
miR-541-3p Promoted Porcine Reproductive and Respiratory Syndrome Virus 2 (PRRSV-2) Replication by Targeting Interferon Regulatory Factor 7

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cells and Virus
2.2. Antibodies and Reagents
2.3. Plasmids
2.4. RNA Quantification
2.5. Dual Luciferase Reporter Assay
2.6. RNA Interference Assay
2.7. Virus Titration
2.8. Statistical Analysis
3. Results
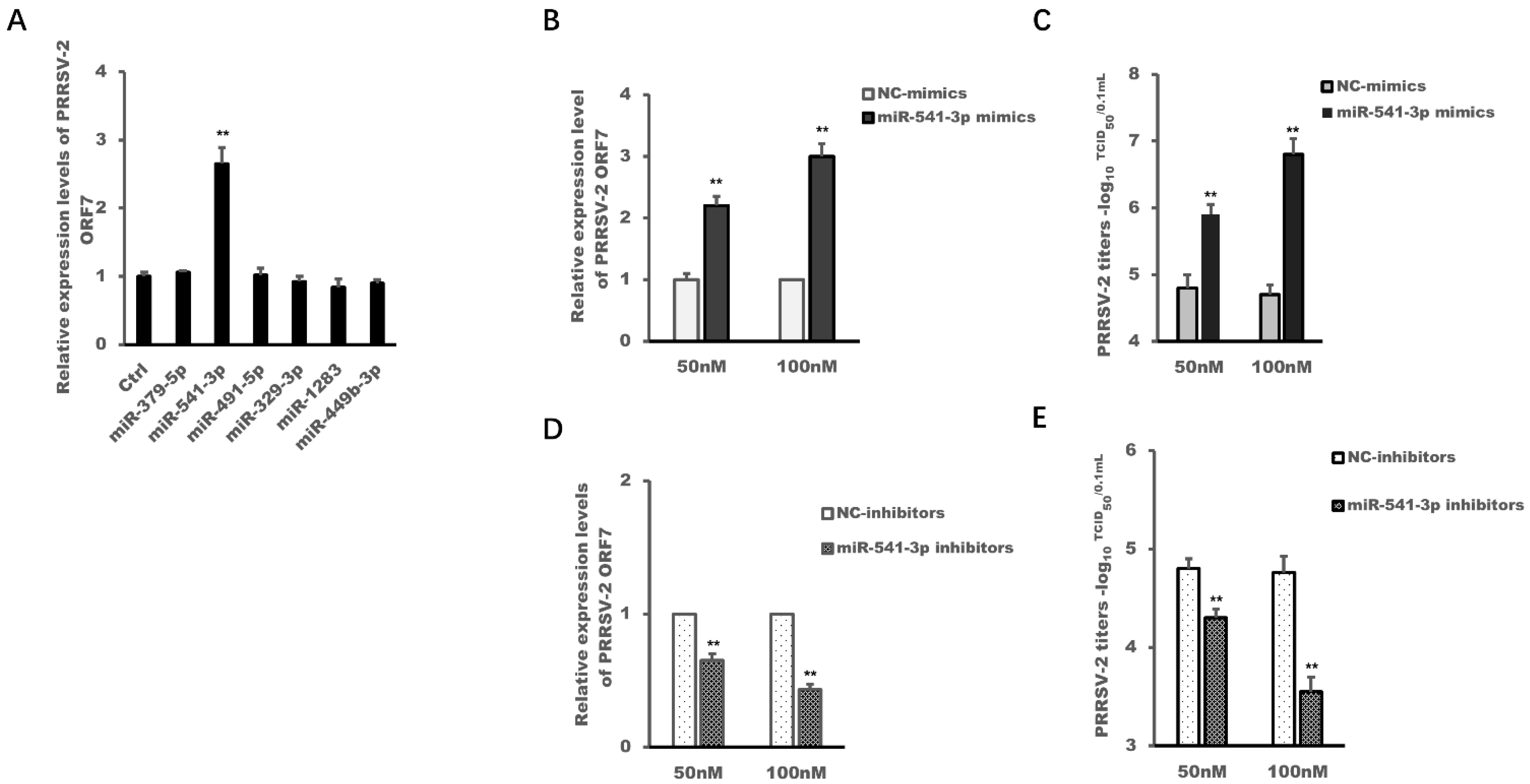
3.1. miR-541-3p Facilitated the Replication of PRRSV-2 in MARC-145 Cells
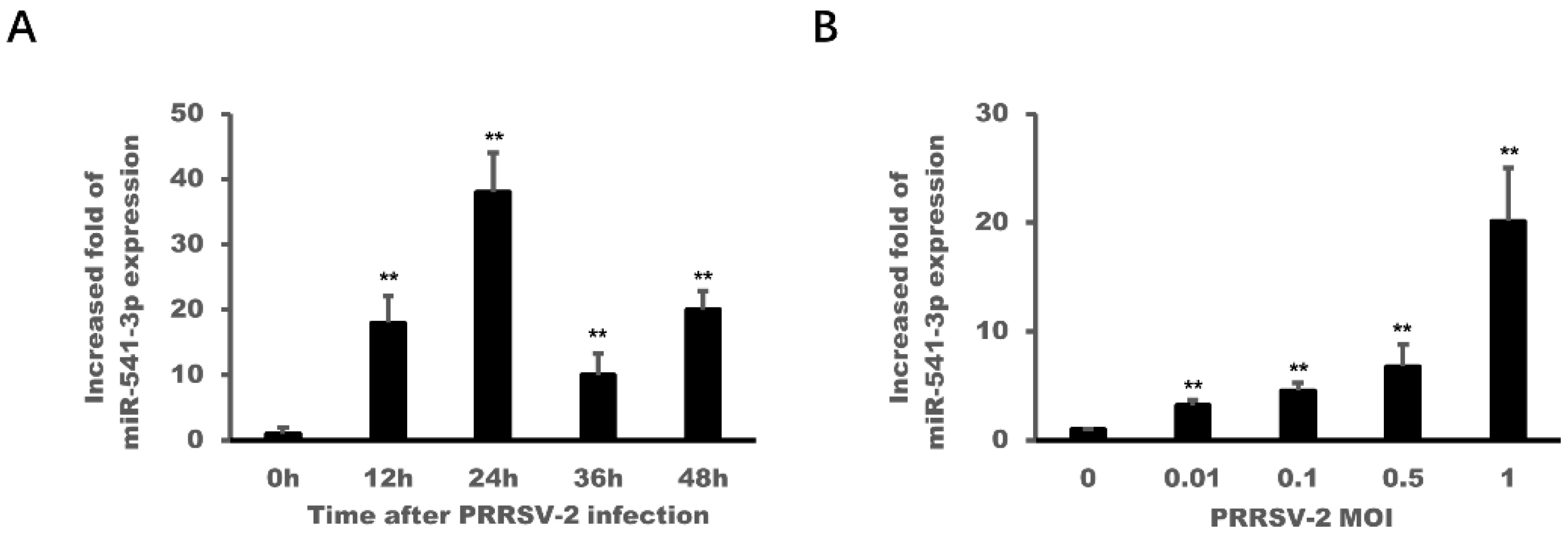
3.2. PRRSV-2 Infection Up-Regulated the Expression of miR-541-3p in MARC-145 Cells
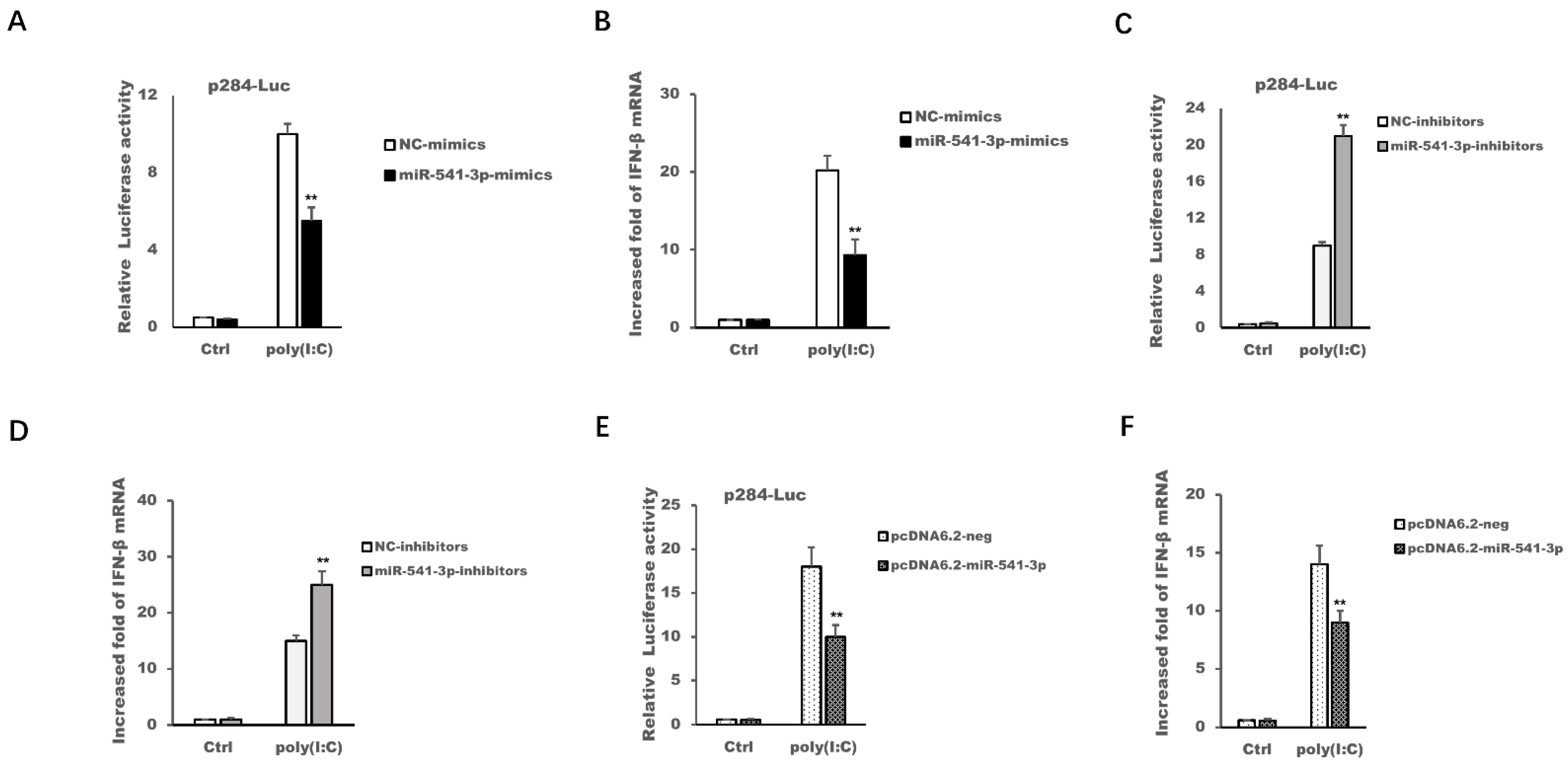
3.3. miR-541-3p Negatively Regulated Poly(I:C)-Induced Transcription of Type I Interferon
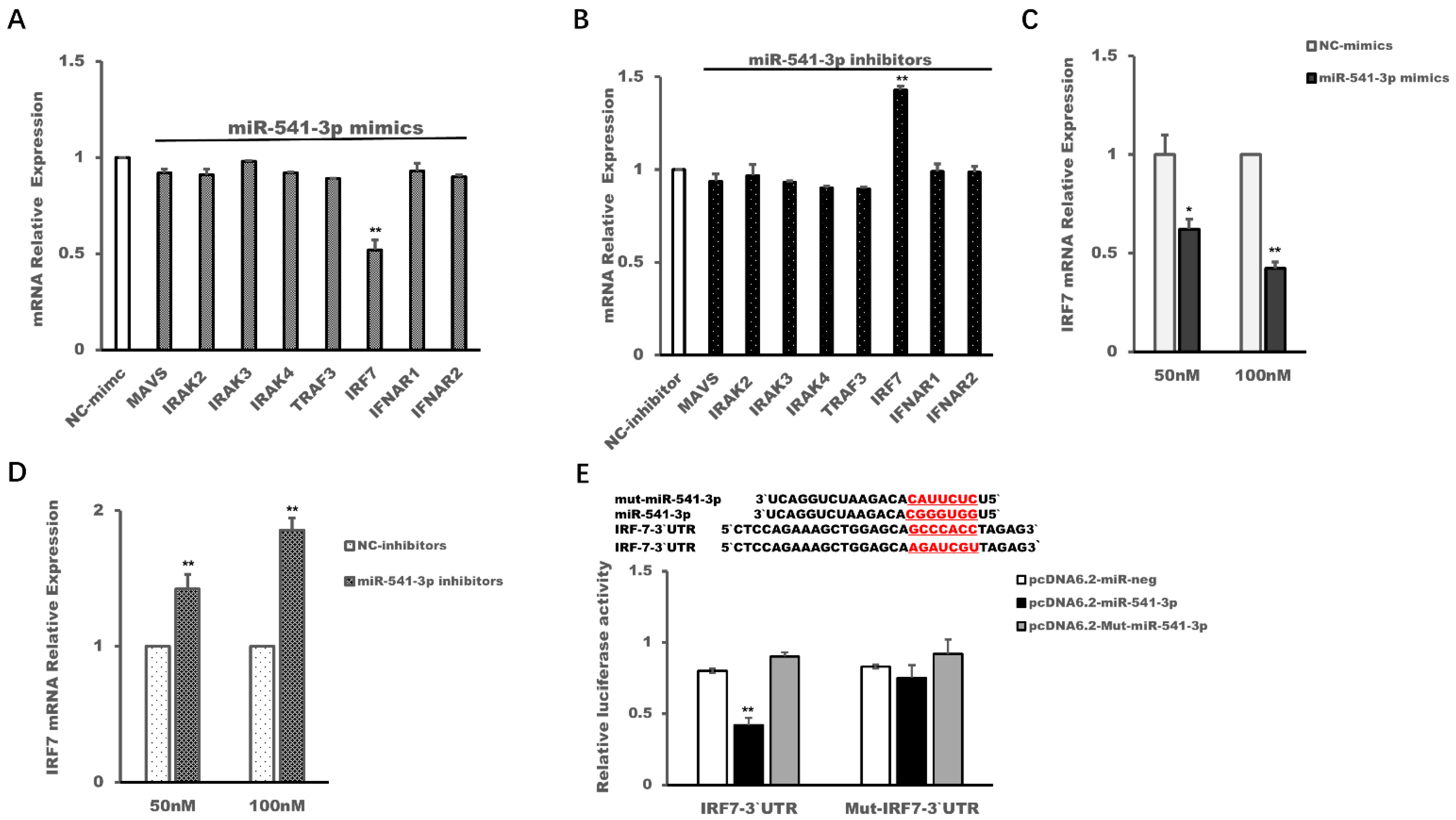
3.4. Interferon Regulatory Factor (IRF) 7 Was a Target Gene of miR-541-3p
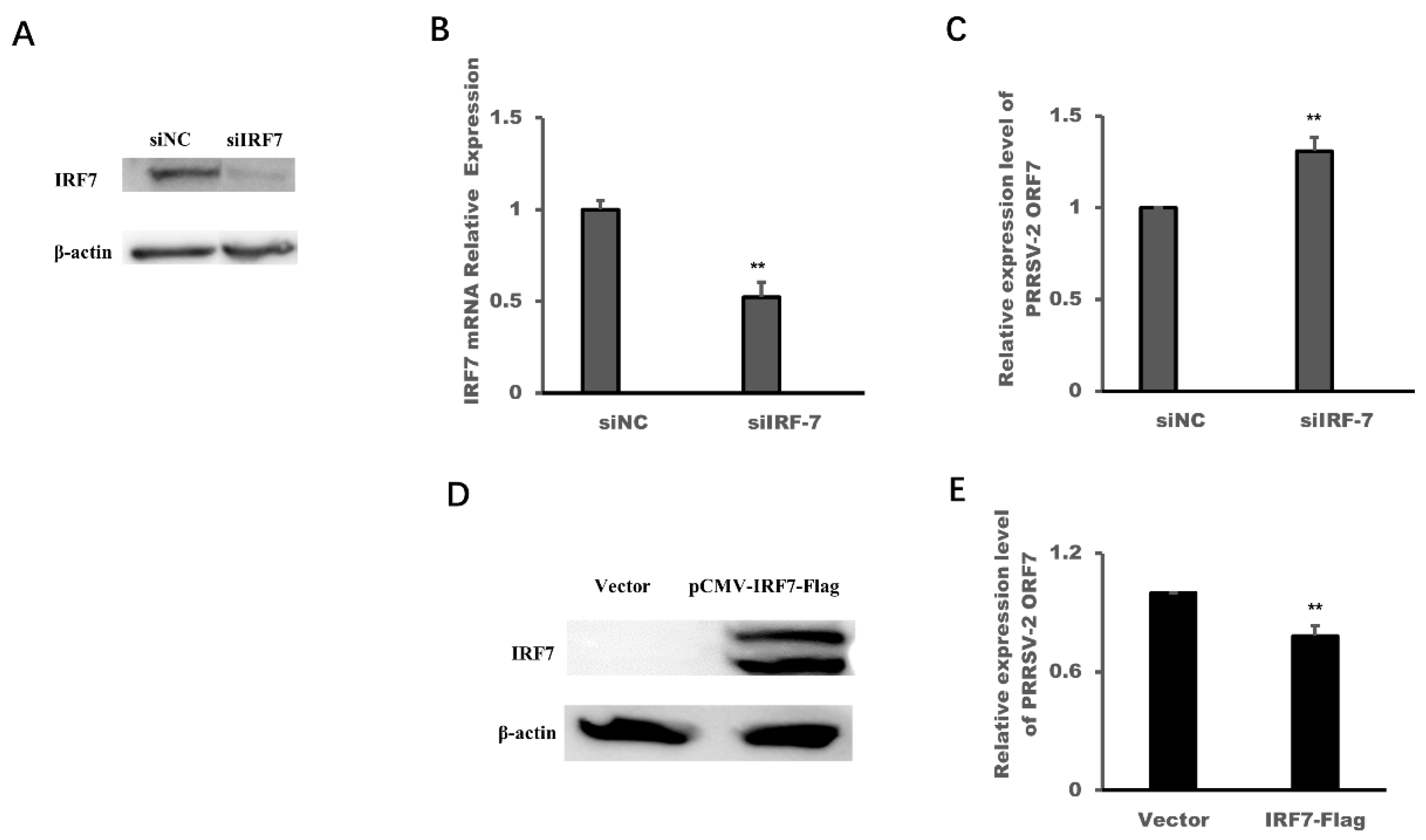
3.5. PRRSV-2 Infection Down-Regulated the Expression of IRF7 and the Over-Expression of IRF7 Could Inhibit the Replication of PRRSV-2 in MARC-145 Cells
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Holtkamp, D.J.; Kliebenstein, J.B.; Neumann, E.J.; Zimmerman, J.J.; Rotto, H.F.; Yoder, T.K.; Wang, C.; Yeske, P.E.; Mowrer, C.L.; Haley, C.A. Assessment of the economic impact of porcine reproductive and respiratory syndrome virus on United States pork producers. J. Swine Health Prod. 2013, 21, 72–84. [Google Scholar]
- Ruedas-Torres, I.; Rodriguez-Gomez, I.M.; Sanchez-Carvajal, J.M.; Larenas-Munoz, F.; Pallares, F.J.; Carrasco, L.; Gomez-Laguna, J. The jigsaw of PRRSV virulence. Vet. Microbiol. 2021, 260, 109168. [Google Scholar] [CrossRef]
- Walker, P.J.; Siddell, S.G.; Lefkowitz, E.J.; Mushegian, A.R.; Adriaenssens, E.M.; Alfenas-Zerbini, P.; Davison, A.J.; Dempsey, D.M.; Dutilh, B.E.; Garcia, M.L.; et al. Changes to virus taxonomy and to the International Code of Virus Classification and Nomenclature ratified by the International Committee on Taxonomy of Viruses (2021). Arch. Virol. 2021, 166, 2633–2648. [Google Scholar] [CrossRef]
- Chrun, T.; Maze, E.A.; Vatzia, E.; Martini, V.; Paudyal, B.; Edmans, M.D.; McNee, A.; Manjegowda, T.; Salguero, F.J.; Wanasen, N.; et al. Simultaneous Infection With Porcine Reproductive and Respiratory Syndrome and Influenza Viruses Abrogates Clinical Protection Induced by Live Attenuated Porcine Reproductive and Respiratory Syndrome Vaccination. Front. Immunol. 2021, 12. [Google Scholar] [CrossRef] [PubMed]
- Montaner-Tarbes, S.; del Portillo, H.A.; Montoya, M.; Fraile, L. Key Gaps in the Knowledge of the Porcine Respiratory Reproductive Syndrome Virus (PRRSV). Front. Vet. Sci. 2019, 6, 38. [Google Scholar] [CrossRef] [Green Version]
- An, T.Q.; Li, J.N.; Su, C.M.; Yoo, D. Molecular and Cellular Mechanisms for PRRSV Pathogenesis and Host Response to Infection. Virus Res. 2020, 286, 197980. [Google Scholar] [CrossRef] [PubMed]
- Grey, F.; Tirabassi, R.; Meyers, H.; Wu, G.; McWeeney, S.; Hook, L.; Nelson, J.A. A viral microRNA down-regulates multiple cell cycle genes through mRNA 5’UTRs. PLoS Pathog. 2010, 6, e1000967. [Google Scholar] [CrossRef] [Green Version]
- Ameres, S.L.; Zamore, P.D. Diversifying microRNA sequence and function. Nat. Rev. Mol. Cell Biol. 2013, 14, 475–488. [Google Scholar] [CrossRef]
- Lin, S.; Gregory, R.I. MicroRNA biogenesis pathways in cancer. Nat. Rev. Cancer 2015, 15, 321–333. [Google Scholar] [CrossRef] [PubMed]
- Forster, S.C.; Tate, M.D.; Hertzog, P.J. MicroRNA as Type I Interferon-Regulated Transcripts and Modulators of the Innate Immune Response. Front. Immunol. 2015, 6, 334. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ma, F.; Xu, S.; Liu, X.; Zhang, Q.; Xu, X.; Liu, M.; Hua, M.; Li, N.; Yao, H.; Cao, X. The microRNA miR-29 controls innate and adaptive immune responses to intracellular bacterial infection by targeting interferon-gamma. Nat. Immunol. 2011, 12, 861–869. [Google Scholar] [CrossRef] [PubMed]
- Zhou, X.; Michal, J.J.; Jiang, Z.H.; Liu, B. MicroRNA expression profiling in alveolar macrophages of indigenous Chinese Tongcheng pigs infected with PRRSV in vivo. J. Appl. Genet. 2017, 58, 539–544. [Google Scholar] [CrossRef]
- Liu, F.; Du, Y.P.; Feng, W.H. New perspective of host microRNAs in the control of PRRSV infection. Vet. Microbiol. 2017, 209, 48–56. [Google Scholar] [CrossRef] [PubMed]
- Zhou, A.; Li, S.F.; Zhang, S.J. miRNAs and genes expression in MARC-145 cell in response to PRRSV infection. Infect. Genet. Evol. 2014, 27, 173–180. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Shi, X.; Zhang, X.; Wang, A.; Wang, L.; Yang, Y.; Deng, R.; Zhang, G.P. MicroRNA 373 Facilitates the Replication of Porcine Reproductive and Respiratory Syndrome Virus by Its Negative Regulation of Type I Interferon Induction. J. Virol. 2017, 91, e01311-16. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Q.; Guo, X.K.; Gao, L.; Huang, C.; Li, N.; Jia, X.; Liu, W.; Feng, W.H. MicroRNA-23 inhibits PRRSV replication by directly targeting PRRSV RNA and possibly by upregulating type I interferons. Virology 2014, 450, 182–195. [Google Scholar] [CrossRef] [Green Version]
- Chang, X.; Shi, X.; Zhang, X.; Chen, J.; Fan, X.; Yang, Y.; Wang, L.; Wang, A.; Deng, R.; Zhou, E.; et al. miR-382-5p promotes porcine reproductive and respiratory syndrome virus (PRRSV) replication by negatively regulating the induction of type I interferon. FASEB J. 2020, 34, 4497–4511. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Q.; Huang, C.; Yang, Q.; Gao, L.; Liu, H.C.; Tang, J.; Feng, W.H. MicroRNA-30c Modulates Type I IFN Responses To Facilitate Porcine Reproductive and Respiratory Syndrome Virus Infection by Targeting JAK1. J. Immunol. 2016, 196, 2272–2282. [Google Scholar] [CrossRef] [Green Version]
- Kim, H.S.; Kwang, J.; Yoon, I.J.; Joo, H.S.; Frey, M.L. Enhanced replication of porcine reproductive and respiratory syndrome (PRRS) virus in a homogeneous subpopulation of MA-104 cell line. Arch. Virol. 1993, 133, 477–483. [Google Scholar] [CrossRef]
- Shi, X.; Wang, L.; Zhi, Y.; Xing, G.; Zhao, D.; Deng, R.; Zhang, G. Porcine reproductive and respiratory syndrome virus (PRRSV) could be sensed by professional beta interferon-producing system and had mechanisms to inhibit this action in MARC-145 cells. Virus Res. 2010, 153, 151–156. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Chang, X.B.; Yang, Y.Q.; Gao, J.C.; Zhao, K.; Guo, J.C.; Ye, C.; Jiang, C.G.; Tian, Z.J.; Cai, X.H.; Tong, G.Z.; et al. Annexin A2 binds to vimentin and contributes to porcine reproductive and respiratory syndrome virus multiplication. Vet. Res. 2018, 49, 75. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reed, L.J.; Muench, H. A simple method of estimating fifty percent endpoints. Am. J. Epidemiol. 1938, 27, 493–497. [Google Scholar] [CrossRef]
- Buddaert, W.; Van Reeth, K.; Pensaert, M. In vivo and in vitro interferon (IFN) studies with the porcine reproductive and respiratory syndrome virus (PRRSV). Adv. Exp. Med. Biol. 1998, 440, 461–467. [Google Scholar]
- Overend, C.; Mitchell, R.; He, D.; Rompato, G.; Grubman, M.J.; Garmendia, A.E. Recombinant swine beta interferon protects swine alveolar macrophages and MARC-145 cells from infection with Porcine reproductive and respiratory syndrome virus. J. Gen. Virol. 2007, 88, 925–931. [Google Scholar] [CrossRef]
- Shi, X.; Zhang, X.; Wang, L.; Li, W.; Jiang, B.; Deng, R.; Wang, A.; Zhang, G. Recombinant beta interferon could clear the low-dose infected porcine reproductive and respiratory syndrome virus (PRRSV) in MARC-145 cells. Acta Virol. 2016, 60, 290–297. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Beura, L.K.; Sarkar, S.N.; Kwon, B.; Subramaniam, S.; Jones, C.; Pattnaik, A.K.; Osorio, F.A. Porcine reproductive and respiratory syndrome virus nonstructural protein 1beta modulates host innate immune response by antagonizing IRF3 activation. J. Virol. 2010, 84, 1574–1584. [Google Scholar] [CrossRef] [Green Version]
- Yang, F.; Xu, Z.L.; Duan, S.B.; Luo, M. MicroRNA-541 promotes the proliferation of vascular smooth muscle cells by targeting IRF7. Am. J. Transl. Res. 2016, 8, 506–515. [Google Scholar]
- Allende, R.; Laegreid, W.W.; Kutish, G.F.; Galeota, J.A.; Wills, R.W.; Osorio, F.A. Porcine reproductive and respiratory syndrome virus: Description of persistence in individual pigs upon experimental infection. J. Virol. 2000, 74, 10834–10837. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, C.; Shi, X.; Zhang, X.; Wang, A.; Wang, L.; Chen, J.; Deng, R.; Zhang, G. The Endoribonuclease Activity Essential for the Nonstructural Protein 11 of Porcine Reproductive and Respiratory Syndrome Virus to Inhibit NLRP3 Inflammasome-Mediated IL-1beta Induction. DNA Cell Biol. 2015, 34, 728–735. [Google Scholar] [CrossRef]
- Chen, J.; Shi, X.; Zhang, X.; Wang, L.; Luo, J.; Xing, G.; Deng, R.; Yang, H.; Li, J.; Wang, A.; et al. Porcine Reproductive and Respiratory Syndrome Virus (PRRSV) Inhibits RNA-Mediated Gene Silencing by Targeting Ago-2. Viruses 2015, 7, 5539–5552. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xia, Y.H.; Ren, L.; Li, J.Z.; Gao, F. Role of miR-541-3p/TMPRSS4 in the metastasis and EMT of hepatocellular carcinoma. Eur. Rev. Med. Pharmacol. Sci. 2019, 23, 10721–10728. [Google Scholar] [PubMed]
- Lu, Y.J.; Liu, R.Y.; Hu, K.; Wang, Y. MiR-541-3p reverses cancer progression by directly targeting TGIF2 in non-small cell lung cancer. Tumor Biol. 2016, 37, 12685–12695. [Google Scholar] [CrossRef] [PubMed]
- He, Z.H.; Shen, F.H.; Qi, P.; Zhai, Z.X.; Wang, Z.P. miR-541-3p enhances the radiosensitivity of prostate cancer cells by inhibiting HSP27 expression and downregulating beta-catenin. Cell Death Discov. 2021, 7, 18. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primers | Sequence (5′-3′) |
|---|---|
| MAVS-For | CTGCCTCACAGCAAGAGACCA |
| MAVS-Rev | GTAGACACAGGCCACTTCGTC |
| IRAK2-For | AGGCACGGGAAGCCATTCGT |
| IRAK2-Rev | AGCCCAGCACAGGTAAGACA |
| IRAK3-For | TTGGTCCTGGGCACAGAAA |
| IRAK3-Rev | AATAGCTCGACGATGCCCCA |
| IRAK4-For | TTGCTCAGGGTGCAGCTAAT |
| IRAK4-Rev | GTGCAAGGCCAAAGTCAGAT |
| TRAF3-For | CTCACAAATGCAGCGTCCAG |
| TRAF3-Rev | GCTCCACTCCTTCAGCAGGT |
| IRF7-For | GCAACGTGAGGGTGTGTCTT |
| IRF7-Rev | GCTCCATGAGCAAGCACTCAA |
| IFNAR1-For | TCGTTTACACCATTTCGCAAAGCTCAAA |
| IFNAR1-Rev | ACGATCCAAAGCCCACATGACACTATCT |
| IFNAR2-For | TGTCATTGAAGAGCAGTCAGAGGGGATT |
| IFNAR2-Rev | GTTGAAGGAGGGTGCATTTTAAGGGAGA |
| GAPDH-For | TGACAACAGCCTCAAGATCG |
| GAPDH-Rev | GTCTTCTGGGTGGCAGTGAT |
| ORF7-For | AAACCAGTCCAGAGGCAAGG |
| ORF7-Rev | GCAAACTAAACTCCACAGTGTAA |
| IFN-β-For | CTAGCACTGGCTGGAATGAGACT |
| IFN-β-Rev | GGCCTTCAGGTAATGCAGAATC |
| Flag-IRF7-For | CCCAAGCTTATGGCCTTGGCTCCTGAGAG |
| Flag-IRF7-Ror | CGCGGATCCCTAGACGGGCTGCTCCAGCTC |
| Top strand oligo for miR-541-3p | GGATCCTGGAGGCTTGCTGAAGGCTGTATGCTGTGGTGGGCACAGAATCTGGACTGTTTTGGCCACTGACTGAC |
| Bottom strand oligo for miR-541-3p | CAAAACCGGTGACTGACTGTCAGGTCTGACACGGGTGGTGTCCTGTGTTCCGGACAATGATCGTGAGTGTACCTTGTTTACCGGGTCTAGA |
| pcDNA6.2-miR-541-3p-For | GGATCCTGGAGGCTTGCTGAA |
| pcDNA6.2-miR-541-3p-Rev | AGATCTGGGCCATTTGTTCCATGT |
| Top strand oligo for mut-miR-541-3p | GGATCCTGGAGGCTTGCTGAAGGCTGTATGCTGTCTCTTACACAGAATCTGGACTGTTTTGGCCACTGACTGAC |
| Bottom strand oligo for mut-miR-541-3p | CAAAACCGGTGACTGACTGTCAGGTCTGACACATTCTCTGTCCTGTGTTCCGGACAATGATCGTGAGTGTACCTTGTTTACCGGGTCTAGA |
| IRF7-3′UTR For | TCGAACTCCAGAAAGCTGGAGCAGCCCACCTAGAGCTGGCCGC |
| IRF7-3′UTR Rev | GGCCGCGGCCAGCTCTAGGTGGGCTGCTCCAGCTTTCTGGAGT |
| Mut-IRF7-3′UTR | TCGAACTCCAGAAAGCTGGAGCAAGATCGTTAGAGCTGGCCGC |
| Mut-IRF7-3′UTR | GGCCGCGGCCAGCTCTAACGATCTTGCTCCAGCTTTCTGGAGT |
| siIRF7-sense | CCAUCUUUGACUUCAGAGUTT |
| siIRF7-antisense | ACUCUGAAGUCAAAGAUGGTT-3 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shi, X.; Yang, Y.; Zhang, X.; Chang, X.; Chen, J.; Wang, C.; Wang, A.; Wang, J.; Qin, J.; Ye, X.; et al. miR-541-3p Promoted Porcine Reproductive and Respiratory Syndrome Virus 2 (PRRSV-2) Replication by Targeting Interferon Regulatory Factor 7. Viruses 2022, 14, 126. https://doi.org/10.3390/v14010126
Shi X, Yang Y, Zhang X, Chang X, Chen J, Wang C, Wang A, Wang J, Qin J, Ye X, et al. miR-541-3p Promoted Porcine Reproductive and Respiratory Syndrome Virus 2 (PRRSV-2) Replication by Targeting Interferon Regulatory Factor 7. Viruses. 2022; 14(1):126. https://doi.org/10.3390/v14010126
Chicago/Turabian StyleShi, Xibao, Yuanhao Yang, Xiaozhuan Zhang, Xiaobo Chang, Jing Chen, Chao Wang, Aiping Wang, Jianhua Wang, Jianru Qin, Xianlong Ye, and et al. 2022. "miR-541-3p Promoted Porcine Reproductive and Respiratory Syndrome Virus 2 (PRRSV-2) Replication by Targeting Interferon Regulatory Factor 7" Viruses 14, no. 1: 126. https://doi.org/10.3390/v14010126