: A protein secondary structure prediction server. Nucleic Acids Res. 2015, 43, W389–W394. [Google Scholar] [CrossRef]
- Oda, K.; Matoba, Y.; Irie, T.; Kawabata, R.; Fukushi, M.; Sugiyama, M.; Sakaguchi, T. Structural Basis of the Inhibition of STAT1 Activity by Sendai Virus C Protein. J. Virol. 2015, 89, 11487–11499. [Google Scholar] [CrossRef] [Green Version]
- Longhi, S.; Bloyet, L.-M.; Gianni, S.; Gerlier, D. How order and disorder within paramyxoviral nucleoproteins and phosphoproteins orchestrate the molecular interplay of transcription and replication. Cell Mol. Life Sci. 2017, 74, 3091–3118. [Google Scholar] [CrossRef] [PubMed]
- Oda, K.; Matoba, Y.; Sugiyama, M.; Sakaguchi, T. Structural Insight into the Interaction of Sendai Virus C Protein with Alix To Stimulate Viral Budding. J. Virol. 2021, 95, e0081521. [Google Scholar] [CrossRef] [PubMed]
- Boeck, R.; Curran, J.; Matsuoka, Y.; Compans, R.; Kolakofsky, D. The parainfluenza virus type 1 P/C gene uses a very efficient GUG codon to start its C’ protein. J. Virol. 1992, 66, 1765–1768. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Curran, J.A.; Richardson, C.; Kolakofsky, D. Ribosomal initiation at alternate AUGs on the Sendai virus P/C mRNA. J. Virol. 1986, 57, 684–687. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kato, A.; Ohnishi, Y.; Kohase, M.; Saito, S.; Tashiro, M.; Nagai, Y. Y2, the smallest of the Sendai virus C proteins, is fully capable of both counteracting the antiviral action of interferons and inhibiting viral RNA synthesis. J. Virol. 2001, 75, 3802–3810. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Siering, O.; Sawatsky, B.; Pfaller, C.K. C Protein is Essential for Canine Distemper Virus Virulence and Pathogenicity in Ferrets. J. Virol. 2020, 95, e01840-20. [Google Scholar] [CrossRef]
- Kato, A.; Ohnishi, Y.; Hishiyama, M.; Kohase, M.; Saito, S.; Tashiro, M.; Nagai, Y. The amino-terminal half of Sendai virus C protein is not responsible for either counteracting the antiviral action of interferons or down-regulating viral RNA synthesis. J. Virol. 2002, 76, 7114–7124. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mao, H.; Chattopadhyay, S.; Banerjee, A.K. N-terminally truncated C protein, CNDelta25, of human parainfluenza virus type 3 is a potent inhibitor of viral replication. Virology 2009, 394, 143–148. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pfaller, C.K.; Bloyet, L.-M.; Donohue, R.C.; Huff, A.L.; Bartemes, W.P.; Yousaf, I.; Urzua, E.; Clavière, M.; Zachary, M.; de Masson d’Autume, V.; et al. The C Protein Is Recruited to Measles Virus Ribonucleocapsids by the Phosphoprotein. J. Virol. 2020, 94. [Google Scholar] [CrossRef]
- Timney, B.L.; Raveh, B.; Mironska, R.; Trivedi, J.M.; Kim, S.J.; Russel, D.; Wente, S.R.; Sali, A.; Rout, M.P. Simple rules for passive diffusion through the nuclear pore complex. J. Cell Biol. 2016, 215, 57–76. [Google Scholar] [CrossRef] [PubMed]
- Portner, A.; Gupta, K.C.; Seyer, J.M.; Beachey, E.H.; Kingsbury, D.W. Localization and characterization of Sendai virus nonstructural C and C’ proteins by antibodies against synthetic peptides. Virus Res. 1986, 6, 109–121. [Google Scholar] [CrossRef]
- Lo, M.K.; Harcourt, B.H.; Mungall, B.A.; Tamin, A.; Peeples, M.E.; Bellini, W.J.; Rota, P.A. Determination of the henipavirus phosphoprotein gene mRNA editing frequencies and detection of the C, V and W proteins of Nipah virus in virus-infected cells. J. Gen. Virol. 2009, 90, 398–404. [Google Scholar] [CrossRef] [PubMed]
- Nishie, T.; Nagata, K.; Takeuchi, K. The C protein of wild-type measles virus has the ability to shuttle between the nucleus and the cytoplasm. Microbes Infect. 2007, 9, 344–354. [Google Scholar] [CrossRef] [PubMed]
- Sparrer, K.M.J.; Pfaller, C.K.; Conzelmann, K.-K. Measles virus C protein interferes with Beta interferon transcription in the nucleus. J. Virol. 2012, 86, 796–805. [Google Scholar] [CrossRef] [Green Version]
- Irie, T.; Yoshida, A.; Sakaguchi, T. Clustered basic amino acids of the small sendai virus C protein Y1 are critical to its RAN GTPase-mediated nuclear localization. PLoS ONE 2013, 8, e73740. [Google Scholar] [CrossRef] [Green Version]
- Horie, R.; Yoneda, M.; Uchida, S.; Sato, H.; Kai, C. Region of Nipah virus C protein responsible for shuttling between the cytoplasm and nucleus. Virology 2016, 497, 294–304. [Google Scholar] [CrossRef] [PubMed]
- Carmody, S.R.; Wente, S.R. mRNA nuclear export at a glance. J. Cell Sci. 2009, 122, 1933–1937. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schnell, M.J.; Mebatsion, T.; Conzelmann, K.K. Infectious rabies viruses from cloned cDNA. EMBO J. 1994, 13, 4195–4203. [Google Scholar] [CrossRef] [Green Version]
- Radecke, F.; Spielhofer, P.; Schneider, H.; Kaelin, K.; Huber, M.; Dötsch, C.; Christiansen, G.; Billeter, M.A. Rescue of measles viruses from cloned DNA. EMBO J. 1995, 14, 5773–5784. [Google Scholar] [CrossRef]
- Desmyter, J.; Melnick, J.L.; Rawls, W.E. Defectiveness of interferon production and of rubella virus interference in a line of African green monkey kidney cells (Vero). J. Virol. 1968, 2, 955–961. [Google Scholar] [CrossRef] [Green Version]
- Osada, N.; Kohara, A.; Yamaji, T.; Hirayama, N.; Kasai, F.; Sekizuka, T.; Kuroda, M.; Hanada, K. The genome landscape of the african green monkey kidney-derived vero cell line. DNA Res. 2014, 21, 673–683. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Garcin, D.; Itoh, M.; Kolakofsky, D. A point mutation in the Sendai virus accessory C proteins attenuates virulence for mice, but not virus growth in cell culture. Virology 1997, 238, 424–431. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kurotani, A.; Kiyotani, K.; Kato, A.; Shioda, T.; Sakai, Y.; Mizumoto, K.; Yoshida, T.; Nagai, Y. Sendai virus C proteins are categorically nonessential gene products but silencing their expression severely impairs viral replication and pathogenesis. Genes Cells 1998, 3, 111–124. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baron, M.D.; Barrett, T. Rinderpest viruses lacking the C and V proteins show specific defects in growth and transcription of viral RNAs. J. Virol. 2000, 74, 2603–2611. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bartlett, E.J.; Cruz, A.-M.; Boonyaratanakornkit, J.; Esker, J.; Castaño, A.; Skiadopoulos, M.H.; Collins, P.L.; Murphy, B.R.; Schmidt, A.C. A novel human parainfluenza virus type 1 (HPIV1) with separated P and C genes is useful for generating C gene mutants for evaluation as live-attenuated virus vaccine candidates. Vaccine 2010, 28, 767–779. [Google Scholar] [CrossRef] [Green Version]
- Escoffier, C.; Manié, S.; Vincent, S.; Muller, C.P.; Billeter, M.; Gerlier, D. Nonstructural C protein is required for efficient measles virus replication in human peripheral blood cells. J. Virol. 1999, 73, 1695–1698. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Devaux, P.; Cattaneo, R. Measles virus phosphoprotein gene products: Conformational flexibility of the P/V protein amino-terminal domain and C protein infectivity factor function. J. Virol. 2004, 78, 11632–11640. [Google Scholar] [CrossRef] [Green Version]
- Otsuki, N.; Nakatsu, Y.; Kubota, T.; Sekizuka, T.; Seki, F.; Sakai, K.; Kuroda, M.; Yamaguchi, R.; Takeda, M. The V protein of canine distemper virus is required for virus replication in human epithelial cells. PLoS ONE 2013, 8, e82343. [Google Scholar] [CrossRef]
- Valsamakis, A.; Schneider, H.; Auwaerter, P.G.; Kaneshima, H.; Billeter, M.A.; Griffin, D.E. Recombinant measles viruses with mutations in the C, V, or F gene have altered growth phenotypes in vivo. J. Virol. 1998, 72, 7754–7761. [Google Scholar] [CrossRef] [Green Version]
- Itoh, M.; Hotta, H.; Homma, M. Increased induction of apoptosis by a Sendai virus mutant is associated with attenuation of mouse pathogenicity. J. Virol. 1998, 72, 2927–2934. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kato, A.; Kiyotani, K.; Kubota, T.; Yoshida, T.; Tashiro, M.; Nagai, Y. Importance of the anti-interferon capacity of Sendai virus C protein for pathogenicity in mice. J. Virol. 2007, 81, 3264–3271. [Google Scholar] [CrossRef] [Green Version]
- Mathieu, C.; Guillaume, V.; Volchkova, V.A.; Pohl, C.; Jacquot, F.; Looi, R.Y.; Wong, K.T.; Legras-Lachuer, C.; Volchkov, V.E.; Lachuer, J.; et al. Nonstructural Nipah virus C protein regulates both the early host proinflammatory response and viral virulence. J. Virol. 2012, 86, 10766–10775. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Satterfield, B.A.; Cross, R.W.; Fenton, K.A.; Borisevich, V.; Agans, K.N.; Deer, D.J.; Graber, J.; Basler, C.F.; Geisbert, T.W.; Mire, C.E. Nipah Virus C and W Proteins Contribute to Respiratory Disease in Ferrets. J. Virol. 2016, 90, 6326–6343. [Google Scholar] [CrossRef] [Green Version]
- Curran, J.; Marq, J.B.; Kolakofsky, D. The Sendai virus nonstructural C proteins specifically inhibit viral mRNA synthesis. Virology 1992, 189, 647–656. [Google Scholar] [CrossRef]
- Latorre, P.; Cadd, T.; Itoh, M.; Curran, J.; Kolakofsky, D. The various Sendai virus C proteins are not functionally equivalent and exert both positive and negative effects on viral RNA accumulation during the course of infection. J. Virol. 1998, 72, 5984–5993. [Google Scholar] [CrossRef] [Green Version]
- Irie, T.; Nagata, N.; Yoshida, T.; Sakaguchi, T. Paramyxovirus Sendai virus C proteins are essential for maintenance of negative-sense RNA genome in virus particles. Virology 2008, 374, 495–505. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tapparel, C.; Hausmann, S.; Pelet, T.; Curran, J.; Kolakofsky, D.; Roux, L. Inhibition of Sendai virus genome replication due to promoter-increased selectivity: A possible role for the accessory C proteins. J. Virol. 1997, 71, 9588–9599. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Irie, T.; Okamoto, I.; Yoshida, A.; Nagai, Y.; Sakaguchi, T. Sendai virus C proteins regulate viral genome and antigenome synthesis to dictate the negative genome polarity. J. Virol. 2014, 88, 690–698. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reutter, G.L.; Cortese-Grogan, C.; Wilson, J.; Moyer, S.A. Mutations in the measles virus C protein that up regulate viral RNA synthesis. Virology 2001, 285, 100–109. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bankamp, B.; Fontana, J.M.; Bellini, W.J.; Rota, P.A. Adaptation to cell culture induces functional differences in measles virus proteins. Virol. J. 2008, 5, 129. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cattaneo, R.; Rebmann, G.; Schmid, A.; Baczko, K.; ter Meulen, V.; Billeter, M.A. Altered transcription of a defective measles virus genome derived from a diseased human brain. EMBO J. 1987, 6, 681–688. [Google Scholar] [CrossRef]
- Pfaller, C.K.; Radeke, M.J.; Cattaneo, R.; Samuel, C.E. Measles virus C protein impairs production of defective copyback double-stranded viral RNA and activation of protein kinase R. J. Virol. 2014, 88, 456–468. [Google Scholar] [CrossRef] [Green Version]
- Malur, A.G.; Hoffman, M.A.; Banerjee, A.K. The human parainfluenza virus type 3 (HPIV 3) C protein inhibits viral transcription. Virus Res. 2004, 99, 199–204. [Google Scholar] [CrossRef] [PubMed]
- Malur, A.G.; Wells, G.; McCoy, A.; Banerjee, A.K. Evidence for phosphorylation of human parainfluenza virus type 3 C protein: Mutant C proteins exhibit variable inhibitory activities in vitro. Virus Res. 2009, 144, 180–187. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wells, G.; Addington-Hall, M.; Malur, A.G. Mutations within the human parainfluenza virus type 3 (HPIV 3) C protein affect viral replication and host interferon induction. Virus Res. 2012, 167, 385–390. [Google Scholar] [CrossRef] [PubMed]
- Sleeman, K.; Bankamp, B.; Hummel, K.B.; Lo, M.K.; Bellini, W.J.; Rota, P.A. The C, V and W proteins of Nipah virus inhibit minigenome replication. J. Gen. Virol. 2008, 89, 1300–1308. [Google Scholar] [CrossRef]
- Magoffin, D.E.; Mackenzie, J.S.; Wang, L.-F. Genetic analysis of J-virus and Beilong virus using minireplicons. Virology 2007, 364, 103–111. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takeuchi, K.; Komatsu, T.; Kitagawa, Y.; Sada, K.; Gotoh, B. Sendai virus C protein plays a role in restricting PKR activation by limiting the generation of intracellular double-stranded RNA. J. Virol. 2008, 82, 10102–10110. [Google Scholar] [CrossRef] [Green Version]
- Toth, A.M.; Devaux, P.; Cattaneo, R.; Samuel, C.E. Protein kinase PKR mediates the apoptosis induction and growth restriction phenotypes of C protein-deficient measles virus. J. Virol. 2009, 83, 961–968. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McAllister, C.S.; Toth, A.M.; Zhang, P.; Devaux, P.; Cattaneo, R.; Samuel, C.E. Mechanisms of protein kinase PKR-mediated amplification of beta interferon induction by C protein-deficient measles virus. J. Virol. 2010, 84, 380–386. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boonyaratanakornkit, J.; Bartlett, E.; Schomacker, H.; Surman, S.; Akira, S.; Bae, Y.-S.; Collins, P.; Murphy, B.; Schmidt, A. The C proteins of human parainfluenza virus type 1 limit double-stranded RNA accumulation that would otherwise trigger activation of MDA5 and protein kinase R. J. Virol. 2011, 85, 1495–1506. [Google Scholar] [CrossRef] [Green Version]
- Pfaller, C.K.; Mastorakos, G.M.; Matchett, W.E.; Ma, X.; Samuel, C.E.; Cattaneo, R. Measles Virus Defective Interfering RNAs Are Generated Frequently and Early in the Absence of C Protein and Can Be Destabilized by Adenosine Deaminase Acting on RNA-1-Like Hypermutations. J. Virol. 2015, 89, 7735–7747. [Google Scholar] [CrossRef] [Green Version]
- Sánchez-Aparicio, M.T.; Garcin, D.; Rice, C.M.; Kolakofsky, D.; García-Sastre, A.; Baum, A. Loss of Sendai virus C protein leads to accumulation of RIG-I immunostimulatory defective interfering RNA. J. Gen. Virol. 2017, 98, 1282–1293. [Google Scholar] [CrossRef] [PubMed]
- Perrault, J. Origin and replication of defective interfering particles. Curr. Top. Microbiol. Immunol. 1981, 93, 151–207. [Google Scholar] [CrossRef] [PubMed]
- Calain, P.; Curran, J.; Kolakofsky, D.; Roux, L. Molecular cloning of natural paramyxovirus copy-back defective interfering RNAs and their expression from DNA. Virology 1992, 191, 62–71. [Google Scholar] [CrossRef]
- Yamada, H.; Hayata, S.; Omata-Yamada, T.; Taira, H.; Mizumoto, K.; Iwasaki, K. Association of the Sendai virus C protein with nucleocapsids. Arch. Virol. 1990, 113, 245–253. [Google Scholar] [CrossRef] [PubMed]
- Horikami, S.M.; Hector, R.E.; Smallwood, S.; Moyer, S.A. The Sendai virus C protein binds the L polymerase protein to inhibit viral RNA synthesis. Virology 1997, 235, 261–270. [Google Scholar] [CrossRef] [Green Version]
- Smallwood, S.; Moyer, S.A. The L polymerase protein of parainfluenza virus 3 forms an oligomer and can interact with the heterologous Sendai virus L, P and C proteins. Virology 2004, 318, 439–450. [Google Scholar] [CrossRef] [Green Version]
- Sweetman, D.A.; Miskin, J.; Baron, M.D. Rinderpest virus C and V proteins interact with the major (L) component of the viral polymerase. Virology 2001, 281, 193–204. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grogan, C.C.; Moyer, S.A. Sendai virus wild-type and mutant C proteins show a direct correlation between L polymerase binding and inhibition of viral RNA synthesis. Virology 2001, 288, 96–108. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cevik, B.; Smallwood, S.; Moyer, S.A. The L-L oligomerization domain resides at the very N-terminus of the sendai virus L RNA polymerase protein. Virology 2003, 313, 525–536. [Google Scholar] [CrossRef] [Green Version]
- Holmes, D.E.; Moyer, S.A. The phosphoprotein (P) binding site resides in the N terminus of the L polymerase subunit of sendai virus. J. Virol. 2002, 76, 3078–3083. [Google Scholar] [CrossRef] [Green Version]
- Liang, B.; Li, Z.; Jenni, S.; Rahmeh, A.A.; Morin, B.M.; Grant, T.; Grigorieff, N.; Harrison, S.C.; Whelan, S.P.J. Structure of the L Protein of Vesicular Stomatitis Virus from Electron Cryomicroscopy. Cell 2015, 162, 314–327. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nishie, T.; Nagata, K. Measles virus C protein facilitates transcription by the control of N protein-viral genomic RNA interaction in early phases of infection. Biochem. Biophys. Res. Commun. 2015, 463, 1262–1266. [Google Scholar] [CrossRef] [PubMed]
- Delpeut, S.; Noyce, R.S.; Siu, R.W.C.; Richardson, C.D. Host factors and measles virus replication. Curr. Opin. Virol. 2012, 2, 773–783. [Google Scholar] [CrossRef] [PubMed]
- Bloyet, L.-M.; Welsch, J.; Enchery, F.; Mathieu, C.; de Breyne, S.; Horvat, B.; Grigorov, B.; Gerlier, D. HSP90 Chaperoning in Addition to Phosphoprotein Required for Folding but Not for Supporting Enzymatic Activities of Measles and Nipah Virus L Polymerases. J. Virol. 2016, 90, 6642–6656. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ma, D.; George, C.X.; Nomburg, J.L.; Pfaller, C.K.; Cattaneo, R.; Samuel, C.E. Upon Infection, Cellular WD Repeat-Containing Protein 5 (WDR5) Localizes to Cytoplasmic Inclusion Bodies and Enhances Measles Virus Replication. J. Virol. 2018, 92. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ito, M.; Iwasaki, M.; Takeda, M.; Nakamura, T.; Yanagi, Y.; Ohno, S. Measles virus nonstructural C protein modulates viral RNA polymerase activity by interacting with host protein SHCBP1. J. Virol. 2013, 87, 9633–9642. [Google Scholar] [CrossRef] [Green Version]
- Ayasoufi, K.; Pfaller, C.K. Seek and hide: The manipulating interplay of measles virus with the innate immune system. Curr. Opin. Virol. 2020, 41, 18–30. [Google Scholar] [CrossRef]
- Koyama, A.H.; Irie, H.; Kato, A.; Nagai, Y.; Adachi, A. Virus multiplication and induction of apoptosis by Sendai virus: Role of the C proteins. Microbes Infect. 2003, 5, 373–378. [Google Scholar] [CrossRef]
- Komatsu, T.; Takeuchi, K.; Yokoo, J.; Gotoh, B. C and V proteins of Sendai virus target signaling pathways leading to IRF-3 activation for the negative regulation of interferon-beta production. Virology 2004, 325, 137–148. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Komatsu, T.; Takeuchi, K.; Gotoh, B. Bovine parainfluenza virus type 3 accessory proteins that suppress beta interferon production. Microbes Infect. 2007, 9, 954–962. [Google Scholar] [CrossRef] [PubMed]
- Strähle, L.; Marq, J.-B.; Brini, A.; Hausmann, S.; Kolakofsky, D.; Garcin, D. Activation of the beta interferon promoter by unnatural Sendai virus infection requires RIG-I and is inhibited by viral C proteins. J. Virol. 2007, 81, 12227–12237. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boxer, E.L.; Nanda, S.K.; Baron, M.D. The rinderpest virus non-structural C protein blocks the induction of type 1 interferon. Virology 2009, 385, 134–142. [Google Scholar] [CrossRef] [Green Version]
- Sanz Bernardo, B.; Goodbourn, S.; Baron, M.D. Control of the induction of type I interferon by Peste des petits ruminants virus. PLoS ONE 2017, 12, e0177300. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meignié, A.; Combredet, C.; Santolini, M.; Kovács, I.A.; Douché, T.; Gianetto, Q.G.; Eun, H.; Matondo, M.; Jacob, Y.; Grailhe, R.; et al. Proteomic Analysis Uncovers Measles Virus Protein C Interaction With p65-iASPP Protein Complex. Mol. Cell. Proteomics 2021, 20, 100049. [Google Scholar] [CrossRef] [PubMed]
- Schafer, S.L.; Lin, R.; Moore, P.A.; Hiscott, J.; Pitha, P.M. Regulation of type I interferon gene expression by interferon regulatory factor-3. J. Biol. Chem. 1998, 273, 2714–2720. [Google Scholar] [CrossRef] [Green Version]
- Linjie, L.; Xiaoling, S.; Xiaoxia, M.; Xin, C.; Ali, A.; Jialin, B. Peste des petits ruminants virus non-structural C protein inhibits the induction of interferon-β by potentially interacting with MAVS and RIG-I. Virus Genes 2021, 57, 60–71. [Google Scholar] [CrossRef] [PubMed]
- Schneider, W.M.; Chevillotte, M.D.; Rice, C.M. Interferon-stimulated genes: A complex web of host defenses. Annu. Rev. Immunol. 2014, 32, 513–545. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Palosaari, H.; Parisien, J.-P.; Rodriguez, J.J.; Ulane, C.M.; Horvath, C.M. STAT protein interference and suppression of cytokine signal transduction by measles virus V protein. J. Virol. 2003, 77, 7635–7644. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ramachandran, A.; Parisien, J.-P.; Horvath, C.M. STAT2 is a primary target for measles virus V protein-mediated alpha/beta interferon signaling inhibition. J. Virol. 2008, 82, 8330–8338. [Google Scholar] [CrossRef] [Green Version]
- Gotoh, B.; Takeuchi, K.; Komatsu, T.; Yokoo, J.; Kimura, Y.; Kurotani, A.; Kato, A.; Nagai, Y. Knockout of the Sendai virus C gene eliminates the viral ability to prevent the interferon-alpha/beta-mediated responses. FEBS Lett. 1999, 459, 205–210. [Google Scholar] [CrossRef] [Green Version]
- Takeuchi, K.; Komatsu, T.; Yokoo, J.; Kato, A.; Shioda, T.; Nagai, Y.; Gotoh, B. Sendai virus C protein physically associates with Stat1. Genes Cells 2001, 6, 545–557. [Google Scholar] [CrossRef] [PubMed]
- Garcin, D.; Marq, J.-B.; Strahle, L.; Le Mercier, P.; Kolakofsky, D. All four Sendai Virus C proteins bind Stat1, but only the larger forms also induce its mono-ubiquitination and degradation. Virology 2002, 295, 256–265. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Komatsu, T.; Takeuchi, K.; Yokoo, J.; Gotoh, B. Sendai virus C protein impairs both phosphorylation and dephosphorylation processes of Stat1. FEBS Lett. 2002, 511, 139–144. [Google Scholar] [CrossRef] [Green Version]
- Saito, S.; Ogino, T.; Miyajima, N.; Kato, A.; Kohase, M. Dephosphorylation failure of tyrosine-phosphorylated STAT1 in IFN-stimulated Sendai virus C protein-expressing cells. Virology 2002, 293, 205–209. [Google Scholar] [CrossRef] [Green Version]
- Garcin, D.; Curran, J.; Itoh, M.; Kolakofsky, D. Longer and shorter forms of Sendai virus C proteins play different roles in modulating the cellular antiviral response. J. Virol. 2001, 75, 6800–6807. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Garcin, D.; Marq, J.-B.; Iseni, F.; Martin, S.; Kolakofsky, D. A short peptide at the amino terminus of the Sendai virus C protein acts as an independent element that induces STAT1 instability. J. Virol. 2004, 78, 8799–8811. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gotoh, B.; Komatsu, T.; Takeuchi, K.; Yokoo, J. The C-terminal half-fragment of the Sendai virus C protein prevents the gamma-activated factor from binding to a gamma-activated sequence site. Virology 2003, 316, 29–40. [Google Scholar] [CrossRef] [Green Version]
- Mao, X.; Ren, Z.; Parker, G.N.; Sondermann, H.; Pastorello, M.A.; Wang, W.; McMurray, J.S.; Demeler, B.; Darnell, J.E.; Chen, X. Structural bases of unphosphorylated STAT1 association and receptor binding. Mol. Cell 2005, 17, 761–771. [Google Scholar] [CrossRef] [PubMed]
- Oda, K.; Oda, T.; Matoba, Y.; Sato, M.; Irie, T.; Sakaguchi, T. Structural analysis of the STAT1:STAT2 heterodimer revealed the mechanism of Sendai virus C protein-mediated blockade of type 1 interferon signaling. J. Biol. Chem. 2017, 292, 19752–19766. [Google Scholar] [CrossRef] [Green Version]
- Gotoh, B.; Takeuchi, K.; Komatsu, T. Inhibition of the gamma interferon response by a Sendai virus C protein mutant with no STAT1-binding ability. FEBS Lett. 2004, 567, 291–296. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kitagawa, Y.; Yamaguchi, M.; Kohno, M.; Sakai, M.; Itoh, M.; Gotoh, B. Respirovirus C protein inhibits activation of type I interferon receptor-associated kinases to block JAK-STAT signaling. FEBS Lett. 2020, 594, 864–877. [Google Scholar] [CrossRef] [PubMed]
- Malur, A.G.; Chattopadhyay, S.; Maitra, R.K.; Banerjee, A.K. Inhibition of STAT 1 phosphorylation by human parainfluenza virus type 3 C protein. J. Virol. 2005, 79, 7877–7882. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mao, H.; Chattopadhyay, S.; Banerjee, A.K. Domain within the C protein of human parainfluenza virus type 3 that regulates interferon signaling. Gene Expr. 2010, 15, 43–50. [Google Scholar] [CrossRef] [PubMed]
- Shaffer, J.A.; Bellini, W.J.; Rota, P.A. The C protein of measles virus inhibits the type I interferon response. Virology 2003, 315, 389–397. [Google Scholar] [CrossRef] [Green Version]
- Nanda, S.K.; Baron, M.D. Rinderpest virus blocks type I and type II interferon action: Role of structural and nonstructural proteins. J. Virol. 2006, 80, 7555–7568. [Google Scholar] [CrossRef] [Green Version]
- Takeuchi, K.; Kadota, S.; Takeda, M.; Miyajima, N.; Nagata, K. Measles virus V protein blocks interferon (IFN)-alpha/beta but not IFN-gamma signaling by inhibiting STAT1 and STAT2 phosphorylation. FEBS Lett. 2003, 545, 177–182. [Google Scholar] [CrossRef] [Green Version]
- Tisoncik, J.R.; Korth, M.J.; Simmons, C.P.; Farrar, J.; Martin, T.R.; Katze, M.G. Into the eye of the cytokine storm. Microbiol. Mol. Biol. Rev. 2012, 76, 16–32. [Google Scholar] [CrossRef] [Green Version]
- Paessler, S.; Walker, D.H. Pathogenesis of the viral hemorrhagic fevers. Annu. Rev. Pathol. 2013, 8, 411–440. [Google Scholar] [CrossRef]
- Vora, S.M.; Lieberman, J.; Wu, H. Inflammasome activation at the crux of severe COVID-19. Nat. Rev. Immunol. 2021, 21, 694–703. [Google Scholar] [CrossRef] [PubMed]
- Spel, L.; Martinon, F. Detection of viruses by inflammasomes. Curr. Opin. Virol. 2021, 46, 59–64. [Google Scholar] [CrossRef] [PubMed]
- Ogura, Y.; Sutterwala, F.S.; Flavell, R.A. The inflammasome: First line of the immune response to cell stress. Cell 2006, 126, 659–662. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tschopp, J.; Schroder, K. NLRP3 inflammasome activation: The convergence of multiple signalling pathways on ROS production? Nat. Rev. Immunol. 2010, 10, 210–215. [Google Scholar] [CrossRef]
- Mishra, B.B.; Rathinam, V.A.K.; Martens, G.W.; Martinot, A.J.; Kornfeld, H.; Fitzgerald, K.A.; Sassetti, C.M. Nitric oxide controls the immunopathology of tuberculosis by inhibiting NLRP3 inflammasome-dependent processing of IL-1β. Nat. Immunol. 2013, 14, 52–60. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Akaike, T.; Maeda, H. Nitric oxide and virus infection. Immunology 2000, 101, 300–308. [Google Scholar] [CrossRef] [Green Version]
- Uehara, E.U.; Shida, B.d.S.; de Brito, C.A. Role of nitric oxide in immune responses against viruses: Beyond microbicidal activity. Inflamm. Res. 2015, 64, 845–852. [Google Scholar] [CrossRef] [PubMed]
- Lo, M.K.; Peeples, M.E.; Bellini, W.J.; Nichol, S.T.; Rota, P.A.; Spiropoulou, C.F. Distinct and overlapping roles of Nipah virus P gene products in modulating the human endothelial cell antiviral response. PLoS ONE 2012, 7, e47790. [Google Scholar] [CrossRef] [Green Version]
- Lara, A.; Cong, Y.; Jahrling, P.B.; Mednikov, M.; Postnikova, E.; Yu, S.; Munster, V.; Holbrook, M.R. Peripheral immune response in the African green monkey model following Nipah-Malaysia virus exposure by intermediate-size particle aerosol. PLoS Negl. Trop. Dis. 2019, 13, e0007454. [Google Scholar] [CrossRef] [Green Version]
- Odkhuu, E.; Komatsu, T.; Naiki, Y.; Koide, N.; Yokochi, T. Sendai virus C protein inhibits lipopolysaccharide-induced nitric oxide production through impairing interferon-β signaling. Int. Immunopharmacol. 2014, 23, 267–272. [Google Scholar] [CrossRef]
- Odkhuu, E.; Komatsu, T.; Koide, N.; Naiki, Y.; Takeuchi, K.; Tanaka, Y.; Tsolmongyn, B.; Jambalganiin, U.; Morita, N.; Yoshida, T.; et al. Sendai virus C protein limits NO production in infected RAW264.7 macrophages. Innate Immun. 2018, 24, 430–438. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Komatsu, T.; Tanaka, Y.; Kitagawa, Y.; Koide, N.; Naiki, Y.; Morita, N.; Gotoh, B.; Yokochi, T. Sendai Virus V Protein Inhibits the Secretion of Interleukin-1β by Preventing NLRP3 Inflammasome Assembly. J. Virol. 2018, 92, e00842-18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shil, N.K.; Pokharel, S.M.; Banerjee, A.K.; Hoffman, M.; Bose, S. Inflammasome Antagonism by Human Parainfluenza Virus Type 3 C Protein. J. Virol. 2018, 92. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peeples, M.E. Paramyxovirus M Proteins. In The Paramyxoviruses; Kingsbury, D.W., Ed.; Springer: Boston, MA, USA, 1991; pp. 427–456. ISBN 978-0-306-43553-9. [Google Scholar]
- Cathomen, T.; Mrkic, B.; Spehner, D.; Drillien, R.; Naef, R.; Pavlovic, J.; Aguzzi, A.; Billeter, M.A.; Cattaneo, R. A matrix-less measles virus is infectious and elicits extensive cell fusion: Consequences for propagation in the brain. EMBO J. 1998, 17, 3899–3908. [Google Scholar] [CrossRef] [PubMed]
- Takimoto, T.; Portner, A. Molecular mechanism of paramyxovirus budding. Virus Res. 2004, 106, 133–145. [Google Scholar] [CrossRef] [PubMed]
- Cox, R.M.; Plemper, R.K. Structure and organization of paramyxovirus particles. Curr. Opin. Virol. 2017, 24, 105–114. [Google Scholar] [CrossRef] [PubMed]
- Coronel, E.C.; Murti, K.G.; Takimoto, T.; Portner, A. Human parainfluenza virus type 1 matrix and nucleoprotein genes transiently expressed in mammalian cells induce the release of virus-like particles containing nucleocapsid-like structures. J. Virol. 1999, 73, 7035–7038. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takimoto, T.; Murti, K.G.; Bousse, T.; Scroggs, R.A.; Portner, A. Role of matrix and fusion proteins in budding of Sendai virus. J. Virol. 2001, 75, 11384–11391. [Google Scholar] [CrossRef] [Green Version]
- Ciancanelli, M.J.; Basler, C.F. Mutation of YMYL in the Nipah virus matrix protein abrogates budding and alters subcellular localization. J. Virol. 2006, 80, 12070–12078. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pohl, C.; Duprex, W.P.; Krohne, G.; Rima, B.K.; Schneider-Schaulies, S. Measles virus M and F proteins associate with detergent-resistant membrane fractions and promote formation of virus-like particles. J. Gen. Virol. 2007, 88, 1243–1250. [Google Scholar] [CrossRef] [PubMed]
- Schmitt, A.P.; Leser, G.P.; Waning, D.L.; Lamb, R.A. Requirements for budding of paramyxovirus simian virus 5 virus-like particles. J. Virol. 2002, 76, 3952–3964. [Google Scholar] [CrossRef] [Green Version]
- Li, M.; Schmitt, P.T.; Li, Z.; McCrory, T.S.; He, B.; Schmitt, A.P. Mumps virus matrix, fusion, and nucleocapsid proteins cooperate for efficient production of virus-like particles. J. Virol. 2009, 83, 7261–7272. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sugahara, F.; Uchiyama, T.; Watanabe, H.; Shimazu, Y.; Kuwayama, M.; Fujii, Y.; Kiyotani, K.; Adachi, A.; Kohno, N.; Yoshida, T.; et al. Paramyxovirus Sendai virus-like particle formation by expression of multiple viral proteins and acceleration of its release by C protein. Virology 2004, 325, 1–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Patch, J.R.; Crameri, G.; Wang, L.-F.; Eaton, B.T.; Broder, C.C. Quantitative analysis of Nipah virus proteins released as virus-like particles reveals central role for the matrix protein. Virol. J. 2007, 4, 1. [Google Scholar] [CrossRef] [Green Version]
- Johnston, G.P.; Contreras, E.M.; Dabundo, J.; Henderson, B.A.; Matz, K.M.; Ortega, V.; Ramirez, A.; Park, A.; Aguilar, H.C. Cytoplasmic Motifs in the Nipah Virus Fusion Protein Modulate Virus Particle Assembly and Egress. J. Virol. 2017, 91, e02150-16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pantua, H.D.; McGinnes, L.W.; Peeples, M.E.; Morrison, T.G. Requirements for the assembly and release of Newcastle disease virus-like particles. J. Virol. 2006, 80, 11062–11073. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Park, A.; Yun, T.; Vigant, F.; Pernet, O.; Won, S.T.; Dawes, B.E.; Bartkowski, W.; Freiberg, A.N.; Lee, B. Nipah Virus C Protein Recruits Tsg101 to Promote the Efficient Release of Virus in an ESCRT-Dependent Pathway. PLoS Pathog. 2016, 12, e1005659. [Google Scholar] [CrossRef]
- Sakaguchi, T.; Kato, A.; Sugahara, F.; Shimazu, Y.; Inoue, M.; Kiyotani, K.; Nagai, Y.; Yoshida, T. AIP1/Alix is a binding partner of Sendai virus C protein and facilitates virus budding. J. Virol. 2005, 79, 8933–8941. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martin-Serrano, J.; Yarovoy, A.; Perez-Caballero, D.; Bieniasz, P.D.; Yaravoy, A. Divergent retroviral late-budding domains recruit vacuolar protein sorting factors by using alternative adaptor proteins. Proc. Natl. Acad. Sci. USA 2003, 100, 12414–12419. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Katoh, K.; Shibata, H.; Suzuki, H.; Nara, A.; Ishidoh, K.; Kominami, E.; Yoshimori, T.; Maki, M. The ALG-2-interacting protein Alix associates with CHMP4b, a human homologue of yeast Snf7 that is involved in multivesicular body sorting. J. Biol. Chem. 2003, 278, 39104–39113. [Google Scholar] [CrossRef] [Green Version]
- Peck, J.W.; Bowden, E.T.; Burbelo, P.D. Structure and function of human Vps20 and Snf7 proteins. Biochem. J. 2004, 377, 693–700. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Katzmann, D.J.; Babst, M.; Emr, S.D. Ubiquitin-dependent sorting into the multivesicular body pathway requires the function of a conserved endosomal protein sorting complex, ESCRT-I. Cell 2001, 106, 145–155. [Google Scholar] [CrossRef] [Green Version]
- Babst, M.; Katzmann, D.J.; Estepa-Sabal, E.J.; Meerloo, T.; Emr, S.D. Escrt-III: An endosome-associated heterooligomeric protein complex required for mvb sorting. Dev. Cell 2002, 3, 271–282. [Google Scholar] [CrossRef] [Green Version]
- Irie, T.; Nagata, N.; Yoshida, T.; Sakaguchi, T. Recruitment of Alix/AIP1 to the plasma membrane by Sendai virus C protein facilitates budding of virus-like particles. Virology 2008, 371, 108–120. [Google Scholar] [CrossRef] [Green Version]
- Marq, J.-B.; Brini, A.; Kolakofsky, D.; Garcin, D. Targeting of the Sendai virus C protein to the plasma membrane via a peptide-only membrane anchor. J. Virol. 2007, 81, 3187–3197. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boonyaratanakornkit, J.; Schomacker, H.; Collins, P.; Schmidt, A. Alix serves as an adaptor that allows human parainfluenza virus type 1 to interact with the host cell ESCRT system. PLoS ONE 2013, 8, e59462. [Google Scholar] [CrossRef] [Green Version]
- Irie, T.; Shimazu, Y.; Yoshida, T.; Sakaguchi, T. The YLDL sequence within Sendai virus M protein is critical for budding of virus-like particles and interacts with Alix/AIP1 independently of C protein. J. Virol. 2007, 81, 2263–2273. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gosselin-Grenet, A.-S.; Marq, J.-B.; Abrami, L.; Garcin, D.; Roux, L. Sendai virus budding in the course of an infection does not require Alix and VPS4A host factors. Virology 2007, 365, 101–112. [Google Scholar] [CrossRef] [PubMed]
- Salditt, A.; Koethe, S.; Pohl, C.; Harms, H.; Kolesnikova, L.; Becker, S.; Schneider-Schaulies, S. Measles virus M protein-driven particle production does not involve the endosomal sorting complex required for transport (ESCRT) system. J. Gen. Virol. 2010, 91, 1464–1472. [Google Scholar] [CrossRef] [PubMed]
- Singh, B.K.; Hornick, A.L.; Krishnamurthy, S.; Locke, A.C.; Mendoza, C.A.; Mateo, M.; Miller-Hunt, C.L.; Cattaneo, R.; Sinn, P.L. The Nectin-4/Afadin Protein Complex and Intercellular Membrane Pores Contribute to Rapid Spread of Measles Virus in Primary Human Airway Epithelia. J. Virol. 2015, 89, 7089–7096. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Singh, B.K.; Pfaller, C.K.; Cattaneo, R.; Sinn, P.L. Measles Virus Ribonucleoprotein Complexes Rapidly Spread across Well-Differentiated Primary Human Airway Epithelial Cells along F-Actin Rings. mBio 2019, 10. [Google Scholar] [CrossRef] [Green Version]
- Cifuentes-Muñoz, N.; Dutch, R.E.; Cattaneo, R. Direct cell-to-cell transmission of respiratory viruses: The fast lanes. PLoS Pathog. 2018, 14, e1007015. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Contreras, E.M.; Monreal, I.A.; Ruvalcaba, M.; Ortega, V.; Aguilar, H.C. Antivirals targeting paramyxovirus membrane fusion. Curr. Opin. Virol. 2021, 51, 34–47. [Google Scholar] [CrossRef] [PubMed]
- Cox, R.M.; Sourimant, J.; Toots, M.; Yoon, J.-J.; Ikegame, S.; Govindarajan, M.; Watkinson, R.E.; Thibault, P.; Makhsous, N.; Lin, M.J.; et al. Orally efficacious broad-spectrum allosteric inhibitor of paramyxovirus polymerase. Nat. Microbiol. 2020, 5, 1232–1246. [Google Scholar] [CrossRef] [PubMed]
- Plemper, R.K. Measles Resurgence and Drug Development. Curr. Opin. Virol. 2020, 41, 8–17. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Genus | Species | Abbreviation | Genome Sequence | C Protein Sequence | Reference | |
|---|---|---|---|---|---|---|
| (GenBank) | (GenBank) | (UniProt/UniParc) | ||||
| Aquapara-myxovirus | Oncorhynchus aquaparamyxovirus | PSPV | MH900516.1 | AYN62575.1 | I1TLL1 | [29] |
| Salmo aquaparamyxovirus | ASPV | NC_025360.1 | YP_009094145.1 | B2BX73 | [30] | |
| Henipavirus | Cedar henipavirus | CeV | NC_025351.1 | YP_009094083.1 | J7H4I1 | [31] |
| Hendra henipavirus | HeV | NC_001906.3 | NP_047109.1 | O55779 | [32] | |
| Nipah henipavirus | NiV | NC_002728.1 | NP_112024.1 | Q997F1 | [33] | |
| Jeilongvirus | Beilong jeilongvirus | BeV | NC_007803.1 | YP_512248.1 | Q287X7 | [34] |
| Jun jeilongvirus | J-V | NC_007454.1 | YP_338079.1 | Q49HN8 | [35] | |
| Morbillivirus | Canine morbillivirus | CDV | NC_001921.1 | NP_047203.1 | P06941 | [36] |
| Cetacean morbillivirus | CeMV | NC_005283.1 | NP_945026.1 | Q709E7 | [37] | |
| Feline morbillivirus | FeMV | NC_039196.1 | YP_009512960.1 | UPI000259F006 | [38] | |
| Measles morbillivirus | MeV | NC_001498.1 | NP_056920.1 | Q9YZN9 | [39] | |
| Phocine morbillivirus | PMV | NC_028249.1 | YP_009177600.1 | P35940 | [40] | |
| Rinderpest morbillivirus | RPV | NC_006296.2 | YP_087122.1 | P35948 | [41] | |
| Small ruminant morbillivirus | PPRV | NC_006383.2 | YP_133824.1 | Q5ZER5 | [42] | |
| Narmovirus | Mossman narmovirus | MoV | NC_005339.1 | NP_958051.1 | Q6WGM3 | [43] |
| Myodes narmovirus | BaVV | NC_055167.1 | YP_010085011.1 | N/A | [44] | |
| Nariva narmovirus | NarV | NC_017937.1 | YP_006347585.1 | B8XH61 | [45] | |
| Tupaia narmovirus | TPMV | NC_002199.1 | NP_054693.1 | Q9WS38 | [46] | |
| Respirovirus | Bovine respirovirus 3 | BPIV3 | NC_002161.1 | N/A | N/A | [47] |
| Caprine respirovirus 3 | CPIV3 | NC_028362.1 | N/A | N/A | [48] | |
| Human respirovirus 1 | HPIV1 | NC_003461.1 | NP_604436.1 | Q8QT30 | [49] | |
| Human respirovirus 3 | HPIV3 | NC_001796.2 | NP_599251.1 | UPI0000161E9C | [50] | |
| Murine respirovirus | SeV | NC_001552.1 | NP_056872.1 | O55527 | [51] | |
| Porcine respirovirus 1 | PPIV1 | NC_025402.1 | YP_009094446.1 | S5LSI4 | [52] | |
| Squirrel respirovirus | GSqRV | LS992584.1 | SYZ47172.1 | A0A383S9W5 | [53] | |
| Salemvirus | Salem salemvirus | SalV | NC_025386.1 | YP_009094334.1 | Q9IZB9 | [54] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Siering, O.; Cattaneo, R.; Pfaller, C.K. C Proteins: Controllers of Orderly Paramyxovirus Replication and of the Innate Immune Response. Viruses 2022, 14, 137. https://doi.org/10.3390/v14010137
Siering O, Cattaneo R, Pfaller CK. C Proteins: Controllers of Orderly Paramyxovirus Replication and of the Innate Immune Response. Viruses. 2022; 14(1):137. https://doi.org/10.3390/v14010137
Chicago/Turabian StyleSiering, Oliver, Roberto Cattaneo, and Christian K. Pfaller. 2022. "C Proteins: Controllers of Orderly Paramyxovirus Replication and of the Innate Immune Response" Viruses 14, no. 1: 137. https://doi.org/10.3390/v14010137