
Construction, Identification and Analysis of the Interaction Network of African Swine Fever Virus MGF360-9L with Host Proteins

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cells and Virus
2.2. Antibodies and Reagents
2.3. Plasmid Construction and Cell Transfection
2.4. LC-MS/MS
2.5. LC-MS/MS Data Analysis
2.6. Construction and Analysis of PPI Network
2.7. Protein Functional Enrichment Analysis
2.8. Co-Immunoprecipitation
2.9. SDS-PAGE and Immunoblotting Analyses
2.10. Indirect Immunofluorescence Assay (IFA)
2.11. Real-Time qPCR
2.12. Viral Titration (50% Hemadsorption Doses)
2.13. Biosafety Statement and Facility
2.14. Statistical Analysis
3. Results
3.1. Identification of MGF360-9L-Interacting Factors in PK-15 Cells by Liquid Chromatography–Mass Spectrometry
3.2. ASFV MGF360-9L-Host Interactome
3.3. GO Analysis
3.4. KEGG Pathway Enrichment Analysis
3.5. Domain Enrichment Analysis of ASFV MGF360-9L-Interacting Partners
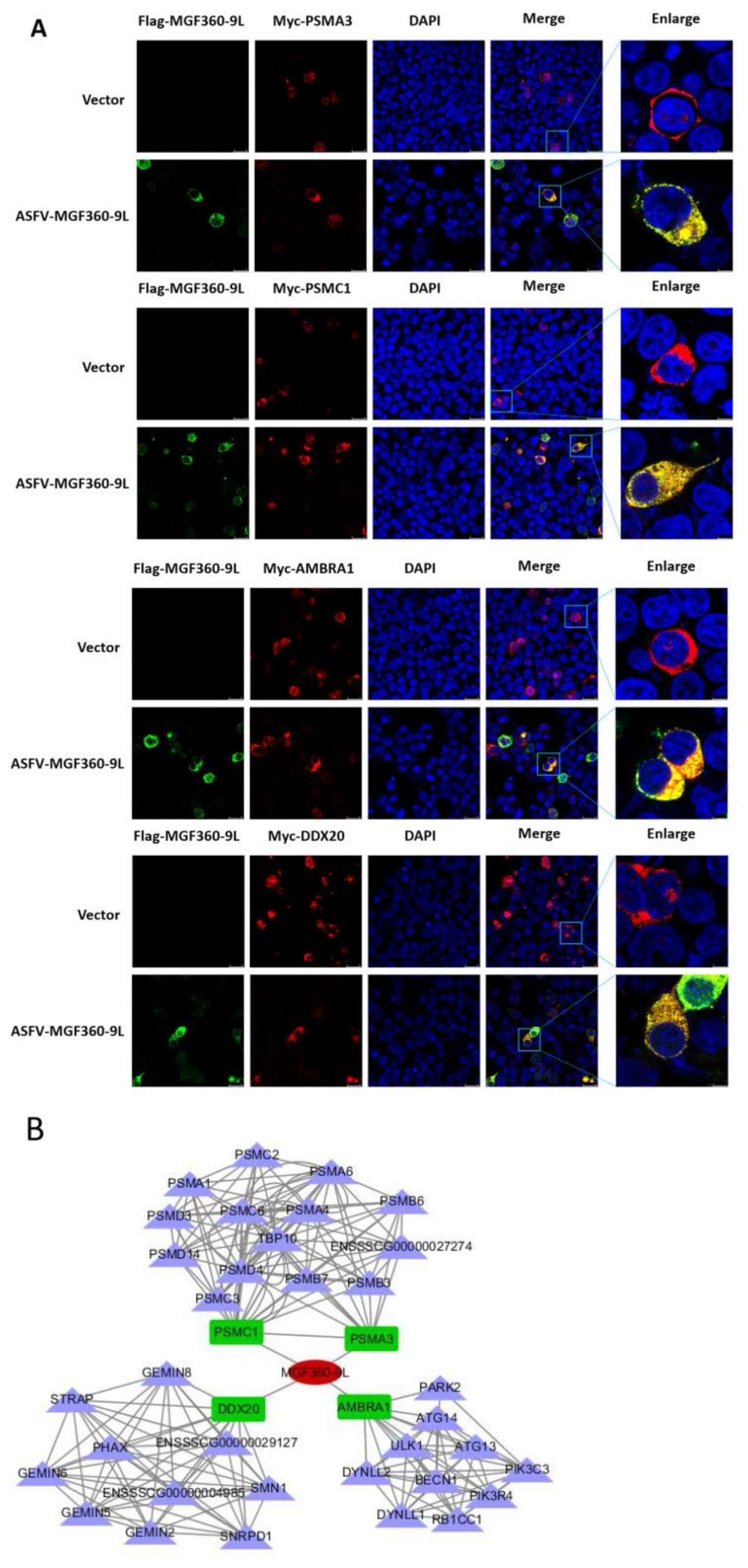
3.6. Validating the Interactions between Cellular Proteins with MGF360-9L Protein, and MGF360-9L May Be Involved in Regulation of Host Metabolism by ASFV
3.7. PSMA3 and PSMC1 Overexpression Significantly Promoted ASFV Replication, and MGF360-9L Maintained the Transcriptional Level of PSMA3 and PSMC1
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Penrith, M.; Bastos, A.; Etter, E.; Beltrán-Alcrudo, D. Epidemiology of African swine fever in Africa today: Sylvatic cycle versus socio-economic imperatives. Transbound. Emerg. Dis. 2019, 66, 672–686. [Google Scholar] [CrossRef]
- Alonso, C.; Borca, M.; Dixon, L.; Revilla, Y.; Rodriguez, F.; Escribano, J.M.; Consortium, I.R. ICTV Virus Taxonomy Profile: Asfarviridae. J. Gen. Virol. 2018, 99, 613–614. [Google Scholar] [CrossRef] [PubMed]
- Zhou, X.; Li, N.; Luo, Y.; Liu, Y.; Miao, F.; Chen, T.; Zhang, S.; Cao, P.; Li, X.; Tian, K.; et al. Emergence of African Swine Fever in China, 2018. Transbound. Emerg. Dis. 2018, 65, 1482–1484. [Google Scholar] [CrossRef] [Green Version]
- Penrith, M.-L. History of ’swine fever’ in southern Africa. J. S. Afr. Vet. Assoc. 2013, 84, 1–6. [Google Scholar] [CrossRef] [Green Version]
- Costard, S.; Wieland, B.; de Glanville, W.; Jori, F.; Rowlands, R.; Vosloo, W.; Roger, F.; Pfeiffer, D.; Dixon, L. African swine fever: How can global spread be prevented? Philos. Trans. R. Soc. London. Ser. B Biol. Sci. 2009, 364, 2683–2696. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Karger, A.; Pérez-Núñez, D.; Urquiza, J.; Hinojar, P.; Alonso, C.; Freitas, F.; Revilla, Y.; Le Potier, M.; Montoya, M. An Update on African Swine Fever Virology. Viruses 2019, 11, 864. [Google Scholar] [CrossRef] [Green Version]
- Dixon, L.K.; Chapman, D.A.G.; Netherton, C.L.; Upton, C. African swine fever virus replication and genomics. Virus Res. 2013, 173, 3–14. [Google Scholar] [CrossRef]
- Chapman, D.; Darby, A.; Da Silva, M.; Upton, C.; Radford, A.; Dixon, L. Genomic analysis of highly virulent Georgia 2007/1 isolate of African swine fever virus. Emerg. Infect. Dis. 2011, 17, 599–605. [Google Scholar] [CrossRef] [PubMed]
- Galindo, I.; Alonso, C. African Swine Fever Virus: A Review. Viruses 2017, 9, 103. [Google Scholar] [CrossRef] [Green Version]
- Barber, C.; Netherton, C.; Goatley, L.; Moon, A.; Goodbourn, S.; Dixon, L. Identification of residues within the African swine fever virus DP71L protein required for dephosphorylation of translation initiation factor eIF2 alpha and inhibiting activation of pro-apoptotic CHOP. Virology 2017, 504, 107–113. [Google Scholar] [CrossRef] [PubMed]
- Shen, Z.; Chen, C.; Yang, Y.; Xie, Z.; Ao, Q.; Lv, L.; Zhang, S.; Chen, H.; Hu, R.; Chen, H.; et al. A novel function of African Swine Fever Virus pE66L in inhibition of host translation by the PKR/eIF2α pathway. J. Virol. 2020, e01872-20. [Google Scholar] [CrossRef]
- Hernaez, B.; Cabezas, M.; Munoz-Moreno, R.; Galindo, I.; Cuesta-Geijo, M.A.; Alonso, C. A179L, a New Viral Bcl2 Homolog Targeting Beclin 1 Autophagy Related Protein. Curr. Mol. Med. 2013, 13, 305–316. [Google Scholar] [CrossRef] [PubMed]
- Nogal, M.L.; de Buitrago, G.G.; Rodriguez, C.; Cubelos, B.; Carrascosa, A.L.; Salas, M.L.; Revilla, Y. African swine fever virus IAP homologue inhibits caspase activation and promotes cell survival in mammalian cells. J. Virol. 2001, 75, 2535–2543. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hurtado, C.; Granja, A.; Bustos, M.; Nogal, M.; González de Buitrago, G.; de Yébenes, V.; Salas, M.; Revilla, Y.; Carrascosa, A. The C-type lectin homologue gene (EP153R) of African swine fever virus inhibits apoptosis both in virus infection and in heterologous expression. Virology 2004, 326, 160–170. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zsak, L.; Lu, Z.; Burrage, T.G.; Neilan, J.G.; Kutish, G.F.; Moore, D.M.; Rock, D.L. African swine fever virus multigene family 360 and 530 genes are novel macrophage host range determinants. J. Virol. 2001, 75, 3066–3076. [Google Scholar] [CrossRef] [Green Version]
- Henriques, E.; Brito, R.; Soares, H.; Ventura, S.; de Oliveira, V.; Parkhouse, R. Modeling of the Toll-like receptor 3 and a putative Toll-like receptor 3 antagonist encoded by the African swine fever virus. Protein Sci. A Publ. Protein Soc. 2011, 20, 247–255. [Google Scholar] [CrossRef] [Green Version]
- Neilan, J.; Zsak, L.; Lu, Z.; Kutish, G.; Afonso, C.; Rock, D. Novel swine virulence determinant in the left variable region of the African swine fever virus genome. J. Virol. 2002, 76, 3095–3104. [Google Scholar] [CrossRef] [Green Version]
- Gallardo, C.; Sánchez, E.; Pérez-Núñez, D.; Nogal, M.; de León, P.; Carrascosa, Á.; Nieto, R.; Soler, A.; Arias, M.; Revilla, Y. African swine fever virus (ASFV) protection mediated by NH/P68 and NH/P68 recombinant live-attenuated viruses. Vaccine 2018, 36, 2694–2704. [Google Scholar] [CrossRef] [PubMed]
- Zhuo, Y.; Guo, Z.; Ba, T.; Zhang, C.; He, L.; Zeng, C.; Dai, H. African Swine Fever Virus MGF360-12L Inhibits Type I Interferon Production by Blocking the Interaction of Importin α and NF-κB Signaling Pathway. Virol. Sin. 2021, 36, 176–186. [Google Scholar] [CrossRef] [PubMed]
- O’Donnell, V.; Holinka, L.; Krug, P.; Gladue, D.; Carlson, J.; Sanford, B.; Alfano, M.; Kramer, E.; Lu, Z.; Arzt, J.; et al. African Swine Fever Virus Georgia 2007 with a Deletion of Virulence-Associated Gene 9GL (B119L), when Administered at Low Doses, Leads to Virus Attenuation in Swine and Induces an Effective Protection against Homologous Challenge. J. Virol. 2015, 89, 8556–8566. [Google Scholar] [CrossRef] [Green Version]
- Reis, A.; Goatley, L.; Jabbar, T.; Sanchez-Cordon, P.; Netherton, C.; Chapman, D.; Dixon, L. Deletion of the African Swine Fever Virus Gene DP148R Does Not Reduce Virus Replication in Culture but Reduces Virus Virulence in Pigs and Induces High Levels of Protection against Challenge. J. Virol. 2017, 91, e01428-17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Borca, M.; Ramirez-Medina, E.; Silva, E.; Vuono, E.; Rai, A.; Pruitt, S.; Holinka, L.; Velazquez-Salinas, L.; Zhu, J.; Gladue, D. Development of a Highly Effective African Swine Fever Virus Vaccine by Deletion of the I177L Gene Results in Sterile Immunity against the Current Epidemic Eurasia Strain. J. Virol. 2020, 94, e02017-19. [Google Scholar] [CrossRef]
- O’Donnell, V.; Risatti, G.; Holinka, L.; Krug, P.; Carlson, J.; Velazquez-Salinas, L.; Azzinaro, P.; Gladue, D.; Borca, M. Simultaneous Deletion of the 9GL and UK Genes from the African Swine Fever Virus Georgia 2007 Isolate Offers Increased Safety and Protection against Homologous Challenge. J. Virol. 2017, 91, e01760-16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reis, A.; Abrams, C.; Goatley, L.; Netherton, C.; Chapman, D.; Sanchez-Cordon, P.; Dixon, L. Deletion of African swine fever virus interferon inhibitors from the genome of a virulent isolate reduces virulence in domestic pigs and induces a protective response. Vaccine 2016, 34, 4698–4705. [Google Scholar] [CrossRef] [Green Version]
- O’Donnell, V.; Holinka, L.G.; Gladue, D.P.; Sanford, B.; Krug, P.W.; Lu, X.; Arzt, J.; Reese, B.; Carrillo, C.; Risatti, G.R.; et al. African Swine Fever Virus Georgia Isolate Harboring Deletions of MGF360 and MGF505 Genes Is Attenuated in Swine and Confers Protection against Challenge with Virulent Parental Virus. J. Virol. 2015, 89, 6048–6056. [Google Scholar] [CrossRef] [Green Version]
- Shen, C.; Li, G.; Zhang, D.; Xu, G.; Hou, J.; Li, D.; Dang, W.; Zhang, K.; Zheng, H.; Liu, X. Gene Sequence Analysis, Protein Structure Prediction and Subcellular Localization of MGF 360-9L from African Swine Fever Virus. Acta Vet. Et Zootech. Sin. 2020, 51, 1371–1381. [Google Scholar] [CrossRef]
- Parker, E.; Silverstein, R.; Verma, S.; Mysorekar, I. Viral-Immune Cell Interactions at the Maternal-Fetal Interface in Human Pregnancy. Front. Immunol. 2020, 11, 522047. [Google Scholar] [CrossRef] [PubMed]
- Dixon, L.K.; Islam, M.; Nash, R.; Reis, A.L. African swine fever virus evasion of host defences. Virus Res. 2019, 266, 25–33. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Liu, R.; Li, Q.; Wang, F.; Zhu, B.; Zheng, M.; Cui, H.; Wen, J.; Zhao, G. Host cell interactome of PB1 N40 protein of H5N1 influenza A virus in chicken cells. J. Proteom. 2019, 197, 34–41. [Google Scholar] [CrossRef]
- Zhou, J.; Li, H.; Yu, T.; Li, J.; Dong, W.; Ojha, N.; Jin, Y.; Gu, J.; Zhou, J. Protein Interactions Network of Porcine Circovirus Type 2 Capsid With Host Proteins. Front. Microbiol. 2020, 11, 1129. [Google Scholar] [CrossRef]
- Carrascosa, A.; Santarén, J.; Viñuela, E. Production and titration of African swine fever virus in porcine alveolar macrophages. J. Virol. Methods 1982, 3, 303–310. [Google Scholar] [CrossRef]
- Zheng, H.; Zhang, K.; Shen, C.; Dang, W.; Li, D.; Liu, H.; Cao, W.; Tian, H.; Ru, Y.; Yang, F.; et al. An Attenuated African Classical Swine Fever Virus Strain with Deletion of MGF360-9L Gene and Its Application. Chinese Patent CN202010294850.2, 28 August 2020. [Google Scholar]
- Schmittgen, T.; Livak, K. Analyzing real-time PCR data by the comparative C(T) method. Nat. Protoc. 2008, 3, 1101–1108. [Google Scholar] [CrossRef]
- Reed, L.J.; Muench, H. A Simple Method of Estimating Fifty Per Cent Endpoints. Am. J. Trop. Med. Hyg. 1938, 27, 493–497. [Google Scholar] [CrossRef]
- Ojha, N.; Liu, J.; Yu, T.; Fang, C.; Zhou, J.; Liao, M. Interplay of the ubiquitin proteasome system and the innate immune response is essential for the replication of infectious bronchitis virus. Arch. Virol. 2021, 166, 2173–2185. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Chen, X.; Kong, N.; Jiao, Y.; Sun, D.; Dong, S.; Qin, W.; Zhai, H.; Yu, L.; Zheng, H.; et al. TRIM21 inhibits porcine epidemic diarrhea virus proliferation by proteasomal degradation of the nucleocapsid protein. Arch. Virol. 2021, 166, 1903–1911. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Guo, R.; Kim, S.; Shah, H.; Zhang, S.; Liang, J.; Fang, Y.; Gentili, M.; Leary, C.; Elledge, S.; et al. SARS-CoV-2 hijacks folate and one-carbon metabolism for viral replication. Nat. Commun. 2021, 12, 1676. [Google Scholar] [CrossRef]
- Yutin, N.; Koonin, E. Hidden evolutionary complexity of Nucleo-Cytoplasmic Large DNA viruses of eukaryotes. Virol. J. 2012, 9, 161. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Golding, J.; Goatley, L.; Goodbourn, S.; Dixon, L.; Taylor, G.; Netherton, C. Sensitivity of African swine fever virus to type I interferon is linked to genes within multigene families 360 and 505. Virology 2016, 493, 154–161. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lloyd, R. Translational control by viral proteinases. Virus Res. 2006, 119, 76–88. [Google Scholar] [CrossRef]
- Gao, Q.; Yang, C.; Ren, C.; Zhang, S.; Gao, X.; Jin, M.; Chen, H.; Ma, W.; Zhou, H. Eukaryotic Translation Elongation Factor 1 Delta Inhibits the Nuclear Import of the Nucleoprotein and PA-PB1 Heterodimer of Influenza A Virus. J. Virol. 2020, 95, e01391-20. [Google Scholar] [CrossRef]
- Sun, N.; Jiang, L.; Ye, M.; Wang, Y.; Wang, G.; Wan, X.; Zhao, Y.; Wen, X.; Liang, L.; Ma, S.; et al. TRIM35 mediates protection against influenza infection by activating TRAF3 and degrading viral PB2. Protein Cell 2020, 11, 894–914. [Google Scholar] [CrossRef] [PubMed]
- Roth, H.; Magg, V.; Uch, F.; Mutz, P.; Klein, P.; Haneke, K.; Lohmann, V.; Bartenschlager, R.; Fackler, O.; Locker, N.; et al. Flavivirus Infection Uncouples Translation Suppression from Cellular Stress Responses. MBio 2017, 8, e02150-16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Castelló, A.; Quintas, A.; Sánchez, E.; Sabina, P.; Nogal, M.; Carrasco, L.; Revilla, Y. Regulation of host translational machinery by African swine fever virus. PLoS Pathog. 2009, 5, e1000562. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barrado-Gil, L.; Galindo, I.; Martínez-Alonso, D.; Viedma, S.; Alonso, C. The ubiquitin-proteasome system is required for African swine fever replication. PLoS ONE 2017, 12, e0189741. [Google Scholar] [CrossRef] [Green Version]
- Ogura, T.; Wilkinson, A. AAA+ superfamily ATPases: Common structure—Diverse function. Genes Cells Devoted Mol. Cell. Mech. 2001, 6, 575–597. [Google Scholar] [CrossRef]
- Cianfanelli, V.; De Zio, D.; Di Bartolomeo, S.; Nazio, F.; Strappazzon, F.; Cecconi, F. Ambra1 at a glance. J. Cell Sci. 2015, 128, 2003–2008. [Google Scholar] [CrossRef] [Green Version]
- Wang, S.; Ni, H.-M.; Dorko, K.; Kumer, S.C.; Schmitt, T.M.; Nawabi, A.; Komatsu, M.; Huang, H.; Ding, W.-X. Increased hepatic receptor interacting protein kinase 3 expression due to impaired proteasomal functions contributes to alcohol-induced steatosis and liver injury. Oncotarget 2016, 7, 17681–17698. [Google Scholar] [CrossRef]
- Curmi, F.; Cauchi, R.J. The multiple lives of DEAD-box RNA helicase DP103/DDX20/Gemin3. Biochem. Soc. Trans. 2018, 46, 329–341. [Google Scholar] [CrossRef]
- Fedorova, O.A.; Moiseeva, T.N.; Nikiforov, A.A.; Tsimokha, A.S.; Livinskaya, V.A.; Hodson, M.; Bottrill, A.; Evteeva, I.N.; Ermolayeva, J.B.; Kuznetzova, I.M.; et al. Proteomic analysis of the 20S proteasome (PSMA3)-interacting proteins reveals a functional link between the proteasome and mRNA metabolism. Biochem. Biophys. Res. Commun. 2011, 416, 258–265. [Google Scholar] [CrossRef]
- Zhang, J.; Ling, N.; Lei, Y.; Peng, M.; Hu, P.; Chen, M. Multifaceted Interaction Between Hepatitis B Virus Infection and Lipid Metabolism in Hepatocytes: A Potential Target of Antiviral Therapy for Chronic Hepatitis B. Front. Microbiol. 2021, 12, 636897. [Google Scholar] [CrossRef] [PubMed]
- Mullen, P.; Garcia, G.; Purkayastha, A.; Matulionis, N.; Schmid, E.; Momcilovic, M.; Sen, C.; Langerman, J.; Ramaiah, A.; Shackelford, D.; et al. SARS-CoV-2 infection rewires host cell metabolism and is potentially susceptible to mTORC1 inhibition. Nat. Commun. 2021, 12, 1876. [Google Scholar] [CrossRef] [PubMed]
- Rai, A.; Pruitt, S.; Ramirez-Medina, E.; Vuono, E.; Silva, E.; Velazquez-Salinas, L.; Carrillo, C.; Borca, M.; Gladue, D. Identification of a Continuously Stable and Commercially Available Cell Line for the Identification of Infectious African Swine Fever Virus in Clinical Samples. Viruses 2020, 12, 820. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Zhang, M.; Ye, C.; Ruan, K.; Xu, A.; Gao, F.; Tong, G.; Zheng, H. Inhibition of the DNA-Sensing pathway by pseudorabies virus UL24 protein via degradation of interferon regulatory factor 7. Vet. Microbiol. 2021, 255, 109023. [Google Scholar] [CrossRef] [PubMed]
- Zhang, K.; Yan, M.; Hao, J.; Shen, C.; Zhu, Z.; Zhang, D.; Hou, J.; Xu, G.; Li, D.; Zheng, H.; et al. Foot-and-Mouth Disease Virus Structural Protein VP1 Destroys the Stability of TPL2 Trimer by Degradation TPL2 to Evade Host Antiviral Immunity. J. Virol. 2020, 95, e02149-20. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primers | Sequences (5′-3′) |
|---|---|
| Porcine PSMA3-F | GTGGATAAAGGGGACGCCAT |
| Porcine PSMA3-R | AGCTGCTCCAGTCTGTTTCC |
| Porcine PSMC1-F | GCCAAAGCAAGATGGGTCAAA |
| Porcine PSMC1-R | GGCACTGAGTGTGAGGTGTC |
| ASFV P30 (CP204L)-F | CTCCGATGAGGGCTCTTGCT |
| ASFV P30 (CP204L)-R | AGACGGAATCCTCAGCATCTTC |
| Porcine GAPDH-F | ACATGGCCTCCAAGGAGTAAGA |
| Porcine GAPDH-R | GATCGAGTTGGGGCTGTGACT |
| Protein Domain | p Value |
|---|---|
| AAA | 6.23311 ×10−10 |
| Proteasome_A_N | 3.42772 × 10−6 |
| Iso_dh | 3.35825 × 10−5 |
| RRM | 0.000561107 |
| PP2Ac | 0.000877893 |
| small_GTPase | 0.001859956 |
| S4 | 0.002218629 |
| KH | 0.003060462 |
| KOW | 0.00602828 |
| coiled coil region | 0.008893437 |
| ARM | 0.009385491 |
| PHB | 0.011492043 |
| RAN | 0.015154577 |
| OMPdecase | 0.015154577 |
| CRM1_C | 0.015154577 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, B.; Zhang, D.; Shi, X.; Shen, C.; Hao, Y.; Zhang, T.; Yang, J.; Yuan, X.; Chen, X.; Zhao, D.; et al. Construction, Identification and Analysis of the Interaction Network of African Swine Fever Virus MGF360-9L with Host Proteins. Viruses 2021, 13, 1804. https://doi.org/10.3390/v13091804
Yang B, Zhang D, Shi X, Shen C, Hao Y, Zhang T, Yang J, Yuan X, Chen X, Zhao D, et al. Construction, Identification and Analysis of the Interaction Network of African Swine Fever Virus MGF360-9L with Host Proteins. Viruses. 2021; 13(9):1804. https://doi.org/10.3390/v13091804
Chicago/Turabian StyleYang, Bo, Dajun Zhang, Xijuan Shi, Chaochao Shen, Yu Hao, Ting Zhang, Jinke Yang, Xingguo Yuan, Xuehui Chen, Dengshuai Zhao, and et al. 2021. "Construction, Identification and Analysis of the Interaction Network of African Swine Fever Virus MGF360-9L with Host Proteins" Viruses 13, no. 9: 1804. https://doi.org/10.3390/v13091804