
The Bunyavirales: The Plant-Infecting Counterparts



Abstract
:1. Introduction
1.1. Tospoviridae: Genus Orthotospovirus
1.2. Family Phenuiviridae: Genus Tenuivirus
1.3. Fimoviridae: Genus Emaravirus
2. Virion Composition, Genome Organization, and Intracellular Replication Cycle
2.1. Orthotospoviruses
2.2. Tenuiviruses
2.3. Emaraviruses
3. The Role of Non-Structural, Accessory Proteins in Host Adaptation: Inter/Intracellular Movement and Counter-Defense of Antiviral RNAi
3.1. Orthotospoviruses
3.2. Tenuiviruses
3.3. Emaraviruses
4. Transmission
4.1. Orthotospoviruses
4.2. Tenuiviruses
4.3. Emaraviruses
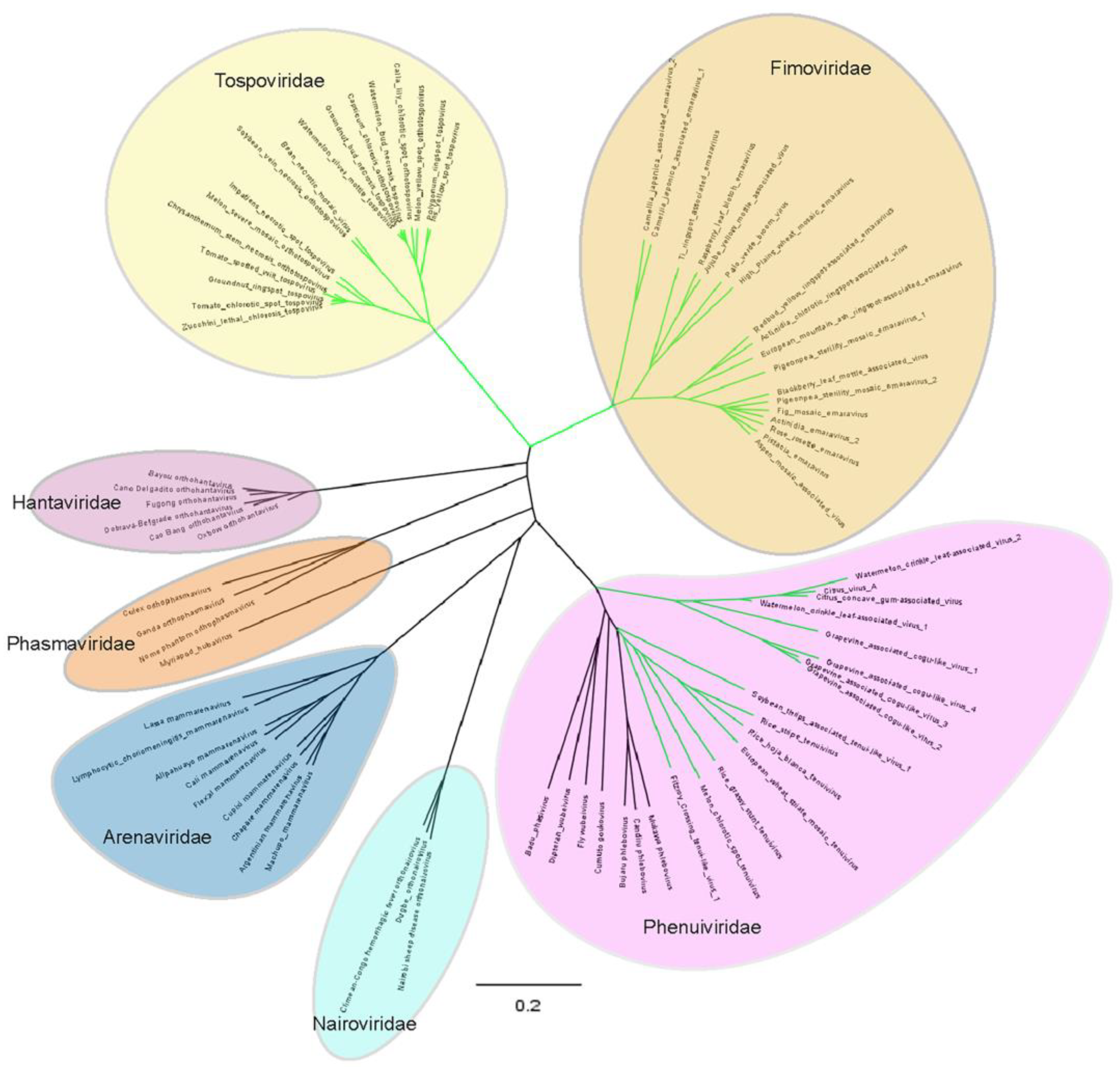
5. Evolution
6. Conclusions and Perspectives
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- Kormelink, R.; Garcia, M.-L.; Goodin, M.; Sasaya, T.; Haenni, A.-L. Negative-Strand RNA Viruses: The plant-infecting counterparts. Virus Res. 2011, 162, 184–202. [Google Scholar] [CrossRef]
- Gorbalenya, A.E.; Siddell, S.G. Recognizing species as a new focus of virus research. PLoS Pathog. 2021, 17, e1009318. [Google Scholar] [CrossRef] [PubMed]
- de Haan, P.; Wagemakers, L.; Peters, D.; Goldbach, R. Molecular cloning and terminal sequence determination of the S and M RNA species of Tomato spotted wilt virus. J. Gen. Virol. 1989, 70, 3469–3474. [Google Scholar] [CrossRef] [PubMed]
- Turina, M.; Kormelink, R.; Resende, R.O. Resistance to Tospovirusus in Vegetable Crops: Epidemiological and Molecular Aspects. Ann. Rev. Phytopathol. 2016, 54, 347–371. [Google Scholar] [CrossRef] [PubMed]
- Oliver, J.E.; Whitfield, A.E. The genus Tospovirus: Emerging bunyaviruses that threaten food security. Ann. Rev. Virol. 2016, 3, 101–124. [Google Scholar] [CrossRef]
- Rotenberg, D.; Jacobson, A.L.; Schneweis, D.J.; Whitfield, A.E. Thrips transmission of tospoviruses. Curr. Opin. Virol. 2015, 15, 80–89. [Google Scholar] [CrossRef]
- Gilbertson, R.L.; Batuman, O.; Webster, C.G.; Adkins, S. Role of the Insect supervectors Bemisia tabaci and Frankliniella occidentalis in the emergence and global spread of plant viruses. Ann. Rev. Virol. 2015, 2, 67–93. [Google Scholar] [CrossRef] [PubMed]
- Scholthof, K.B.; Adkins, S.; Czosnek, H.; Palukaitis, P.; Jacquot, E.; Hohn, T.; Hohn, B.; Saunders, K.; Candresse, T.; Ahlquist, P.; et al. Top 10 plant viruses in molecular plant pathology. Mol. Plant. Pathol. 2011, 12, 938–954. [Google Scholar] [CrossRef] [PubMed]
- de Oliveira, A.S.; Silva Boiteux, L.; Kormelink, R.; Resende, R.O. The Sw-5 gene cluster: Tomato breeding and research towards orthotospovirus disease control. Front. Plant. Sci. 2018, 9, 1055. [Google Scholar] [CrossRef]
- Hedil, M.; Kormelink, R. Viral RNA silencing suppression: The enigma of bunyavirus NSs proteins. Viruses 2016, 8, 208. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kormelink, R. The molecular biology of tospoviruses and resistance strategies. In The Bunyaviridae: Molecular and Cellular Biology; Plyusnin, A., Elliott, R.M., Eds.; Horizon Scientific Press: Norwhich, UK, 2011. [Google Scholar]
- Zhu, M.; van Grinsven, I.L.; Kormelink, R.; Tao, X. Paving the Way to Tospovirus Infection: Multilined Interplays with Plant Innate Immunity. Ann. Rev. Phytopathol. 2019, 57, 2.1–2.22. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gingery, R.E. The rice stripe virus group. In The Plant Viruses; Milne, R.G., Ed.; Plenum Press: New York, NY, USA, 1988; Volume 4, pp. 297–329. [Google Scholar]
- Lecoq, H.; Wipf-Scheibel, C.; Verdin, E.; Desbiez, C. Characterization of the first tenuivirus naturally infecting dicotyledonous plants. Arch. Virol. 2019, 164, 297–301. [Google Scholar] [CrossRef] [PubMed]
- Falk, B.W.; Tsai, J.H. Biology and molecular biology of viruses in the genus Tenuivirus. Ann. Rev. Phytopathol. 1998, 36, 139–163. [Google Scholar] [CrossRef] [PubMed]
- Shiba, T.; Hirae, M.; Hayano-Saito, Y.; Ohto, Y.; Uematsu, H.; Sugiyama, A.; Okuda, M. Spread and yield loss mechanisms of rice stripe disease in rice paddies. Field Crops Res. 2018, 217, 211–217. [Google Scholar] [CrossRef]
- Somera, M.; Kvarnheden, A.; Desbiez, C.; Blystad, D.R.; Soovali, P.; Kundu, J.K.; Gantsovski, M.; Nygren, J.; Lecoq, H.; Verdin, E.; et al. Sixty Years After the First Description: Genome Sequence and Biological Characterization of European Wheat Striate Mosaic Virus Infecting Cereal Crops. Phytopathology 2020, 110, 68–79. [Google Scholar] [CrossRef]
- Mielke, N.; Muehlbach, H.-P. A novel, multipartite, negative-strand RNA virus is associated with the ringspot disease of European mountain ash (Sorbus aucuparia L.). J. Gen. Virol. 2007, 88, 1337–1346. [Google Scholar] [CrossRef] [PubMed]
- Mielke-Ehret, N.; Mühlbach, H.-P. Emaravirus: A novel genus of multipartite, negative strand RNA plant viruses. Viruses 2012, 4, 1515–1536. [Google Scholar] [CrossRef] [Green Version]
- Alemandri, V.; Mattio, M.F.; Rodriguez, S.M.; Truol, G. Geographical distribution and first molecular detection of an Emaravirus, High Plains wheat mosaic virus, in Argentina. Eur. J. Plant. Pathol. 2017. [Google Scholar] [CrossRef]
- Katsiani, A.; Stainton, D.; Lamour, K.; Tzanetakis, I.E. The population structure of Rose rosette virus in the USA. J. Gen. Virol. 2020. [Google Scholar] [CrossRef] [PubMed]
- von Bargen, S.; Dieckmann, H.-L.; Candresse, T.; Mühlbach, H.-P.; Roßbach, J.; Büttner, C. Determination of the complete genome sequence of European mountain ash ringspot-associated emaravirus from Sorbus intermedia reveals two additional genome segments. Arch. Virol. 2019, 164, 1937–1941. [Google Scholar] [CrossRef]
- Herath, V.; Romay, G.; Urrutia, C.D.; Verchot, J. Family Level Phylogenies Reveal Relationships of Plant Viruses within the Order Bunyavirales. Viruses 2020, 12, 1010. [Google Scholar] [CrossRef] [PubMed]
- Gupta, A.K.; Hein, G.L.; Graybosch, R.A.; Tatineni, S. Octapartite negative-sense RNA genome of High Plains wheat mosaic virus encodes two suppressors of RNA silencing. Virology 2018, 518, 152–162. [Google Scholar] [CrossRef]
- Olmedo-Velarde, A.; Park, A.C.; Sugano, J.; Uchida, J.Y.; Kawate, M.; Borth, W.B.; Hu, J.S.; Melzer, M.J. Characterization of Ti Ringspot-Associated Virus, a Novel Emaravirus Associated with an Emerging Ringspot Disease of Cordyline fruticosa. Plant. Dis. 2019, 103, 2345–2352. [Google Scholar] [CrossRef] [PubMed]
- de Haan, P.; Wagemakers, L.; Peters, D.; Goldbach, R. The S RNA segment of Tomato spotted wilt virus has an ambisense character. J. Gen. Virol. 1990, 71, 1001–1008. [Google Scholar] [CrossRef]
- de Haan, P.; Kormelink, R.; Resende, D.; van Poelwijk, F.; Peters, D.; Goldbach, R. Tomato spotted wilt virus L RNA encodes a putative RNA polymerase. J. Gen. Virol. 1991, 72, 2207–2216. [Google Scholar] [CrossRef] [PubMed]
- Kormelink, R.; De Haan, P.; Meurs, C.; Peters, D.; Goldbach, R. The nucleotide sequence of the M RNA segment of Tomato spotted wilt virus: A bunyavirus with two ambisense RNA segments. J. Gen. Virol. 1992, 73, 2795–2804. [Google Scholar] [CrossRef]
- Kormelink, R.; Storms, M.; van Lent, J.; Peters, D.; Goldbach, R. Expression and subcellular location of the NSm protein of Tomato spotted wilt virus (TSWV), a putative viral movement protein. Virology 1994, 200, 56–65. [Google Scholar] [CrossRef]
- Kellman, J.-F.; Liebisch, P.; Schmitz, K.-P.; Piechulla, B. Visual representation by atomif force microscopy (AFM) of tomato spotted wiltvirus ribonucleoproteins. Biol. Chem. 2001, 382, 1559–1562. [Google Scholar] [CrossRef]
- Kormelink, R.; de Haan, P.; Peters, D.; Goldbach, G. Viral RNA synthesis in Tomato spotted wilt virus-infected N. rustica plants. J. Gen. Virol. 1992, 73, 687–693. [Google Scholar] [CrossRef]
- van Knippenberg, I.; Goldbach, R.; Kormelink, R. Tomato spotted wilt virus S-segment mRNAs have overlapping 3’-ends containing a predicted stem-loop structure and conserved sequence motif. Virus Res. 2005, 110, 125–131. [Google Scholar] [CrossRef]
- Kormelink, R.; Van Poelwijk, F.; Peters, D.; Goldbach, R. Non-viral heterogeneous sequences at the 5’ ends of tomato spotted wilt virus (TSWV) mRNAs. J. Gen. Virol. 1992, 73, 2125–2128. [Google Scholar] [CrossRef] [PubMed]
- Geerts-Dimitriadou, C.; Lu, Y.-Y.; Geertsema, C.; Goldbach, R.; Kormelink, R. Analysis of the Tomato spotted wilt virus ambisense S RNA-encoded hairpin structure in translation. PLoS ONE 2012, 7, e31013. [Google Scholar] [CrossRef] [PubMed]
- Guo, Y.; Liu, B.; Ding, Z.; Li, G.; Liu, M.; Zhu, D.; Sun, Y.; Dong, S.; Lou, Z. Distinct Mechanism for the Formation of the Ribonucleoprotein Complex of Tomato Spotted Wilt Virus. J. Virol. 2017, 91, e00892-17. [Google Scholar] [CrossRef] [Green Version]
- Komoda, K.; Narita, M.; Yamashita, K.; Tanaka, I.; Yao, M. Asymmetric Trimeric Ring Structure of the Nucleocapsid Protein of Tospovirus. J. Virol. 2017, 91, e01002-17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, J.; Feng, Z.K.; Wu, J.Y.; Huang, Y.; Lu, G.; Zhu, M.; Wang, B.; Mao, X.; Tao, X. Structure and function analysis of nucleocapsid protein of Tomato spotted wilt virus interacting with RNA using homology modeling. J. Biol. Chem. 2015, 290, 3950–3961. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lima, R.N.; Faheem, M.; Barbosa, J.A.; Polêto, M.D.; Verli, H.; Melo, F.L.; Resende, R.O. Homology modeling and molecular dynamics provide structural insights into tospovirus nucleoprotein. BMC Bioinform. 2016, 17 (Suppl. 18), 489. [Google Scholar] [CrossRef] [Green Version]
- Wichgers Schreur, P.; Kormelink, R.; Kortekaas, J. Genome packaging of the Bunyavirales. Curr. Opin. Virol. 2018, 33, 151–155. [Google Scholar] [CrossRef]
- Reguera, J.; Weber, F.; Cusack, S. Bunyaviridae RNA polymerases (L protein) have an N-terminal, Influenza-like endonuclease domain, essential for viral cap-dependent transcription. PLoS Path. 2010, 6, e1001101. [Google Scholar] [CrossRef] [PubMed]
- Zhao, S.L.; Xu, G.D.; He, G.H.; Peng, Y.Q.; Liang, C.Y. Characterization of an endonuclease in rice stripe tenuivirus Pc1 in vitro. Virus Res. 2019, 260, 33–37. [Google Scholar] [CrossRef]
- Arragain, B.; Effantin, G.; Gerlach, P.; Reguera, J.; Schoehn, G.; Cusack, S.; Malet, H. Pre-initiation and elongation structures of full-length La Crosse virus polymerase reveal functionally important conformational changes. Nat. Commun. 2020, 11, 3590. [Google Scholar] [CrossRef]
- Gogrefe, N.; Reindl, S.; Günther, S.; Rosenthal, M. Structure of a functional cap-binding domain in Rift Valley fever virus L protein. PLoS Pathog. 2019, 15, e1007829. [Google Scholar] [CrossRef] [Green Version]
- Rosenthal, M.; Gogrefe, N.; Vogel, D.; Reguera, J.; Rauschenberger, B.; Cusack, S.; Günther, S.; Reindl, S. Structural insights into reptarenavirus cap-snatching machinery. PLoS Pathog. 2017, 13, e1006400. [Google Scholar] [CrossRef] [Green Version]
- Reguera, J.; Gerlach, P.; Cusack, S. Towards a structural understanding of RNA synthesis by negative strand RNA viral polymerases. Curr. Opin. Struct. Biol. 2016, 36, 75–84. [Google Scholar] [CrossRef] [Green Version]
- Reguera, J.; Gerlach, P.; Rosenthal, M.; Gaudon, S.; Coscia, F.; Günther, S.; Cusack, S. Comparative Structural and Functional Analysis of Bunyavirus and Arenavirus Cap-Snatching Endonucleases. PLoS Pathog. 2016, 12, e1005636. [Google Scholar] [CrossRef]
- Chan, A.Y.; Vreede, F.T.; Smith, M.; Engelhardt, O.G.; Fodor, E. Influenza virus inhibits RNA polymerase II elongation. Virology 2006, 351, 210–217. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Engelhardt, O.G.; Smith, M.; Fodor, E. Association of the influenza A virus RNA-dependent RNA polymerase with cellular RNA polymerase II. J. Virol. 2005, 79, 5812–5818. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ma, X.; Zhou, Y.; Moffett, P. Alterations in cellular RNA decapping dynamics affect tomato spotted wilt virus cap snatching and infection in Arabidopsis. New Phytol. 2019, 224, 789–803. [Google Scholar] [CrossRef]
- Mir, M.A.; Duran, W.A.; Hjelle, B.; Ye, C.; Panganiban, A.T. Storage of cellular 5’ mRNA caps in P bodies for viral cap-snatching. Proc. Natl. Acad. Sci. USA 2008, 105, 19294–19299. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cheng, E.; Mir, M.A. Signatures of host mRNA 5’ terminus for efficient hantavirus cap snatching. J. Virol. 2012, 86, 10173–10185. [Google Scholar] [CrossRef] [Green Version]
- Kitajima, E.W.; de Avila, A.C.; Resende, R.D.O.; Goldbach, R.W.; Peters, D. Comparitive cytological and immunogold labeling studies on different isolates of tomato spotted wilt virus. J. Submicrosc. Cytol. Pathol. 1992, 24, 1–14. [Google Scholar]
- Ribeiro, D.; Borst, J.-W.; Goldbach, R.; Kormelink, R. Tomato spotted wilt virus nucleocapsid protein interacts with both viral glycoproteins Gn and Gc in planta. Virology 2009, 383, 121–130. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ribeiro, D.; Goldbach, R.; Kormelink, R. Trafficking of Tomato spotted wilt virus glycoproteins: Requirements for ER-arrest and exit to the Golgi. Traffic 2009, 10, 664–672. [Google Scholar] [CrossRef] [PubMed]
- Ribeiro, D.; Jung, M.; Moling, S.; Borst, J.-W.; Goldbach, R.; Kormelink, R. The cytosolic nucleoprotein of the plant-infecting bunyavirus Tomato spotted wilt recruits ER-resident proteins to ER export sites. Plant. Cell 2013, 25, 3602–3614. [Google Scholar] [CrossRef] [Green Version]
- Snippe, M.; Smeenk, L.; Goldbach, R.; Kormelink, R. The cytoplasmic domain of tomato spotted wilt virus Gn glycoprotein is required for Golgi localization and interaction with Gc. Virology 2007, 363, 272–279. [Google Scholar] [CrossRef] [Green Version]
- Snippe, M.; Willem Borst, J.; Goldbach, R.; Kormelink, R. Tomato spotted wilt virus Gc and N proteins interact in vivo. Virology 2007, 357, 115–123. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Widana Gamage, S.M.K.; Dietzgen, R.G. Intracellular Localization, Interactions and Functions of Capsicum Chlorosis Virus Proteins. Front. Microbiol. 2017, 8, 612. [Google Scholar] [CrossRef] [Green Version]
- Dietzgen, R.G.; Martin, K.M.; Anderson, G.; Goodin, M.M. In planta localization and interactions of impatiens necrotic spot tospovirus proteins. J. Gen. Virol. 2012, 93, 2490–2495. [Google Scholar] [CrossRef] [Green Version]
- Feng, Z.; Chen, X.; Bao, Y.; Dong, J.; Zhang, Z.; Tao, X. Nucleocapsid of Tomato spotted wilt tospovirus forms mobile particles that traffic on an actin/endoplasmic reticulum network driven by myosin XI-K. New Phytol. 2013, 200, 1212–1224. [Google Scholar] [CrossRef]
- Kikkert, M.; Van Lent, J.; Storms, M.; Bodegom, P.; Kormelink, R.; Goldbach, R. Tomato spotted wilt virus particle morphogenesis in plant cells. J. Virol. 1999, 73, 2288–2297. [Google Scholar] [CrossRef] [Green Version]
- Kikkert, M.; Verschoor, A.; Kormelink, R.; Rottier, P.; Goldbach, R. Tomato spotted wilt virus glycoproteins exhibit trafficking and localization signals that are functional in mammalian cells. J. Virol. 2001, 75, 1004–1012. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lu, G.; Li, J.; Zhou, Y.J.; Zhou, X.P.; Tao, X.R. Model-based structural and functional characterization of the Rice stripe tenuivirus nucleocapsid protein interacting with viral genomic RNA. Virology 2017, 506, 73–83. [Google Scholar] [CrossRef]
- Cowley, J.A. The genomes of Mourilyan virus and Wenzhou shrimp virus 1 of prawns comprise 4 RNA segments. Virus Res. 2021, 292. [Google Scholar] [CrossRef]
- Sicard, A.; Michalakis, Y.; Gutierrez, S.; Blanc, S. The Strange Lifestyle of Multipartite Viruses. PLoS Pathog. 2016, 12, e1005819. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yao, M.; Liu, X.; Li, S.; Xu, Y.; Zhou, Y.; Zhou, X.; Tao, X. Rice Stripe Tenuivirus NSvc2 Glycoproteins Targeted to Golgi Body by N-Terminal Transmembrane Domain and Adjacent Cytosolic 24 Amino-Acids via COP I- and COP II-Dependent Secretion Pathway. J. Virol. 2014, 88, 3223–3234. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lu, G.; Li, S.; Zhou, C.W.; Qian, X.; Xiang, Q.; Yang, T.Q.; Wu, J.X.; Zhou, X.P.; Zhou, Y.J.; Ding, X.S.; et al. Tenuivirus utilizes its glycoprotein as a helper component to overcome insect midgut barriers for its circulative and propagative transmission. PLoS Pathog. 2019, 15, e1007655. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ren, F.L.; Zhou, M.; Deng, F.; Wang, H.L.; Ning, Y.J. Combinatorial Minigenome Systems for Emerging Banyangviruses Reveal Viral Reassortment Potential and Importance of a Protruding Nucleotide in Genome “Panhandle” for Promoter Activity and Reassortment. Frontiers Microbiol. 2020, 11, 599. [Google Scholar] [CrossRef] [Green Version]
- Jiang, S.S.; Lu, Y.W.; Li, K.F.; Lin, L.; Zheng, H.Y.; Yan, F.; Chen, J.P. Heat shock protein 70 is necessary for Rice stripe virus infection in plants. Mol. Plant. Pathol. 2014, 15, 907–917. [Google Scholar] [CrossRef]
- Zhao, W.; Wang, Q.S.; Xu, Z.T.; Liu, R.Y.; Qui, F. Distinct replication and gene expression strategies of the Rice Stripe virus in vector insects and host plants. J. Gen. Virol. 2019, 100, 877–888. [Google Scholar] [CrossRef]
- Flick, K.; Katz, A.; Overby, A.; Feldmann, H.; Pettersson, R.F.; Flick, R. Functional analysis of the noncoding regions of the Uukuniemi virus (Bunyaviridae) RNA segments. J. Virol. 2004, 78, 11726–11738. [Google Scholar] [CrossRef] [Green Version]
- Walter, C.T.; Barr, J.N. Bunyamwera virus can repair both insertions and deletions during RNA replication. RNA 2010, 16, 1138–1145. [Google Scholar] [CrossRef] [Green Version]
- Zhao, W.; Xu, Z.T.; Zhang, X.M.; Yang, M.L.; Kang, L.; Liu, R.Y.; Cui, F. Genomic variations in the 3’-termini of Rice stripe virus in the rotation between vector insect and host plant. New Phytol. 2018, 219, 1085–1096. [Google Scholar] [CrossRef] [Green Version]
- Zhao, W.; Yang, P.C.; Kang, L.; Cui, F. Different pathogenicities of Rice stripe virus from the insect vector and from viruliferous plants. New Phytol. 2016, 210, 196–207. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, W.; Yu, J.; Jiang, F.; Wang, W.; Kang, L.; Cui, F. Coordination between terminal variation of the viral genome and insect microRNAs regulates rice stripe virus replication in insect vectors. PLoS Pathog. 2021, 17, e1009424. [Google Scholar] [CrossRef]
- Olschewski, S.; Cusack, S.; Rosenthal, M. The Cap-Snatching Mechanism of Bunyaviruses. Trends Microbiol. 2020, 28, 293–303. [Google Scholar] [CrossRef] [Green Version]
- Liu, X.J.; Jin, J.; Qiu, P.; Gao, F.L.; Lin, W.Z.; Xie, G.H.; He, S.M.; Liu, S.M.; Du, Z.G.; Wu, Z.J. Rice Stripe Tenuivirus Has a Greater Tendency To Use the Prime-and-Realign Mechanism in Transcription of Genomic than in Transcription of Antigenomic Template RNAs. J. Virol. 2018, 92, e01414-17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yao, M.; Zhang, T.; Zhou, T.; Zhou, Y.; Zhou, X.; Tao, X. Repetitive prime-and-realign mechanism converts short capped RNA leaders into longer ones that may be more suitable for elongation during rice stripe virus transcription initiation. J. Gen. Virol. 2012, 93, 194–202. [Google Scholar] [CrossRef]
- Zhao, S.L.; Gu, X.M.; Li, J.; Liang, C.Y. The N-terminal cysteine protease domain of rice stripe tenuivirus Pc1 possesses deubiquitinating enzyme activity. Virus Genes 2021, 57, 117–120. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Zhou, Z.; Shen, M.; Ge, L.; Liu, F. Ubiquitin-Conjugating Enzyme E2 E Inhibits the Accumulation of Rice Stripe Virus in Laodelphax striatellus (Fallén). Viruses 2020, 12, 908. [Google Scholar] [CrossRef] [PubMed]
- Xiong, G.H.; Liu, X.J.; Qiu, P.; Wu, X.Y.; Du, Z.G.; Zhang, J.; Yang, L.; Wu, Z.J. Rice grassy stunt virus p5 interacts with two protein components of the plant-specific CBL-CIPK Ca+2 signaling network of rice. Virus Genes 2017, 53, 446–453. [Google Scholar] [CrossRef]
- Zhang, H.H.; Li, L.L.; He, Y.Q.; Qin, Q.Q.; Chen, C.H.; Wei, Z.Y.; Tan, X.X.; Xie, K.L.; Zhang, R.F.; Hong, G.J.; et al. Distinct modes of manipulation of rice auxin response factor OsARF17 by different plant RNA viruses for infection. Proc. Nat. Ac. Sci. USA 2020, 117, 9112–9121. [Google Scholar] [CrossRef]
- Hu, J.L.; Huang, J.; Xu, H.S.; Wang, Y.S.; Li, C.; Wen, P.Z.; You, X.M.; Zhang, X.; Pan, G.; Li, Q.; et al. Rice stripe virus suppresses jasmonic acid-mediated resistance by hijacking brassinosteroid signaling pathway in rice. PLoS Pathog. 2020, 16, e1008801. [Google Scholar] [CrossRef] [PubMed]
- Satoh, K.; Yoneyama, K.; Kondoh, H.; Shimizu, T.; Sasaya, T.; Choi, I.R.; Omura, T.; Kikuchi, S. Relationship between gene responses and symptoms induced by Rice grassy stunt virus. Front. Microbiol. 2013, 4, 313. [Google Scholar] [CrossRef] [Green Version]
- Zhang, C.; Wei, Y.; Xu, L.; Wu, K.C.; Yang, L.; Shi, C.N.; Yang, G.Y.; Chen, D.; Yu, F.F.; Xie, Q.; et al. A Bunyavirus-Inducible Ubiquitin Ligase Targets RNA Polymerase IV for Degradation during Viral Pathogenesis in Rice. Mol. Plant. 2020, 13, 836–850. [Google Scholar] [CrossRef] [PubMed]
- Shi, B.B.; Lin, L.; Wang, S.H.; Guo, Q.; Zhou, H.; Rong, L.L.; Li, J.M.; Peng, J.J.; Lu, Y.W.; Zheng, H.Y.; et al. Identification and regulation of host genes related to Rice stripe virus symptom production. New Phytol. 2016, 209, 1106–1119. [Google Scholar] [CrossRef] [PubMed]
- Zhao, J.P.; Zhang, X.; Hong, Y.G.; Liu, Y.L. Chloroplast in Plant-Virus Interaction. Front. Microbiol. 2016, 7, 1565. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kong, L.F.; Wu, J.X.; Lu, L.N.; Xu, Y.; Zhou, X.P. Interaction between Rice stripe virus Disease-Specific Protein and Host PsbP Enhances Virus Symptoms. Mol. Plant. 2014, 7, 691–708. [Google Scholar] [CrossRef] [Green Version]
- Xu, Y.; Zhou, X.P. Role of rice stripe virus NSvc4 in cell-to-cell movement and symptom development in Nicotiana benthamiana. Front. Plant. Sci. 2012, 3, 269. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, J.P.; Xu, J.J.; Chen, B.H.; Cui, W.J.; Zhou, Z.J.; Song, X.J.; Chen, Z.; Zheng, H.Y.; Lin, L.; Peng, J.J.; et al. Characterization of Proteins Involved in Chloroplast Targeting Disturbed by Rice Stripe Virus by Novel Protoplast-Chloroplast Proteomics. Int. J. Mol. Sci. 2019, 20, 253. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Patil, B.L.; Kumar, P.L. Pigeonpea sterility mosaic virus: A legume-infecting Emaravirus from South Asia. Mol. Plant Pathol. 2015, 16, 775–786. [Google Scholar] [CrossRef]
- Zheng, Y.; Navarro, B.; Wang, G.; Wang, Y.; Yang, Z.; Xu, W.; Zhu, C.; Wang, L.; Serio, F.D.; Hong, N. Actinidia chlorotic ringspot-associated virus: A novel emaravirus infecting kiwifruit plants. Mol. Plant. Pathol. 2017, 18, 569–581. [Google Scholar] [CrossRef]
- Walia, J.J.; Falk, B.W. Fig mosaic virus mRNAs show generation by cap-snatching. Virology 2012, 426, 162–166. [Google Scholar] [CrossRef] [Green Version]
- Huazhcn, E.; Wang, G.; Yang, Z.; Wang, Y.; Zhang, Z.; Lj, L.; Waqas, M.; Hong, N.; Liu, H.; Wang, G.; et al. Identification and characterization of a pear chiorotic leaf spot-associated virus, a novel emaravirus associated with a severe disease of pear trees in China. Plant Dis. 2020, 104, 2786–2798. [Google Scholar] [CrossRef]
- Verchot, J.; Herath, V.; Urrutia, C.D.; Gayral, M.; Lyle, K.; Shires, M.K.; Ong, K.; Byrne, D. Development of a reverse genetic system for studying rose rosette virus in whole plants. Mol. Plant. Microbe Interact. 2020, 33, 1209–1221. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Subbarao, B.; Hallan, V. Molecular characterization of emaraviruses associated with Pigeonpea sterility mosaic disease. Sci. Rep. 2017, 7, 1–20. [Google Scholar] [CrossRef] [Green Version]
- Yang, C.; Zhang, S.; Han, T.; Fu, J.; Di Serio, F.; Cao, M. Identification and characterization of a novel emaravirus associated with jujube (Ziziphus jujuba Mill.) yellow mottle disease. Front. Microbiol. 2019. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kraatz, F.; Wernike, K.; Reiche, S.; Aebischer, A.; Reimann, I.; Beer, M. Schmallenberg virus non-structural protein NSm: Intracellular distribution and role of non-hydrophobic domains. Virology 2018, 516, 46–54. [Google Scholar] [CrossRef] [PubMed]
- Elbeaino, T.; Digiaro, M.; Martelli, G.P. Complete nucleotide sequence of four RNA segments of fig mosaic virus. Arch. Virol. 2009, 154, 1719–1727. [Google Scholar] [CrossRef] [PubMed]
- Elbeaino, T.; Digiaro, M.; Uppala, M.; Sudini, H. Deep sequencing of dsRNAs recovered from mosaic-diseased pigeonpea reveals the presence of a novel emaravirus: Pigeonpea sterility mosaic virus 2. Arch. Virol. 2015. [Google Scholar] [CrossRef]
- Elbeaino, T.; Digiaro, M.; Uppala, M.; Sudini, H. Deep sequencing of pigeonpea sterility mosaic virus discloses five RNA segments related to emaraviruses. Virus Res. 2014, 188, 27–31. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ishikawa, K.; Miura, C.; Maejima, K.; Komatsu, K.; Hashimoto, M.; Tomomitsu, T.; Fukuoka, M.; Yusa, A.; Yamaji, Y.; Namba, S. Nucleocapsid protein from Fig mosaic virus forms cytoplasmic agglomerates that are hauled by endoplasmic reticulum streaming. J. Virol. 2015, 89, 480–491. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Buzkan, N.; Chiumenti, M.; Massart, S.; Sarpkaya, K.; Karadağ, S.; Minafra, A. A new emaravirus discovered in Pistacia from Turkey. Virus Res. 2019, 263, 159–163. [Google Scholar] [CrossRef] [PubMed]
- Kubota, K.; Usugi, T.; Tomitaka, Y.; Shimomoto, Y.; Takeuchi, S.; Kadono, F.; Yanagisawa, H.; Chiaki, Y.; Tsuda, S. Perilla mosaic virus is a highly divergent emaravirus transmitted by Shevtchenkella sp.(Acari: Eriophyidae). Phytopathology 2020, 110, 1352–1361. [Google Scholar] [CrossRef] [PubMed]
- Tatineni, S.; McMechan, A.J.; Wosula, E.N.; Wegulo, S.N.; Graybosch, R.A.; French, R.; Hein, G.L. An eriophyid mite-transmitted plant virus contains eight genomic RNA segments with unusual heterogeneity in the nucleocapsid protein. J. Virol. 2014, 88, 11834–11845. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ishikawa, K.; Maejima, K.; Komatsu, K.; Netsu, O.; Keima, T.; Shiraishi, T.; Okano, Y.; Hashimoto, M.; Yamaji, Y.; Namba, S. Fig mosaic emaravirus p4 protein is involved in cell-to-cell movement. J. Gen. Virol. 2013, 94, 682–686. [Google Scholar] [CrossRef] [PubMed]
- McGavin, W.J.; Mitchell, C.; Cock, P.J.A.; Wright, K.M.; MacFarlane, S.A. Raspberry leaf blotch virus, a putative new member of the genus Emaravirus, encodes a novel genomic RNA. J. Gen. Virol. 2012, 93. [Google Scholar] [CrossRef] [PubMed]
- Yu, C.; Karlin, D.G.; Lu, Y.; Wright, K.; Chen, J.; MacFarlane, S. Experimental and bioinformatic evidence that raspberry leaf blotch emaravirus P4 is a movement protein of the 30K superfamily. J. Gen. Virol. 2013, 94, 2117–2128. [Google Scholar] [CrossRef]
- Di Bello, P.L.; Ho, T.; Tzanetakis, I.E. The evolution of emaraviruses is becoming more complex: Seven segments identified in the causal agent of Rose rosette disease. Virus Res. 2015, 210, 241–244. [Google Scholar] [CrossRef]
- Elbeaino, T.; Marais, A.; Faure, C.; Trioano, E.; Candresse, T.; Parrella, G. High-Throughput Sequencing Reveals Cyclamen persicum Mill. as a Natural Host for Fig Mosaic Virus. Viruses 2018, 10, 684. [Google Scholar] [CrossRef] [Green Version]
- Samarfard, S.; McTaggart, A.R.; Sharman, M.; Bejerman, N.E.; Dietzgen, R.G. Viromes of Ten Alfalfa Plants in Australia Reveal Diverse Known Viruses and a Novel RNA Virus. Pathogens 2020, 9, 214. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- von Bargen, S.; Al Kubrusli, R.; Gaskin, T.; Fürl, S.; Hüttner, F.; Blystad, D.; Karlin, D.G.; Jalkanen, R.; Büttner, C. Characterisation of a novel Emaravirus identified in mosaic-diseased Eurasian aspen (Populus tremula). Ann. Appl. Biol. 2020, 176, 210–222. [Google Scholar] [CrossRef]
- Desbiez, C.; Moury, B.; Lecoq, H. The hallmarks of “green” viruses: Do plant viruses evolve differently from the others? Inf. Genet. Evol. 2011, 11, 812–824. [Google Scholar] [CrossRef]
- Storms, M.M.H.; Kormelink, R.; Peters, D.; van Lent, J.W.M.; Goldbach, R.W. The nonstructural NSm protein of Tomato spotted wilt virus induces tubular structures in plant and insect cells. Virology 1995, 214, 485–493. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Soellick, T.R.; Uhrig, J.F.; Bucher, G.L.; Kellmann, J.W.; Schreier, P.H. The movement protein NSm of tomato spotted wilt tospovirus (TSWV): RNA binding, interaction with the TSWV N protein, and identification of interacting plant proteins. Proc. Nat. Acad. Sci. USA 2000, 97, 2373–2378. [Google Scholar] [CrossRef] [Green Version]
- Tripathi, D.; Raikhy, G.; Goodin, M.M.; Dietzgen, R.G.; Pappu, H.R. In Vivo Localization of Iris yellow spot Tospovirus (Bunyaviridae)-Encoded Proteins and Identification of Interacting Regions of Nucleocapsid and Movement Proteins. PLoS ONE 2015, 10, e0118973. [Google Scholar] [CrossRef] [PubMed]
- Tripathi, D.; Raikhy, G.; Pappu, H.R. Movement and nucleocapsid proteins coded by two tospovirus species interact through multiple binding regions in mixed infections. Virology 2015, 478, 137–147. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leastro, M.O.; Pallas, V.; Resende, R.O.; Sanchez-Navarro, J.A. The movement proteins (NSm) of distinct tospoviruses peripherally associate with cellular membranes and interact with homologous and heterologous NSm and nucleocapsid proteins. Virology 2015, 478, 39–49. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leastro, M.O.; Pallás, V.; Resende, R.O.; Sánchez-Navarro, J.A. The functional analysis of distinct tospovirus movement proteins (NSM) reveals different capabilities in tubule formation, cell-to-cell and systemic virus movement among the tospovirus species. Virus Res. 2017, 227, 57–68. [Google Scholar] [CrossRef] [PubMed]
- Storms, M.M.H.; van der schoot, C.; Prins, M.; Kormelink, R.; van Lent, J.W.M.; Goldbach, R.W. A comparison of two methods of microinjection for assessing altered plasmodesmal gating in tissues expressing viral movement proteins. Plant J. 1998, 13, 131–140. [Google Scholar] [CrossRef]
- Qian, X.; Xiang, Q.; Yang, T.; Ma, H.; Ding, X.S.; Tao, X. Molecular Co-Chaperone SGT1 is Critical for Cell-to-Cell Movement and Systemic Infection of Tomato Spotted Wilt Virus in Nicotiana benthamiana. Viruses 2018, 10, 647. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Paape, M.; Solovyev, A.G.; Erokhina, T.N.; Minina, E.A.; Schepetilnikov, M.V.; Lesemann, D.-E.; Schiemann, J.E.; Morozov, S.Y.; Kellmann, J.-W. At-4/1, an interactor of the Tomato spotted wilt virus movement protein, belongs to a new family of plant proteins capable of directed intra- and intercellular trafficking. Mol. Plant Micr. Int. 2006, 19, 874–883. [Google Scholar] [CrossRef] [Green Version]
- Feng, Z.K.; Xue, F.; Xu, M.; Chen, X.J.; Zhao, W.Y.; Garcia-Murria, M.J.; Mingarro, I.; Liu, Y.; Huang, Y.; Jiang, L.; et al. The ER-membrane transport system is critical for intercellular trafficking of the NSm movement Protein and Tomato Spotted Wilt Tospovirus. PLoS Pathog. 2016, 12, e1005443. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lewandowski, D.J.; Adkins, S. The tubule-forming NSm protein from Tomato spotted wilt virus complements cell-to-cell and long-distance movement of Tobacco mosaic virus hybrids. Virology 2005, 342, 26–37. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shen, Y.; Zhao, X.; Yao, M.; Li, C.; Miriam, K.; Zhang, X.; Tao, X. A versatile complementation assay for cell-to-cell and long distance movements by cucumber mosaic virus based agro-infiltration. Virus Res. 2014, 190, 25–33. [Google Scholar] [CrossRef] [PubMed]
- Hollidge, B.S.; Weiss, S.R.; Soldan, S.S. The role of interferon antagonist, non-structural proteins in pathogenesis and emergence of arboviruses. Viruses 2011, 3, 629–648. [Google Scholar] [CrossRef]
- Bucher, E.; Sijen, T.; de Haan, P.; Goldbach, R.; Prins, M. Negative-strand tospoviruses and tenuiviruses carry a gene for a suppressor of gene silencing at analogous genomic positions. J. Virol. 2003, 77, 1329–1336. [Google Scholar] [CrossRef] [Green Version]
- Takeda, A.; Sugiyama, K.; Nagano, H.; Mori, M.; Kaido, M.; Mise, K.; Tsuda, S.; Okuno, T. Identification of a novel RNA silencing suppressor, NSs protein of Tomato spotted wilt virus. FEBS Lett. 2002, 532, 75–79. [Google Scholar] [CrossRef] [Green Version]
- Hedil, M.; Sterken, M.; de Ronde, D.; Lohuis, D.; Kormelink, R. Analysis of Tospovirus NSs Proteins in Suppression of Systemic Silencing. PLoS ONE 2015, 10, e0134517. [Google Scholar] [CrossRef]
- Goswami, S.; Sahana, N.; Pandey, V.; Doblas, P.; Jain, R.K.; Palukaitis, P.; Canto, T.; Praveen, S. Interference in plant defense and development by non-structural protein NSs of Groundnut bud necrosis virus. Virus Res. 2012, 163, 368–373. [Google Scholar] [CrossRef] [Green Version]
- Huang, C.H.; Hsiao, W.R.; Huang, C.W.; Chen, K.C.; Lin, S.S.; Chen, T.C.; Raja, J.A.; Wu, H.W.; Yeh, S.D. Two Novel Motifs of Watermelon Silver Mottle Virus NSs Protein Are Responsible for RNA Silencing Suppression and Pathogenicity. PLoS ONE 2015, 10, e0126161. [Google Scholar] [CrossRef]
- Schnettler, E.; Hemmes, H.; Huismann, R.; Goldbach, R.; Prins, M.; Kormelink, R. Diverging affinity of tospovirus RNA silencing suppressor proteins, NSs, for various RNA duplex molecules. J. Virol. 2010, 84, 11542–11554. [Google Scholar] [CrossRef] [Green Version]
- Hedil, M.; de Ronde, D.; Kormelink, R. Biochemical analysis of NSs from different tospoviruses. Virus Res. 2017, 242, 149–155. [Google Scholar] [CrossRef]
- de Ronde, D.; Pasquier, A.; Ying, S.; Butterbach, P.; Lohuis, D.; Kormelink, R. Analysis of Tomato spotted wilt virus NSs protein indicates the importance of the N-terminal domain for avirulence and RNA silencing suppression. Mol. Plant. Pathol. 2014, 15, 185–195. [Google Scholar] [CrossRef]
- Giner, A.; Lakatos, L.; García-Chapa, M.; López-Moya, J.J.; Burgyán, J. Viral Protein Inhibits RISC Activity by Argonaute Binding through Conserved WG/GW Motifs. PLoS Pathog 2010, 6, e1000996. [Google Scholar] [CrossRef] [Green Version]
- Huang, C.H.; Foo, M.H.; Raja, J.A.J.; Tan, Y.R.; Lin, T.T.; Lin, S.S.; Yeh, S.D. A Conserved Helix in the C-Terminal Region of Watermelon Silver Mottle Virus Nonstructural Protein S Is Imperative For Protein Stability Affecting Self-Interaction, RNA Silencing Suppression, and Pathogenicity. Mol. Plant. Microbe Interact. 2020, 33, 637–652. [Google Scholar] [CrossRef] [PubMed]
- Bhushan, L.; Abraham, A.; Choudhury, N.R.; Rana, V.S.; Mukherjee, S.K.; Savithri, H.S. Demonstration of helicase activity in the nonstructural protein, NSs, of the negative-sense RNA virus, groundnut bud necrosis virus. Arch. Virol. 2015, 160, 959–967. [Google Scholar] [CrossRef] [PubMed]
- Lokesh, B.; Rashmi, P.R.; Amruta, B.S.; Srisathiyanarayanan, D.; Murthy, M.R.N.; Savithri, H.S.; Handanahal, S. NSs encoded by groundnut bud necrosis virus Is a bifunctional enzyme. PLoS ONE 2010, 5, e9757. [Google Scholar] [CrossRef]
- Ocampo Ocampo, T.; Gabriel Peralta, S.M.; Bacheller, N.; Uiterwaal, S.; Knapp, A.; Hennen, A.; Ochoa-Martinez, D.L.; Garcia-Ruiz, H. Antiviral RNA silencing suppression activity of Tomato spotted wilt virus NSs protein. Genet. Mol. Res. 2016, 15. [Google Scholar] [CrossRef]
- Garcia-Ruiz, H.; Peralta, S.M.G.; Harte-Maxwell, P.A. Tomato spotted wilt virus NSs protein supports infection and systemic movement of a potyvirus and is a symptom determinant. Viruses 2018, 10, 129. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hassani-Mehraban, A.; Brenkman, A.B.; van den Broek, N.J.F.; Goldbach, R.; Kormelink, R. RNAi-Mediated Transgenic Tospovirus Resistance Broken by Intraspecies Silencing Suppressor Protein Complementation. Mol. Plant Microb. Int. 2009, 10, 1250–1257. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bag, S.; Mitter, N.; Eid, S.; Pappu, H.R. Complementation between two tospoviruses facilitates the systemic movement of a plant virus silencing suppressor in an otherwise restrictive host. PLoS ONE 2012, 7, e44803. [Google Scholar] [CrossRef] [PubMed]
- Kormelink, R.; Kitajima, E.W.; de Haan, P.; Zuidema, D.; Peters, D.; Goldbach, R. The nonstructural protein (NSs) encoded by the ambisense S RNA of Tomato spotted wilt virus is associated with fibrous structures in infected plant cells. Virology 1991, 181, 459–468. [Google Scholar] [CrossRef]
- Wijkamp, I.; van Lent, J.; Kormelink, R.; Goldbach, R.; Peters, D. Multiplication of Tomato spotted wilt virus in its insect vector, Frankliniella occidentalis. Phytopathology 1993, 83, 986–991. [Google Scholar] [CrossRef]
- de Assis Filho, F.M.; Naidu, R.A.; Deom, C.M.; Sherwood, J.L. Dynamics of Tomato spotted wilt virus Replication in the Alimentary Canal of Two Thrips Species. Phytopathology 2002, 92, 729–733. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kritzman, A.; Gera, A.; Raccah, B.; van Lent, J.W.; Peters, D. The route of tomato spotted wilt virus inside the thrips body in relation to transmission efficiency. Arch. Virol. 2002, 147, 2143–2156. [Google Scholar] [CrossRef] [PubMed]
- Nagata, T.; Inoue-Nagata, A.K.; Smid, H.M.; Goldbach, R.; Peters, D. Tissue tropism related to vector competence of Frankliniella occidentalis for tomato spotted wilt tospovirus. J. Gen. Virol. 1999, 80, 507–515. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Montero-Astúa, M.; Ullman, D.E.; Whitfield, A.E. Salivary gland morphology, tissue tropism and the progression of tospovirus infection in Frankliniella occidentalis. Virology 2016, 493, 39–51. [Google Scholar] [CrossRef]
- Ghosh, A.; Priti Mandal, B.; Dietzgen, R.G. Progression of Watermelon Bud Necrosis Virus Infection in Its Vector, Thrips palmi. Cells 2021, 10, 392. [Google Scholar] [CrossRef]
- Storms, M.M.; Nagata, T.; Kormelink, R.; Goldbach, R.W.; van Lent, J.W. Expression of the movement protein of Tomato spotted wilt virus in its insect vector Frankliniella occidentalis. Arch. Virol. 2002, 147, 825–831. [Google Scholar] [CrossRef]
- Zhang, C.; Pei, X.W.; Wang, Z.X.; Jia, S.R.; Guo, S.W.; Zhang, Y.Q.; Li, W.M. The Rice stripe virus pc4 functions in movement and foliar necrosis expression in Nicotiana benthamiana. Virology 2012, 425, 113–121. [Google Scholar] [CrossRef] [Green Version]
- Rong, L.L.; Lu, Y.W.; Lin, L.; Zheng, H.Y.; Yan, F.; Chen, J.P. A transmembrane domain determines the localization of rice stripe virus pc4 to plasmodesmata and is essential for its function as a movement protein. Virus Res. 2021, 183, 112–116. [Google Scholar] [CrossRef]
- Fu, S.; Xu, Y.; Li, C.Y.; Li, Y.; Wu, J.X.; Zhou, X.P. Rice Stripe Virus Interferes with S-acylation of Remorin and Induces Its Autophagic Degradation to Facilitate Virus Infection. Mol. Plant. 2018, 11, 269–287. [Google Scholar] [CrossRef] [Green Version]
- Jarsch, I.K.; Ott, T. Perspectives on Remorin Proteins, Membrane Rafts, and Their Role during Plant–Microbe Interactions. Mol. Plant Microbe Int. 2011, 24, 7–12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, Y. Remorins: Essential Regulators in Plant-Microbe Interaction and Cell Death Induction. Plant Physiol. 2020, 183, 435–436. [Google Scholar] [CrossRef] [PubMed]
- Cheng, G.Y.; Yang, Z.T.; Zhang, H.; Zhang, J.S.; Xu, J.S. Remorin interacting with PCaP1 impairs Turnip mosaic virus intercellular movement but is antagonised by VPg. New Phytol. 2020, 225, 2122–2139. [Google Scholar] [CrossRef] [PubMed]
- Hiraguri, A.; Netsu, O.; Shimizu, T.; Uehara-Ichiki, T.; Omura, T.; Sasaki, N.; Nyunoya, H.; Sasaya, T. The nonstructural protein pC6 of rice grassy stunt virus trans-complements the cell-to-cell spread of a movement-defective tomato mosaic virus. Arch. Virol. 2011, 156, 911–916. [Google Scholar] [CrossRef] [PubMed]
- Yuan, Z.; Chen, H.; Chen, Q.; Omura, T.; Xie, L.; Wu, Z.; Wei, T. The early secretory pathway and an actin-myosin VIII motility system are required for plasmodesmatal localization of the NSvc4 protein of Rice stripe virus. Vir. Res. 2011, 159, 62–68. [Google Scholar] [CrossRef] [PubMed]
- Sui, X.L.; Liu, X.J.; Lin, W.W.; Wu, Z.J.; Yang, L. Targeting of rice grassy stunt virus pc6 protein to plasmodesmata requires the ER-to-Golgi secretory pathway and an actin-myosin VIII motility system. Arch. Virol. 2018, 163, 1317–1323. [Google Scholar] [CrossRef]
- Zheng, L.P.; Du, Z.G.; Lin, C.; Mao, Q.Z.; Wu, K.C.; Wu, J.G.; Wei, T.Y.; Wu, Z.J.; Xie, L.H. Rice stripe tenuivirus p2 may recruit or manipulate nucleolar functions through an interaction with fibrillarin to promote virus systemic movement. Mol. Plant. Pathol. 2015, 16, 921–930. [Google Scholar] [CrossRef]
- Zheng, L.P.; Hong, P.X.; Guo, X.N.; Li, Y.; Xie, L. Rice stripe virusp2 Colocalizes and Interacts with Arabidopsis Cajal Bodies and Its Domains in Plant Cells. Biomed. Res. Int. 2020. [Google Scholar] [CrossRef]
- Hemmes, H.; Lakatos, L.; Goldbach, R.; Burgyan, J.; Prins, M. The NS3 protein of Rice hoja blanca tenuivirus suppresses RNA silencing in plant and insect hosts by efficiently binding both siRNAs and miRNAs. RNA 2007, 13, 1079–1089. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xiong, R.; Wu, J.; Zhou, Y.; Zhou, X. Characterization and subcellular localization of an RNA silencing suppressor encoded by Rice stripe tenuivirus. Virology 2009, 387, 29–40. [Google Scholar] [CrossRef] [Green Version]
- Schnettler, E.; de Vries, W.; Hemmes, H.; Haasnoot, J.; Kormelink, R.; Goldbach, R.; Berkhout, B. The NS3 protein of rice hoja blanca virus complements the RNAi suppressor function of the HIV-1 Tat. EMBO Rep. 2009, 10, 258–263. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, H.; Cho, W.K.; Lian, S.; Kim, K.H. Identification of residues or motif(s) of the rice stripe virus p3 protein required for self-interaction and for silencing suppressor activity. Virus Res. 2017, 235, 14–23. [Google Scholar] [CrossRef] [PubMed]
- Hemmes, H.; Kaaij, L.; Lohuis, D.; Prins, M.; Goldbach, R.; Schnettler, E. Binding of siRNA molecules is crucial for RNAi suppressor activity of Rice hoja blanca virus NS3 in plants. J. Gen. Virol. 2009, 90, 1762–1766. [Google Scholar] [CrossRef] [PubMed]
- Shen, M.; Xu, Y.; Jia, R.; Zhou, X.; Ye, K. Size-independent and noncooperative recognition of dsRNA by the Rice stripe virus RNA silencing suppressor NS3. J. Mol. Biol. 2010, 404, 665–679. [Google Scholar] [CrossRef] [PubMed]
- Wu, G.T.; Zheng, G.X.; Hu, Q.; Ma, M.G.; Li, M.J.; Su, X.C.; Yan, F.; Qing, L. p3 Protein from Rice stripe virus affects the expression of endogenous genes in Nicotiana benthamiana. Virology J. 2018, 15, 105. [Google Scholar] [CrossRef] [Green Version]
- Zheng, L.J.; Zhang, C.; Shi, C.N.; Yang, Z.R.; Wang, Y.; Zhou, T.; Sun, F.; Wang, H.; Zhao, S.S.; Qin, Q.Q.; et al. Rice stripe virus p3 protein regulates primary miRNA processing through association with the miRNA biogenesis factor OsDRB1 and facilitates virus infection in rice. PLoS Pathog. 2017, 13, e1006662. [Google Scholar] [CrossRef] [Green Version]
- Chen, B.H.; Lin, L.; Lu, Y.W.; Peng, J.J.; Zheng, H.Y.; Yang, Q.K.; Rao, S.F.; Wu, G.W.; Li, J.M.; Chen, Z.; et al. Ubiquitin-Like protein 5 interacts with the silencing suppressor p3 of rice stripe virus and mediates its degradation through the 26S proteasome pathway. PLoS Pathog. 2020, 16, e1008780. [Google Scholar] [CrossRef]
- Jiang, L.L.; Lu, Y.W.; Zheng, X.Y.; Yang, X.; Chen, Y.; Zhang, T.H.; Zhao, X.; Wang, S.; Song, X.J.; Zhang, X.X.; et al. The plant protein NbP3IP directs degradation ofRice stripe virusp3 silencing suppressor protein to limit virus infection through interaction with the autophagy-related protein NbATG8. New Phytol. 2021, 229, 1036–1051. [Google Scholar] [CrossRef]
- Du, Z.; Xiao, D.; Wu, J.; Jia, D.; Yuan, Z.; Liu, Y.; Hu, L.; Han, Z.; Wei, T.; Lin, Q.; et al. P2 of Rice stripe virus (RSV) interacts with OsSGS3 and is a silencing suppressor. Mol. Plant. Path. 2011, 12, 808–814. [Google Scholar] [CrossRef]
- Zhang, C.; Chen, D.; Yang, G.Y.; Yu, X.Y.; Wu, J.G. Rice Stripe Mosaic Virus-Encoded P4 Is a Weak Suppressor of Viral RNA Silencing and Is Required for Disease Symptom Development. Mol. Plant Microbe Int. 2020, 33, 412–422. [Google Scholar] [CrossRef]
- Zhang, C.; Liu, X.J.; Wu, K.C.; Zheng, L.P.; Ding, Z.M.; Li, F.; Zou, P.; Yang, L.; Wu, J.G.; Wu, Z.J. Rice grassy stunt virus nonstructural protein p5 serves as a viral suppressor of RNA silencing and interacts with nonstructural protein p3. Arch. Virol. 2015, 160, 2769–2779. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, T.D.; Lacombe, S.; Bangratz, M.; Ta, H.A.; Vinh, D.N.; Gantet, P.; Brugidou, C. p2 of Rice grassy stunt virus (RGSV) and p6 and p9 of Rice ragged stunt virus (RRSV) isolates from Vietnam exert suppressor activity on the RNA silencing pathway. Virus Genes 2015, 51, 267–275. [Google Scholar] [CrossRef] [PubMed]
- Netsu, O.; Hiraguri, A.; Uehara-Ichiki, T.; Komatsu, K.; Sasaya, T. Functional comparison of RNA silencing suppressor between the p5 protein of rice grassy stunt virus and the p3 protein of rice stripe virus. Virus Res. 2015, 203, 10–19. [Google Scholar] [CrossRef] [PubMed]
- Gupta, A.K.; Hein, G.L.; Tatineni, S. P7 and P8 proteins of High Plains wheat mosaic virus, a negative-strand RNA virus, employ distinct mechanisms of RNA silencing suppression. Virology 2019. [Google Scholar] [CrossRef] [Green Version]
- Babu, B.; Washburn, B.K.; Poduch, K.; Knox, G.W.; Paret, M.L. Identification and characterization of two novel genomic RNA segments RNA5 and RNA6 in rose rosette virus infecting roses. Acta Virol. 2016. [Google Scholar] [CrossRef]
- Moritz, G.; Kumm, S.; Mound, L. Tospovirus transmission depends on thrips ontogeny. Virus Res. 2004, 100, 143–149. [Google Scholar] [CrossRef]
- Mou, D.F.; Chen, W.T.; Li, W.H.; Chen, T.C.; Tseng, C.H.; Huang, L.H.; Peng, J.C.; Yeh, S.D.; Tsai, C.W. Transmission mode of watermelon silver mottle virus by Thrips palmi. PLoS ONE 2021, 16, e0247500. [Google Scholar] [CrossRef]
- Van de Wetering, F.; Goldbach, R.; Peters, D. Tomato spotted wilt tospovirus ingestion by first instar larvae of Frankliniella occidentalis is a prerequisite for transmission. Phytopathol. 1996, 86, 900–905. [Google Scholar] [CrossRef]
- Whitfield, A.E.; Ullman, D.E.; German, T.L. Tospovirus-thrips interactions. Ann. Rev. Phytopathol. 2005, 43, 459–489. [Google Scholar] [CrossRef]
- Cortez, I.; Aires, A.; Pereira, A.-M.; Goldbach, R.; Peters, D.; Kormelink, R. Genetic organisation of Iris yellow spot virus M RNA: Indications for functional homology between the G(c) glycoproteins of tospoviruses and animal-infecting bunyaviruses. Arch. Virol. 2002, 147, 2313–2325. [Google Scholar] [CrossRef]
- Serris, A.; Stass, R.; Bignon, E.A.; Muena, N.A.; Manuguerra, J.C.; Jangra, R.K.; Li, S.; Chandran, K.; Tischler, N.D.; Huiskonen, J.T.; et al. The Hantavirus Surface Glycoprotein Lattice and Its Fusion Control Mechanism. Cell 2020, 183, 442–456.e16. [Google Scholar] [CrossRef]
- Bignon, E.A.; Albornoz, A.; Guardado-Calvo, P.; Rey, F.A.; Tischler, N.D. Molecular organization and dynamics of the fusion protein Gc at the hantavirus surface. eLife 2019, 8, e46028. [Google Scholar] [CrossRef] [PubMed]
- Cifuentes-Muñoz, N.; Salazar-Quiroz, N.; Tischler, N.D. Hantavirus Gn and Gc envelope glycoproteins: Key structural units for virus cell entry and virus assembly. Viruses 2014, 6, 1801–1822. [Google Scholar] [CrossRef] [Green Version]
- Willensky, S.; Bar-Rogovsky, H.; Bignon, E.A.; Tischler, N.D.; Modis, Y.; Dessau, M. Crystal Structure of Glycoprotein C from a Hantavirus in the Post-fusion Conformation. PLoS Pathog. 2016, 12, e1005948. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tischler, N.D.; Gonzalez, A.; Perez-Acle, T.; Rosemblatt, M.; Valenzuela, P.D.T. Hantavirus Gc glycoprotein: Evidence for a class II fusion protein. J. Gen. Virol. 2005, 86 Pt 11, 2937–2947. [Google Scholar] [CrossRef]
- Bahat, Y.; Alter, J.; Dessau, M. Crystal structure of tomato spotted wilt virus GN reveals a dimer complex formation and evolutionary link to animal-infecting viruses. Proc. Natl. Acad. Sci. USA 2020, 117, 26237–26244. [Google Scholar] [CrossRef]
- Ohnishi, J.; Knight, L.M.; Hosokawa, D.; Fujisawa, I.; Tsuda, S. Replication of Tomato spotted wilt virus After Ingestion by Adult Thrips setosus is Restricted to Midgut Epithelial Cells. Phytopathology 2001, 91, 1149–1155. [Google Scholar] [CrossRef] [Green Version]
- de Assis Filho, F.M.; Deom, C.M.; Sherwood, J.L. Acquisition of Tomato spotted wilt virus by Adults of Two Thrips Species. Phytopathology 2004, 94, 333–336. [Google Scholar] [CrossRef] [Green Version]
- Nagata, T.; Inoue-Nagata, A.K.; Prins, M.; Goldbach, R.; Peters, D. Impeded Thrips Transmission of Defective Tomato spotted wilt virus Isolates. Phytopathology 2000, 90, 454–459. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sin, S.H.; McNulty, B.C.; Kennedy, G.G.; Moyer, J.W. Viral genetic determinants for thrips transmission of Tomato spotted wilt virus. Proc. Natl. Acad. Sci. USA 2005, 102, 5168–5173. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Whitfield, A.E.; Kumar, N.K.; Rotenberg, D.; Ullman, D.E.; Wyman, E.A.; Zietlow, C.; Willis, D.K.; German, T.L. A soluble form of the Tomato spotted wilt virus (TSWV) glycoprotein G(N) (G(N)-S) inhibits transmission of TSWV by Frankliniella occidentalis. Phytopathology 2008, 98, 45–50. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Whitfield, A.E.; Ullman, D.E.; German, T.L. Expression and characterization of a soluble form of tomato spotted wilt virus glycoprotein GN. J. Virol. 2004, 78, 13197–13206. [Google Scholar] [CrossRef] [Green Version]
- Montero-Astúa, M.; Rotenberg, D.; Leach-Kieffaber, A.; Schneweis, B.A.; Park, S.; Park, J.K.; German, T.L.; Whitfield, A.E. Disruption of vector transmission by a plant-expressed viral glycoprotein. Mol. Plant. Microbe Interact. 2014, 27, 296–304. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, J.; Tzanetakis, I.E. Transmission blockage of an orthotospovirus using synthetic peptides. J. Gen. Virol. 2020, 101, 112–121. [Google Scholar] [CrossRef]
- Badillo-Vargas, I.E.; Chen, Y.; Martin, K.M.; Rotenberg, D.; Whitfield, A.E. Discovery of Novel Thrips Vector Proteins That Bind to the Viral Attachment Protein of the Plant Bunyavirus Tomato Spotted Wilt Virus. J. Virol. 2019, 93, e00699-19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leach, A.; Fuchs, M.; Harding, R.; Nault, B.A. Iris Yellow Spot Virus Prolongs the Adult Lifespan of Its Primary Vector, Onion Thrips (Thrips tabaci) (Thysanoptera: Thripidae). J. Insect Sci. 2019, 19, 8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maris, P.C.; Joosten, N.N.; Goldbach, R.W.; Peters, D. Tomato spotted wilt virus infection improves host suitability for its vector Frankliniella occidentalis. Phytopathology 2004, 94, 706–711. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nachappa, P.; Challacombe, J.; Margolies, D.C.; Nechols, J.R.; Whitfield, A.E.; Rotenberg, D. Tomato Spotted Wilt Virus Benefits Its Thrips Vector by Modulating Metabolic and Plant Defense Pathways in Tomato. Front. Plant. Sci. 2020, 11, 575564. [Google Scholar] [CrossRef] [PubMed]
- Shalileh, S.; Ogada, P.A.; Moualeu, D.P.; Poehling, H.M. Manipulation of Frankliniella occidentalis (Thysanoptera: Thripidae) by Tomato Spotted Wilt Virus (Tospovirus) Via the Host Plant Nutrients to Enhance Its Transmission and Spread. Environ. Entomol. 2016, 45, 1235–1242. [Google Scholar] [CrossRef] [Green Version]
- Stafford, C.A.; Walker, G.P.; Ullman, D.E. Infection with a plant virus modifies vector feeding behavior. Proc. Natl. Acad. Sci. USA 2011, 108, 9350–9355. [Google Scholar] [CrossRef] [Green Version]
- Wan, Y.; Hussain, S.; Merchant, A.; Xu, B.; Xie, W.; Wang, S.; Zhang, Y.; Zhou, X.; Wu, Q. Tomato spotted wilt orthotospovirus influences the reproduction of its insect vector, western flower thrips, Frankliniella occidentalis, to facilitate transmission. Pest. Manag. Sci. 2020, 76, 2406–2414. [Google Scholar] [CrossRef]
- Medeiros, R.B.; Resende, R.D.O.; de Avila, A.C. The plant virus Tomato Spotted Wilt Tospovirus activates the immune system of its main insect vector, Frankliniella occidentalis. J. Virol. 2004, 78, 4976–4982. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ogada, P.A.; Kiirika, L.M.; Lorenz, C.; Senkler, J.; Braun, H.P.; Poehling, H.M. Differential proteomics analysis of Frankliniella occidentalis immune response after infection with Tomato spotted wilt virus (Tospovirus). Dev. Comp. Immunol. 2017, 67, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Schneweis, D.J.; Whitfield, A.E.; Rotenberg, D. Thrips developmental stage-specific transcriptome response to tomato spotted wilt virus during the virus infection cycle in Frankliniella occidentalis, the primary vector. Virology 2017, 500, 226–237. [Google Scholar] [CrossRef]
- Shrestha, A.; Champagne, D.E.; Culbreath, A.K.; Rotenberg, D.; Whitfield, A.E.; Srinivasan, R. Transcriptome changes associated with Tomato spotted wilt virus infection in various life stages of its thrips vector, Frankliniella fusca (Hinds). J. Gen. Virol. 2017, 98, 2156–2170. [Google Scholar] [CrossRef] [PubMed]
- Widana Gamage, S.M.K.; Rotenberg, D.; Schneweis, D.J.; Tsai, C.W.; Dietzgen, R.G. Transcriptome-wide responses of adult melon thrips (Thrips palmi) associated with capsicum chlorosis virus infection. PLoS ONE 2018, 13, e0208538. [Google Scholar] [CrossRef]
- Zhang, Z.; Zhang, P.; Li, W.; Zhang, J.; Huang, F.; Yang, J.; Bei, Y.; Lu, Y. De novo transcriptome sequencing in Frankliniella occidentalis to identify genes involved in plant virus transmission and insecticide resistance. Genomics 2013, 101, 296–305. [Google Scholar] [CrossRef] [Green Version]
- Rebijith, K.B.; Asokan, R.; Hande, H.R.; Krishna Kumar, N.K. The First Report of miRNAs from a Thysanopteran Insect, Thrips palmi Karny Using High-Throughput Sequencing. PLoS ONE 2016, 11, e0163635. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shrestha, A.; Champagne, D.E.; Culbreath, A.K.; Abney, M.R.; Srinivasan, R. Comparison of transcriptomes of an orthotospovirus vector and non-vector thrips species. PLoS ONE 2019, 14, e0223438. [Google Scholar] [CrossRef]
- Chen, W.T.; Tseng, C.H.; Tsai, C.W. Effect of watermelon silver mottle virus on the life history and feeding preference of Thrips palmi. PLoS ONE 2014, 9, e102021. [Google Scholar] [CrossRef]
- Daimei, G.; Raina, H.S.; Devi, P.P.; Saurav, G.K.; Renukadevi, P.; Malathi, V.G.; Senthilraja, C.; Mandal, B.; Rajagopal, R. Influence of Groundnut bud necrosis virus on the Life History Traits and Feeding Preference of Its Vector, Thrips palmi. Phytopathology 2017, 107, 1440–1445. [Google Scholar] [CrossRef] [Green Version]
- Abe, H.; Tomitaka, Y.; Shimoda, T.; Seo, S.; Sakurai, T.; Kugimiya, S.; Tsuda, S.; Kobayashi, M. Antagonistic plant defense system regulated by phytohormones assists interactions among vector insect, thrips and a tospovirus. Plant Cell Physiol. 2012, 53, 204–212. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Du, J.; Song, X.Y.; Shi, X.B.; Tang, X.; Chen, J.B.; Zhang, Z.H.; Chen, G.; Zhang, Z.; Zhou, X.G.; Liu, Y.; et al. NSs, the Silencing Suppressor of Tomato Spotted Wilt Orthotospovirus, Interferes with JA-Regulated Host Terpenoids Expression to Attract Frankliniella occidentalis. Front. Microbiol. 2020, 11, 590451. [Google Scholar] [CrossRef] [PubMed]
- Ben-Mahmoud, S.; Anderson, T.; Chappell, T.M.; Smeda, J.R.; Mutschler, M.A.; Kennedy, G.G.; De Jong, D.M.; Ullman, D.E. A thrips vector of tomato spotted wilt virus responds to tomato acylsugar chemical diversity with reduced oviposition and virus inoculation. Sci. Rep. 2019, 9, 17157. [Google Scholar] [CrossRef]
- Margaria, P.; Bosco, L.; Vallino, M.; Ciuffo, M.; Mautino, G.C.; Tavella, L.; Turina, M.M. The NSs Protein of Tomato Spotted Wilt Virus Is Required for Persistent Infection and Transmission by Frankliniella occidentalis. J. Virol. 2014, 88, 5788–5802. [Google Scholar] [CrossRef] [Green Version]
- Maris, P.C.; Joosten, N.N.; Peters, D.; Goldbach, R.W. Thrips Resistance in Pepper and Its Consequences for the Acquisition and Inoculation of Tomato spotted wilt virus by the Western Flower Thrips. Phytopathology 2003, 93, 96–101. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Srinivasan, R.; Abney, M.R.; Lai, P.C.; Culbreath, A.K.; Tallury, S.; Leal-Bertioli, S.C.M. Resistance to Thrips in Peanut and Implications for Management of Thrips and Thrips-Transmitted Orthotospoviruses in Peanut. Front. Plant. Sci. 2018, 9, 1604. [Google Scholar] [CrossRef] [PubMed]
- Sundaraj, S.; Srinivasan, R.; Culbreath, A.K.; Riley, D.G.; Pappu, H.R. Host plant resistance against tomato spotted wilt virus in peanut (Arachis hypogaea) and its impact on susceptibility to the virus, virus population genetics, and vector feeding behavior and survival. Phytopathology 2014, 104, 202–210. [Google Scholar] [CrossRef] [Green Version]
- Chatzivassiliou, E.K.; Peters, D.; Katis, N.I. The Efficiency by Which Thrips tabaci Populations Transmit Tomato spotted wilt virus Depends on Their Host Preference and Reproductive Strategy. Phytopathology 2002, 92, 603–609. [Google Scholar] [CrossRef] [Green Version]
- Jacobson, A.L.; Booth, W.; Vargo, E.L.; Kennedy, G.G. Thrips tabaci population genetic structure and polyploidy in relation to competency as a vector of tomato spotted wilt virus. PLoS ONE 2013, 8, e54484. [Google Scholar] [CrossRef] [Green Version]
- Jacobson, A.L.; Kennedy, G.G. Specific insect-virus interactions are responsible for variation in competency of different Thrips tabaci isolines to transmit different Tomato Spotted Wilt Virus isolates. PLoS ONE 2013, 8, e54567. [Google Scholar] [CrossRef] [PubMed]
- Linak, J.A.; Jacobson, A.L.; Sit, T.L.; Kennedy, G.G. Relationships of virus titers and transmission rates among sympatric and allopatric virus isolates and thrips vectors support local adaptation. Sci. Rep. 2020, 10, 7649. [Google Scholar] [CrossRef] [PubMed]
- Nagata, T.; Inoue-Nagata, A.K.; van Lent, J.; Goldbach, R.; Peters, D. Factors determining vector competence and specificity for transmission of Tomato spotted wilt virus. J. Gen. Virol. 2002, 83, 663–671. [Google Scholar] [CrossRef]
- Westmore, G.C.; Poke, F.S.; Allen, G.R.; Wilson, C.R. Genetic and host-associated differentiation within Thrips tabaci Lindeman (Thysanoptera: Thripidae) and its links to Tomato spotted wilt virus-vector competence. Heredity 2013, 111, 210–215. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ogada, P.A.; Debener, T.; Poehling, H.M. Inheritance genetics of the trait vector competence in Frankliniella occidentalis (Western flower thrips) in the transmission of Tomato spotted wilt virus. Ecol. Evol. 2016, 6, 7911–7920. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, Y.; Dessau, M.; Rotenberg, D.; Rasmussen, D.A.; Whitfield, A.E. Entry of bunyaviruses into plants and vectors. Adv. Virus Res. 2019, 104, 65–96. [Google Scholar]
- Wu, W.; Zheng, L.M.; Chen, H.Y.; Jia, D.S.; Li, F.; Wei, T.Y. Nonstructural Protein NS4 of Rice Stripe Virus Plays a Critical Role in Viral Spread in the Body of Vector Insects. PLoS ONE 2014, 9, e88636. [Google Scholar] [CrossRef]
- Zheng, L.M.; Mao, Q.Z.; Xie, L.H.; Wei, T.Y. Infection route of rice grassy stunt virus, a tenuivirus, in the body of its brown planthopper vector, Nilaparvata lugens (Hemiptera: Delphacidae) after ingestion of virus. Virus Res. 2014, 188, 170–173. [Google Scholar] [CrossRef]
- Deng, J.; Li, S.; Hong, J.; Ji, Y.; Zhou, Y. Investigation on subcellular localization of Rice stripe virus in its vector small brown planthopper by electron microscopy. Virol. J. 2013, 10, 310. [Google Scholar] [CrossRef] [Green Version]
- Whitfield, A.E.; Falk, B.W.; Rotenberg, D. Insect vector-mediated transmission of plant viruses. Virology 2015, 479, 278–289. [Google Scholar] [CrossRef] [Green Version]
- Liu, W.W.; Gray, S.; Huo, Y.; Li, L.; Wei, T.Y.; Wang, X.F. Proteomic Analysis of Interaction between a Plant Virus and Its Vector Insect Reveals New Functions of Hemipteran Cuticular Protein. Mol. Cell. Proteom. 2015, 14, 2229–2242. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huo, Y.; Liu, W.W.; Zhang, F.J.; Chen, X.Y.; Li, L.; Liu, Q.F.; Zhou, Y.J.; Wei, T.Y.; Fang, R.X.; Wang, X.F. Transovarial Transmission of a Plant Virus Is Mediated by Vitellogenin of Its Insect Vector. PLoS Pathog. 2014, 10, e1003949. [Google Scholar] [CrossRef]
- Huo, Y.; Yu, Y.L.; Chen, L.Y.; Li, Q.; Zhang, M.T.; Song, Z.Y.; Chen, X.Y.; Fang, R.X.; Zhang, L.L. Insect tissue-specific vitellogenin facilitates transmission of plant virus. PLoS Pathog. 2018, 14, e1006909. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huo, Y.; Yu, Y.L.; Liu, Q.; Liu, D.; Zhang, M.T.; Liang, J.N.; Chen, X.Y.; Zhang, L.L.; Fang, R.X. Rice stripe virus hitchhikes the vector insect vitellogenin ligand-receptor pathway for ovary entry. Philosoph. Trans. Royal Soc. B 2019, 374, 20180312. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Y.; Chen, D.Y.; Hu, J.; Zhang, K.; Kang, L.; Chen, Y.; Huang, L.J.; Zhang, L.; Xiang, Y.; Song, Q.S.; et al. The alpha-tubulin ofLaodelphax striatellusmediates the passage of rice stripe virus (RSV) and enhances horizontal transmission. PLoS Pathog. 2020, 16, e1008710. [Google Scholar] [CrossRef]
- Wang, X.; Wang, W.; Zhang, W.Z.; Li, J.; Cui, F.; Qiao, L.Q. Immune function of an angiotensin-converting enzyme against Rice stripe virus infection in a vector insect. Virology 2019, 533, 137–144. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Zhao, W.; Li, J.; Luo, L.; Kang, L.; Cui, F. The c-Jun N-terminal kinase pathway of a vector insect is activated by virus capsid protein and promotes viral replication. eLife 2017, 6, e26591. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, Y.; Wu, J.X.; Fu, S.; Li, C.Y.; Zhu, Z.R.; Zhou, X.P. Rice Stripe Tenuivirus Nonstructural Protein 3 Hijacks the 26S Proteasome of the Small Brown Planthopper via Direct Interaction with Regulatory Particle Non-ATPase Subunit 3. J. Virol. 2015, 89, 4296–4310. [Google Scholar] [CrossRef] [Green Version]
- Chen, X.F.; Yu, J.T.; Wang, W.; Lu, H.; Kang, L.; Cui, F. A Plant Virus Ensures Viral Stability in the Hemolymph of Vector Insects through Suppressing Prophenoloxidase Activation. mBio 2020, 11, e01453-20. [Google Scholar] [CrossRef]
- Li, S.; Wang, S.J.; Wang, X.; Li, X.L.; Zi, J.Y.; Ge, S.S.; Cheng, Z.B.; Zhou, T.; Ji, Y.H.; Deng, J.H.; et al. Rice stripe virus affects the viability of its vector offspring by changing developmental gene expression in embryos. Sci. Rep. 2015, 5, 7883. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wan, G.J.; Jiang, S.L.; Wang, W.J.; Li, G.Q.; Tao, X.R.; Pan, W.D.; Sword, G.A.; Chen, F.J. Rice stripe virus counters reduced fecundity in its insect vector by modifying insect physiology, primary endosymbionts and feeding behavior. Sci. Rep. 2015, 5, 12527. [Google Scholar] [CrossRef] [Green Version]
- Li, S.; Zhou, C.W.; Zhou, Y.J. Olfactory co-receptor Orco stimulated by Rice stripe virus is essential for host seeking behavior in small brown planthopper. Pest. Man. Sci. 2019, 75, 187–194. [Google Scholar] [CrossRef] [Green Version]
- Han, K.L.; Huang, H.J.; Zheng, H.Y.; Ji, M.F.; Yuan, Q.; Cui, W.J.; Zhang, H.H.; Peng, J.J.; Lu, Y.W.; Rao, S.F.; et al. Rice stripe virus coat protein induces the accumulation of jasmonic acid, activating plant defence against the virus while also attracting its vector to feed. Mol. Plant. Pathol. 2020, 21, 1647–1653. [Google Scholar] [CrossRef]
- Skoracka, A.; Smith, L.; Oldfield, G.; Cristofaro, M.; Amrine, J.W. Host-plant specificity and specialization in eriophyoid mites and their importance for the use of eriophyoid mites as biocontrol agents of weeds. In Eriophyoid Mites: Progress and Prognoses; Springer: Dordrecht, The Netherlands, 2009; pp. 93–113. [Google Scholar]
- Kulkarni, N.K.; Kumar, P.L.; Muniyappa, V.; Jones, A.T.; Reddy, D.V.R. Transmission of Pigeon pea sterility mosaic virus by the eriophyid mite, Aceria cajani (Acari: Arthropoda). Plant Dis. 2002, 86, 1297–1302. [Google Scholar] [CrossRef] [Green Version]
- Patil, B.L.; Kumar, P.L. Pigeonpea sterility mosaic emaraviruses: A journey from a mysterious virus to classic emaraviruses. In A Century of Plant Virology in India; Springer: Singapore, 2017; pp. 255–270. [Google Scholar]
- Mahmood, T.; Hein, G.L.; Jensen, S.G. Mixed infection of hard red winter wheat with High Plains virus and wheat streak mosaic virus from wheat curl mites in Nebraska. Plant Dis. 1998, 82, 311–315. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Druciarek, T.; Lewandowski, M.; Stainton, D.; Tzanetakis, I.E. Mite–be more interesting than you think: The story of rose rosette emaravirus vector. In Proceedings of the Entomological Society of America Meeting, Orlando, FL, USA, 15–18 November 2020. [Google Scholar]
- Amrine, J.W., Jr.; Hindal, D.F.; Stasny, T.A.; Williams, R.L.; Coffman, C.C. Transmission of the rose rosette disease agent to Rosa multiflora by Phyllocoptes fructiphylus (Acari: Eriophyidae). Entomol. News 1988, 99, 239–252. [Google Scholar]
- Reddy, M.V.; Nene, Y.L. Influence of sterility mosaic resistant pigeonpeas on multiplication of the mite vector. Ind. Phytopathol. 1980, 33, 61–63. [Google Scholar]
- Nault, L.R.; Styer, W.E. Transmission of an Eriophyid-borne wheat pathogen by Aceria tulipae. Phytopathology 1970, 60, 1616–1618. [Google Scholar] [CrossRef]
- Mielke-Ehret, N.; Thoma, J.; Schlatermund, N.; Mühlbach, H.P. Detection of European mountain ash ringspot-associated virus-specific RNA and protein P3 in the pear leaf blister mite Phytoptus pyri (Eriophyidae). Arch. Virol. 2010, 155, 987–991. [Google Scholar] [CrossRef] [PubMed]
- Butković, A.; González, R.; Elena, S.F. Revisiting Orthotospovirus phylogeny using full-genome data and testing the contribution of selection, recombination and segment reassortment in the origin of members of new species. Arch. Virol. 2021, 166, 491–499. [Google Scholar] [CrossRef] [PubMed]
- Ballinger, M.J.; Bruenn, J.A.; Hay, J.; Czechowski, D.; Taylor, D.J. Discovery and evolution of bunyavirids in arctic phantom midges and ancient bunyavirid-like sequences in insect genomes. J. Virol. 2014, 88, 8783–8794. [Google Scholar] [CrossRef] [Green Version]
- Marklewitz, M.; Zirkel, F.; Kurth, A.; Drosten, C.; Junglen, S. Evolutionary and phenotypic analysis of live virus isolates suggests arthropod origin of a pathogenic RNA virus family. Proc. Nat. Acad. Sci. USA 2015, 112, 7536–7541. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Navarro, B.; Minutolo, M.; De Stradis, A.; Palmisano, F.; Alioto, D.; Di Serio, F. The first phlebo-like virus infecting plants: A case study on the adaptation of negative-stranded RNA viruses to new hosts. Mol. Plant. Pathol. 2018, 19, 1075–1089. [Google Scholar] [CrossRef] [PubMed]
- Mushegian, A.R.; Elena, S.F. Evolution of plant virus movement proteins from the 30K superfamily and of their homologs integrated in plant genomes. Virology 2015, 476, 304–315. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Feng, M.; Cheng, R.; Chen, M.; Guo, R.; Li, L.; Feng, Z.; Wu, J.; Xie, L.; Hong, J.; Zhang, Z.; et al. Rescue of TSWV entirely from cDNA clones; establishment of the first reverse genetics system for a segmented (-)RNA plant virus. Proc. Natl. Acad. Sci. USA 2019. [Google Scholar] [CrossRef]
- Qiu, W.; Moyer, J.W. Tomato spotted wilt tospovirus adapts to the TSWV N gene-derived resistance by genome reassortment. Phytopathology 1999, 89, 575–582. [Google Scholar] [CrossRef]
- Webster, C.G.; Reitz, S.R.; Perry, K.L.; Adkins, S. A natural M RNA reassortant arising from two species of plant-and insect-infecting bunyaviruses and comparison of its sequence and biological properties to parental species. Virology 2011, 413, 216–225. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, L.Z.; Rao, L.X.; Zhou, X.P.; Wu, J.X. Genetic variability and evolution of rice stripe virus. J. Zhejiang Univ. Sci. B 2013, 14, 875–885. [Google Scholar] [CrossRef] [Green Version]
- Stewart, L.R. Sequence diversity of wheat mosaic virus isolates. Virus Res. 2016, 213, 299–303. [Google Scholar] [CrossRef] [Green Version]
- Hassan, M.; Tzanetakis, I.E. Population structure, evolution and detection of blackberry leaf mottle-associated virus, an emerging emaravirus. Plant. Path. 2019, 68, 775–782. [Google Scholar] [CrossRef]
- Patil, B.L.; Dangwal, M.; Mishra, R. Variability of emaravirus species associated with sterility mosaic disease of pigeonpea in India provides evidence of segment reassortment. Viruses 2017, 9, 183. [Google Scholar] [CrossRef] [Green Version]
- Bekal, S.; Domier, L.L.; Niblack, T.L.; Lambert, K.N. Discovery and initial analysis of novel viral genomes in the soybean cyst nematode. J. Gen. Virol. 2011, 92, 1870–1879. [Google Scholar] [CrossRef] [PubMed]
- Gaafar, Y.Z.A.; Richert-Pöggeler, K.R.; Sieg-Müller, A.; Lüddecke, P.; Herz, K.; Hartrick, J.; Seide, Y.; Vetten, H.J.; Ziebell, H. A divergent strain of melon chlorotic spot virus isolated from black medic (Medicago lupulina) in Austria. Virol. J. 2019, 16, 89. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, Y.H.; Fujita, M.; Chiba, S.; Hyodo, K.; Andika, I.B.; Suzuki, N.; Kondo, H. Two novel fungal negative-strand RNA viruses related to mymonaviruses and phenuiviruses in the shiitake mushroom (Lentinula edodes). Virology 2019, 533, 125–136. [Google Scholar] [CrossRef] [PubMed]
- Patterson, E.I.; Villinger, J.; Muthoni, J.N.; Dobel-Ober, L.; Hughes, G.L. Exploiting insect-specific viruses as a novel strategy to control vector-borne disease. Curr. Opin. Insect Sci. 2020, 39, 50–56. [Google Scholar] [CrossRef] [PubMed]
- Neriya, Y.; Morikawa, T.; Hamamoto, K.; Noguchi, K.; Kobayashi, T.; Suzuki, T.; Nishigawa, H.; Natsuaki, T. Characterization of tulip streak virus, a novel virus associated with the family Phenuiviridae. J. Gen. Virol. 2021, 102. [Google Scholar] [CrossRef] [PubMed]
- Kubota, K.; Yanagisawa, H.; Chiaki, Y.; Yamasaki, J.; Horikawa, H.; Tsuekawa, K.; Morita, Y.; Kadono, F. Complete nucleotide sequence of chrysanthemum mosaic-associated virus, a novel emaravirus infecting chrysanthemum. Arch. Virol. 2021, 166, 1241–1245. [Google Scholar] [CrossRef]
- Navarro, B.; Zicca, S.; Minutolo, M.; Saponari, M.; Alioto, D.; Di Serio, F. A Negative-Stranded RNA Virus Infecting Citrus Trees: The Second Member of a New Genus Within the Order Bunyavirales. Front. Microbiol. 2018, 9, 2340. [Google Scholar] [CrossRef]
- Tang, L.; Song, L.; Ye, Z.; Lin, C.; Wang, B.; Lin, J.; Gao, C.; Wang, A. A novel negative-stranded RNA virus of the order Bunyavirales identified in Brassica campestris L. ssp. chinensis. Arch. Virol. 2021. [Google Scholar] [CrossRef]
- Diaz-Lara, A.; Navarro, B.; Di Serio, F.; Stevens, K.; Hwang, M.S.; Kohl, J.; Vu, S.T.; Falk, B.W.; Golino, D.; Al Rwahnih, M. Two novel negative-sense RNA viruses infecting grapevine are members of a newly proposed genus within the family Phenuiviridae. Viruses 2019, 11, 685. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, X.; Sun, K.; Liang, Y.; Wang, S.; Wu, K.; Li, Z. Development of Rice Stripe Tenuivirus Minireplicon Reverse Genetics Systems Suitable for Analyses of Viral Replication and Intercellular Movement. Front. Microbiol. 2021, 12, 655256. [Google Scholar] [CrossRef] [PubMed]
- Feng, M.; Li, L.; Cheng, R.; Yuan, Y.; Dong, Y.; Chen, M.; Guo, R.; Yao, M.; Xu, Y.; Zhou, Y.; et al. Development of a mini-replicon-based reverse-genetics system for rice stripe tenuivirus. J. Virol. 2021. [Google Scholar] [CrossRef]
- de Ronde, D.; Lohuis, D.; Kormelink, R. Identification and characterisation of a new class of temperature-dependent Tsw-based Tomato spotted wilt virus resistance breaking isolates. Plant Pathol. 2019, 68, 60–71. [Google Scholar] [CrossRef] [Green Version]
- Prins, M.; Laimer, M.; Noris, E.; Schubert, J.; Wassenegger, M.; Tepfer, M. Strategies for antiviral resistance in transgenic plants. Mol. Plant. Path. 2008, 9, 73–83. [Google Scholar] [CrossRef] [PubMed]
- Sasaya, T.; Nakazono-Nagaoka, E.; Saika, H.; Aoki, H.; Hiraguri, A.; Netsu, O.; Uehara-Ichiki, T.; Onuki, M.; Toki, S.; Saito, K.; et al. Transgenic strategies to confer resistance against viruses in rice plants. Front. Microbiol. 2014, 4, 409. [Google Scholar] [CrossRef]
- Mitter, N.; Worrall, E.A.; Robinson, K.E.; Li, P.; Jain, R.G.; Taoch, Y.C.; Fletcher, S.J.; Carroll, B.J.; Lu, G.Q.; Xu, Z.P. Clay nanosheets for topical delivery of RNAi for sustained protection against plant viruses. Nat. Plants 2017, 3, 16207. [Google Scholar] [CrossRef]
- Tabein, S.; Jansen, M.; Noris, E.; Vaira, A.M.; Marian, D.; Behjatnia, S.A.A.; Accotto, G.P.; Miozzi, L. The Induction of an Effective dsRNA-Mediated Resistance Against Tomato Spotted Wilt Virus by Exogenous Application of Double-Stranded RNA Largely Depends on the Selection of the Viral RNA Target Region. Front. Plant. Sci. 2020, 11, 533338. [Google Scholar] [CrossRef]
- Bonning, B.C. The Insect Virome: Opportunities and Challenges. Curr. Issues Mol. Biol. 2020, 34, 1–12. [Google Scholar] [CrossRef]
- Li, C.X.; Shi, M.; Tian, J.H.; Lin, X.D.; Kang, Y.J.; Chen, L.J.; Qin, X.C.; Xu, J.; Holmes, E.C.; Zhang, Y.Z. Unprecedented genomic diversity of RNA viruses in arthropods reveals the ancestry of negative-sense RNA viruses. eLife 2015, 4, e05378. [Google Scholar] [CrossRef] [PubMed]
- François, S.; Mutuel, D.; Duncan, A.B.; Rodrigues, L.R.; Danzelle, C.; Lefevre, S.; Santos, I.; Frayssinet, M.; Fernandez, E.; Filloux, D.; et al. New Prevalent Densovirus Discovered in Acari. Insight from Metagenomics in Viral Communities Associated with Two-Spotted Mite (Tetranychus urticae) Populations. Viruses 2019, 11, 233. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kondo, H.; Fujita, M.; Hisano, H.; Hyodo, K.; Andika, I.B.; Suzuki, N. Virome Analysis of Aphid Populations That Infest the Barley Field: The Discovery of Two Novel Groups of Nege/Kita-Like Viruses and Other Novel RNA Viruses. Front. Microbiol. 2020, 11, 509. [Google Scholar] [CrossRef] [PubMed]
- Thekke-Veetil, T.; Lagos-Kutz, D.; McCoppin, N.K.; Hartman, G.L.; Ju, H.K.; Lim, H.S.; Domier, L.L. Soybean Thrips (Thysanoptera: Thripidae) Harbor Highly Diverse Populations of Arthropod, Fungal and Plant Viruses. Viruses 2020, 12, 1376. [Google Scholar] [CrossRef] [PubMed]
- Dolja, V.V.; Koonin, E.V. Metagenomics reshapes the concepts of RNA virus evolution by revealing extensive horizontal virus transfer. Virus Res. 2018, 244, 36–52. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Negarnaviricota- Subphylum | Class | Order | Family | Genus Containing Plant-Infecting Members | Type Species | Natural Mode of Transmission **** |
|---|---|---|---|---|---|---|
| Haploviricotina | Monjiviricetes | Mononegavirales (11 families, 71 genera, 339 species) | Rhabdoviridae (6/30 genera contain plant-infecting viruses) | Alphanucleorhabdovirus (1) | Potato yellow dwarf virus | Arthropods |
| Betanucleorhabdovirus (1) | Sonchus yellow net virus | Arthropods | ||||
| Gammanucleorhabdovirus (1) | Maize fine streak virus | Arthropods | ||||
| Cytorhabdovirus (1) | Lettuce necrotic yellows virus | Arthropods | ||||
| Dichoravirus (2) * | Orchid fleck virus | Arthropods | ||||
| Varicosavirus (2) * | Lettuce big-vein associated virus | Plasmodiophorid protists *** | ||||
| Milneviricetes | Serpentovirales | Aspiviridae (formerly Ophioviridae) | Ophiovirus (4) | Citrus psorosis virus (CPsV) | Plasmodiophorid protists | |
| Polyploviricotina | Ellioviricetes | Bunyavirales (12 families, 45 genera and two unassigned genera) | Phenuiviridae (3/19 genera contain plant-infecting viruses) | Tenuivirus (4-8) | Rice stripe virus (8) | Arthropods (14) |
| Coguvirus (2) ** | Citrus concave gum-associated virus (2) | ND (Grafting) **** | ||||
| Rubodvirus (3) ** | Apple rubbery wood virus (2) | ND (Grafting) **** | ||||
| Tospoviridae | Orthotospovirus (3) | Tomato spotted wilt virus (26) | Arthropods (15) | |||
| Fimoviridae | Emaravirus (5-10) | European Mountain Ash ringspot associated virus (11) | Mites (6) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kormelink, R.; Verchot, J.; Tao, X.; Desbiez, C. The Bunyavirales: The Plant-Infecting Counterparts. Viruses 2021, 13, 842. https://doi.org/10.3390/v13050842
Kormelink R, Verchot J, Tao X, Desbiez C. The Bunyavirales: The Plant-Infecting Counterparts. Viruses. 2021; 13(5):842. https://doi.org/10.3390/v13050842
Chicago/Turabian StyleKormelink, Richard, Jeanmarie Verchot, Xiaorong Tao, and Cecile Desbiez. 2021. "The Bunyavirales: The Plant-Infecting Counterparts" Viruses 13, no. 5: 842. https://doi.org/10.3390/v13050842