
Why Did ZIKV Perinatal Outcomes Differ in Distinct Regions of Brazil? An Exploratory Study of Two Cohorts

 , , , ,
, , , ,  ,
,  , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Clinical Samples
2.2. Adverse Outcomes
2.3. Laboratory Diagnosis of ZIKV and DENV Infection
2.3.1. DENV Virus Stock
2.3.2. Plaque Reduction Neutralization Tests
2.3.3. DENV IgG ELISA
2.3.4. Avidity Assay
2.4. Attack Rates
2.5. Statistical Analyses
3. Results
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- MS. Boletim Epidemiológico Secretaria de Vigilância em Saúde. Vol 46. n° 42, 2015a. Available online: http://portalarquivos.saude.gov.br/images/pdf/2015/dezembro/11/svs-be-2015-047-dengue-se47-final.pdf (accessed on 7 October 2020). (In Portugese)
- Microcefalias. Centro de Operações de Emergências em Saúde Pública sobre Microcefalias.–Informe epidemiológico n° 02/2015-Semana Epidemiológica 47 de 2015. Brasil, Ministério da Saúde. Available online:http://portalsaude.saude.gov.br/images/pdf/2015/novembro/30/COES-Microcefalias---Informe-Epidemiologico---SE-47---30nov2015.pdf (accessed on 7 October 2020). (In Portugese)
- França, G.V.; Schuler-Faccini, L.; Oliveira, W.K.; Henriques, C.M.; Carmo, E.H.; Pedi, V.D.; Nunes, M.L.; Castro, M.C.; Serruya, S.; Silveira, M.F.; et al. Congenital Zika virus syndrome in Brazil: A case series of the first 1501 livebirths with complete investigation. Lancet 2016, 388, 891–897. [Google Scholar] [CrossRef] [Green Version]
- Brito, C. Zika Virus: A New Chapter in the History of Medicine. Acta Med. Port. 2015, 28, 679–680. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Albuquerque, M.F.P.M.; Souza, W.V.; Araújo, T.V.B.; Braga, M.C.; Miranda Filho, D.B.; Ximenes, R.A.A.; de Melo Filho, D.A.; Brito, C.A.A.; Valongueiro, S.; Melo, A.P.L.; et al. The microcephaly epidemic and Zika virus: Building knowledge in epidemiology. Cad. Saude Publica 2018, 34, e00069018. [Google Scholar] [PubMed]
- Brasil, P.; Calvet, G.A.; Siqueira, A.M.; Wakimoto, M.; de Sequeira, P.C.; Nobre, A.; Quintana Mde, S.; Mendonça, M.C.; Lupi, O.; de Souza, R.V.; et al. Zika Virus Outbreak in Rio de Janeiro, Brazil: Clinical Characterization, Epidemiological and Virological Aspects. PLoS Negl. Trop. Dis. 2016, 10, e0004636. [Google Scholar] [CrossRef]
- OMS. Epidemiological Alert-Zika Virus Infection. Available online: https://www.paho.org/hq/dmdocuments/2015/2015-dec-1-cha-epi-alert-zika-neuro-syndrom (accessed on 9 November 2020).
- OMS. Epidemiological Alert-Increase of Microcephaly in the Northeast of Brazil. Available online: https://www.paho.org/hq/dmdocuments/2015/2015-nov-17-cha-microcephaly-epi-alert.pdf (accessed on 9 November 2020).
- Gulland, A. Zika virus is a global public health emergency, declares WHO. BMJ 2016, 352, i657. [Google Scholar] [CrossRef] [Green Version]
- Rodriguez-Barraquer, I.; Costa, F.; Nascimento, E.J.M.; Nery, N.; Castanha, P.M.S.; Sacramento, G.A.; Cruz, J.; Carvalho, M.; De Olivera, D.; Hagan, J.E.; et al. Impact of preexisting dengue immunity on Zika virus emergence in a dengue endemic region. Science 2019, 363, 607–610. [Google Scholar] [CrossRef] [Green Version]
- MS, Monitoramento dos casos de Dengue, Febre de Chikungunya e Febre Pelo Vírus Zika até a Semana Epidemiológica 52. 2016. Available online:https://www.saude.gov.br/images/pdf/2017/abril/06/2017-002-Monitoramento-dos-casos-de-dengue--febre-de-chikungunya-e-febre-pelo-v--rus-Zika-ate-a-Semana-Epidemiologica-52--2016.pdf (accessed on 22 February 2020).
- MS. Ministério da Saúde Confirma 1.709 Casos de Microcefalia. Available online: https://www.saude.gov.br//saude/pt-br/assuntos/noticias/ministerio-da-saude-confirma-1-709-casos-de-microcefalia (accessed on 22 February 2021).
- Heinz, F.X.; Stiasny, K. The Antigenic Structure of Zika Virus and Its Relation to Other Flaviviruses: Implications for Infection and Immunoprophylaxis. Microbiol. Mol. Biol. Rev. 2017, 81, e00055-16. [Google Scholar] [CrossRef] [Green Version]
- Priyamvada, L.; Hudson, W.; Ahmed, R.; Wrammert, J. Humoral cross-reactivity between Zika and dengue viruses: Implications for protection and pathology. Emerg. Microbes Infect. 2017, 6, e33. [Google Scholar] [CrossRef] [Green Version]
- Bardina, S.V.; Bunduc, P.; Tripathi, S.; Duehr, J.; Frere, J.J.; Brown, J.A.; Nachbagauer, R.; Foster, G.A.; Krysztof, D.; Tortorella, D.; et al. Enhancement of Zika virus pathogenesis by preexisting antiflavivirus immunity. Science 2017, 356, 175–180. [Google Scholar] [CrossRef] [Green Version]
- Collins, M.H.; McGowan, E.; Jadi, R.; Young, E.; Lopez, C.A.; Baric, R.S.; Lazear, H.M.; de Silva, A.M. Lack of Durable Cross-Neutralizing Antibodies Against Zika Virus from Dengue Virus Infection. Emerg. Infect. Dis. 2017, 23, 773–781. [Google Scholar] [CrossRef]
- Montoya, M.; Collins, M.; Dejnirattisai, W.; Katzelnick, L.C.; Puerta-Guardo, H.; Jadi, R.; Schildhauer, S.; Supasa, P.; Vasanawathana, S.; Malasit, P.; et al. Longitudinal Analysis of Antibody Cross-neutralization Following Zika Virus and Dengue Virus Infection in Asia and the Americas. J. Infect. Dis. 2018, 218, 536–545. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carvalho, M.S.; Freitas, L.P.; Cruz, O.G.; Brasil, P.; Bastos, L.S. Association of past dengue fever epidemics with the risk of Zika microcephaly at the population level in Brazil. Sci. Rep. 2020, 10, 1752. [Google Scholar] [CrossRef] [Green Version]
- Brady, O.J.; Osgood-Zimmerman, A.; Kassebaum, N.J.; Ray, S.E.; de Araújo, V.E.M.; da Nóbrega, A.A.; Frutuoso, L.C.V.; Lecca, R.C.R.; Stevens, A.; Zoca de Oliveira, B.; et al. The association between Zika virus infection and microcephaly in Brazil 2015-2017: An observational analysis of over 4 million births. PLoS Med. 2019, 16, e1002755. [Google Scholar] [CrossRef] [PubMed]
- Katzelnick, L.C.; Gresh, L.; Halloran, M.E.; Mercado, J.C.; Kuan, G.; Gordon, A.; Balmaseda, A.; Harris, E. Antibody-dependent enhancement of severe dengue disease in humans. Science 2017, 358, 929–932. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Regla-Nava, J.A.; Elong Ngono, A.; Viramontes, K.M.; Huynh, A.T.; Wang, Y.T.; Nguyen, A.T.; Salgado, R.; Mamidi, A.; Kim, K.; Diamond, M.S.; et al. Cross-reactive Dengue virus-specific CD8+ T cells protect against Zika virus during pregnancy. Nat. Commun. 2018, 9, 3042. [Google Scholar] [CrossRef] [Green Version]
- Moreira-Soto, A.; Cabral, R.; Pedroso, C.; Eschbach-Bludau, M.; Rockstroh, A.; Vargas, L.A.; Postigo-Hidalgo, I.; Luz, E.; Sampaio, G.S.; Drosten, C.; et al. Exhaustive TORCH Pathogen Diagnostics Corroborate Zika Virus Etiology of Congenital Malformations in Northeastern Brazil. mSphere 2018, 3, e00278-18. [Google Scholar] [CrossRef] [Green Version]
- Halstead, S.B. Biologic Evidence Required for Zika Disease Enhancement by Dengue Antibodies. Emerg. Infect. Dis. 2017, 23, 569–573. [Google Scholar] [CrossRef]
- Khandia, R.; Munjal, A.; Dhama, K.; Karthik, K.; Tiwari, R.; Malik, Y.S.; Singh, R.K.; Chaicumpa, W. Modulation of Dengue/Zika Virus Pathogenicity by Antibody-Dependent Enhancement and Strategies to Protect Against Enhancement in Zika Virus Infection. Front. Immunol. 2018, 9, 597. [Google Scholar] [CrossRef]
- Castanha, P.M.S.; Nascimento, E.J.M.; Braga, C.; Cordeiro, M.T.; de Carvalho, O.V.; de Mendonça, L.R.; Azevedo, E.A.N.; França, R.F.O.; Dhalia, R.; Marques, E.T.A. Dengue Virus-Specific Antibodies Enhance Brazilian Zika Virus Infection. J. Infect. Dis. 2017, 215, 781–785. [Google Scholar] [CrossRef] [Green Version]
- Brasil, P.; Pereira, J.P.; Moreira, M.E.; Ribeiro Nogueira, R.M.; Damasceno, L.; Wakimoto, M.; Rabello, R.S.; Valderramos, S.G.; Halai, U.A.; Salles, T.S.; et al. Zika Virus Infection in Pregnant Women in Rio de Janeiro. N. Engl. J. Med. 2016, 375, 2321–2334. [Google Scholar] [CrossRef]
- Nogueira, M.L.; Nery Júnior, N.R.R.; Estofolete, C.F.; Bernardes Terzian, A.C.; Guimarães, G.F.; Zini, N.; Alves da Silva, R.; Dutra Silva, G.C.; Junqueira Franco, L.C.; Rahal, P.; et al. Adverse birth outcomes associated with Zika virus exposure during pregnancy in São José do Rio Preto, Brazil. Clin. Microbiol. Infect. 2018, 24, 646–652. [Google Scholar] [CrossRef] [Green Version]
- Redivo, E.F.; Bôtto Menezes, C.; da Costa Castilho, M.; Brock, M.; da Silva Magno, E.; Gomes Saraiva, M.D.G.; Alvarez Fernandes, S.S.; Costa Antony de Andrade, A.B.; Costa Alecrim, M.D.G.; Martinez-Espinosa, F.E. Zika Virus Infection in a Cohort of Pregnant Women with Exanthematic Disease in Manaus, Brazilian Amazon. Viruses 2020, 12, 1362. [Google Scholar] [CrossRef] [PubMed]
- Villar, J.; Cheikh Ismail, L.; Victora, C.G.; Ohuma, E.O.; Bertino, E.; Altman, D.G.; Lambert, A.; Papageorghiou, A.T.; Carvalho, M.; Jaffer, Y.A.; et al. International Fetal and Newborn Growth Consortium for the 21st Century (INTERGROWTH-21st). International standards for newborn weight, length, and head circumference by gestational age and sex: The Newborn Cross-Sectional Study of the INTERGROWTH-21st Project. Lancet 2014, 384, 857–868. [Google Scholar] [PubMed]
- Moore, C.A.; Staples, J.E.; Dobyns, W.B.; Pessoa, A.; Ventura, C.V.; Fonseca, E.B.; Ribeiro, E.M.; Ventura, L.O.; Neto, N.N.; Arena, J.F.; et al. Characterizing the Pattern of Anomalies in Congenital Zika Syndrome for Pediatric Clinicians. JAMA Pediatr. 2017, 171, 288–295. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rasmussen, S.A.; Jamieson, D.J.; Honein, M.A.; Petersen, L.R. Zika Virus and Birth Defects--Reviewing the Evidence for Causality. N. Engl. J. Med. 2016, 374, 1981–1987. [Google Scholar] [CrossRef]
- Lanciotti, R.S.; Kosoy, O.L.; Laven, J.J.; Velez, J.O.; Lambert, A.J.; Johnson, A.J.; Stanfield, S.M.; Duffy, M.R. Genetic and serologic properties of Zika virus associated with an epidemic, Yap State, Micronesia, 2007. Emerg. Infect. Dis. 2008, 14, 1232–1239. [Google Scholar] [CrossRef]
- Figueiredo, L.T. Uso de células de Aedes albopictus C6/36 na propagação e classificação de arbovírus das famílias Togaviridae, Flaviviridae, Bunyaviridae e Rhabdoviridae [The use of Aedes albopictus C6/36 cells in the propagation and classification of arbovirus of the Togaviridae, Flaviviridae, Bunyaviridae and Rhabdoviridae families]. Rev. Soc. Bras. Med. Trop. 1990, 23, 13–18. [Google Scholar]
- Vasilakis, N.; Durbin, A.P.; da Rosa, A.P.; Munoz-Jordan, J.L.; Tesh, R.B.; Weaver, S.C. Antigenic relationships between sylvatic and endemic dengue viruses. Am. J. Trop. Med. Hyg. 2008, 79, 128–132. [Google Scholar] [CrossRef]
- Roehrig, J.T.; Hombach, J.; Barrett, A.D. Guidelines for Plaque-Reduction Neutralization Testing of Human Antibodies to Dengue Viruses. Viral Immunol. 2008, 21, 123–132. [Google Scholar] [CrossRef]
- Vasilakis, N.; Shell, E.J.; Fokam, E.B.; Mason, P.W.; Hanley, K.A.; Estes, D.M.; Weaver, S.C. Potential of ancestral sylvatic dengue-2 viruses to re-emerge. Virology 2007, 358, 402–412. [Google Scholar] [CrossRef] [Green Version]
- Polanec, J.; Seppälä, I.; Rousseau, S.; Hedman, K. Evaluation of protein-denaturing immunoassays for avidity of immunoglobulin G to rubella virus. J. Clin. Lab. Anal. 1994, 8, 16–21. [Google Scholar] [CrossRef]
- Breslow, N.E.; Clayton, D.G. Approximate Inference in Generalized Linear Mixed Models. J. Am. Stat. Assoc. 1993, 88, 9–25. [Google Scholar]
- Villela, D.A.M.; Bastos, L.S.; De Carvalho, L.M.; Cruz, O.G.; Gomes, M.F.C.; Durovni, B.; Lemos, M.C.; Saraceni, V.; Coelho, F.C.; Codeço, C.T. Zika in Rio de Janeiro: Assessment of basic reproduction number and comparison with dengue outbreaks. Epidemiol. Infect. 2017, 145, 1649–1657. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jaenisch, T.; Rosenberger, K.D.; Brito, C.; Brady, O.; Brasil, P.; Marques, E.T. Risk of microcephaly after Zika virus infection in Brazil, 2015 to 2016. Bull. World Health Organ. 2017, 95, 191–198. [Google Scholar] [CrossRef]
- Braga, J.U.; Bressan, C.; Dalvi, A.P.R.; Calvet, G.A.; Daumas, R.P.; Rodrigues, N.; Wakimoto, M.; Nogueira, R.M.R.; Nielsen-Saines, K.; Brito, C.; et al. Accuracy of Zika virus disease case definition during simultaneous Dengue and Chikungunya epidemics. PLoS ONE 2017, 12, e0179725. [Google Scholar] [CrossRef] [PubMed]
- Mansfield, K.L.; Horton, D.L.; Johnson, N.; Li, L.; Barrett, A.D.T.; Smith, D.J.; Galbraith, S.E.; Solomon, T.; Fooks, A.R. Flavivirus-induced antibody cross-reactivity. J. Gen. Virol. 2011, 92, 2821–2829. [Google Scholar] [CrossRef]
- De Góes Cavalcanti, L.P.; Tauil, P.L.; Alencar, C.H.; Oliveira, W.; Teixeira, M.M.; Heukelbach, J. Zika virus infection, associated microcephaly, and low yellow fever vaccination coverage in Brazil: Is there any causal link? J. Infect. Dev. Ctries. 2016, 10, 563–566. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nielsen-Saines, K.; Brasil, P.; Kerin, T.; Vasconcelos, Z.; Gabaglia, C.R.; Damasceno, L.; Pone, M.; Abreu de Carvalho, L.M.; Pone, S.M.; Zin, A.A.; et al. Delayed childhood neurodevelopment and neurosensory alterations in the second year of life in a prospective cohort of ZIKV-exposed children. Nat. Med. 2019, 25, 1213–1217. [Google Scholar] [CrossRef]
- McCracken, M.K.; Gromowski, G.D.; Friberg, H.L.; Lin, X.; Abbink, P.; De La Barrera, R.; Eckles, K.H.; Garver, L.S.; Boyd, M.; Jetton, D.; et al. Impact of prior flavivirus immunity on Zika virus infection in rhesus macaques. PLoS Pathog. 2017, 13, e1006487. [Google Scholar] [CrossRef]
- Pedroso, C.; Fischer, C.; Feldmann, M.; Sarno, M.; Luz, E.; Moreira-Soto, A.; Cabral, R.; Netto, E.M.; Brites, C.; Kümmerer, B.M.; et al. Cross-Protection of Dengue Virus Infection against Congenital Zika Syndrome, Northeastern Brazil. Emerg. Infect. Dis. 2019, 25, 1485–1493. [Google Scholar] [CrossRef] [Green Version]



{kind=link}
{kind=link}
| Municipality | p | |||
|---|---|---|---|---|
| Maternal Characteristics | Total | Rio de Janeiro, RJ (N = 50) | Manaus, AM (N = 64) | |
| Age | N(%) | N(%) | N(%) | 0.09 |
| <20 | 16 (14) | 5 (10) | 11 (17) | |
| 20–29 | 47 (41) | 16 (32) | 31 (48) | |
| 30–39 | 45 (39) | 26 (52) | 19 (30) | |
| >40 | 6 (5) | 3 (6) | 3 (5) | |
| Gestational age at the time of maternal ZIKV infection (weeks) | N(%) | N(%) | N(%) | 0.004 |
| <=26 | 75 (66) | 40 (80) | 35 (55) | |
| >=27 | 35 (31) | 7 (14) | 28 (44) | |
| Missing | 7 (6) | 3 (6) | 4 (6) | |
| DENV IgG | 0.05 | |||
| Reactive | 93 (81) | 45 (90) | 48 (75) | |
| Non-reactive | 21 (18) | 5 (10) | 16 (25) | |
| Adverse Outcomes | 31 (27) | 27 (54) | 4 (6) | <0.001 |
| Disproportionate Microcephaly and cerebral calcifications, irritability, hypertonicity, abnormal funduscopic and hearing exam, and clubfoot or arthrogryposis | 6 (19) | 5 (16) | 1 (3) | |
| Proportional microcephaly, hypertonicity, or abnormal neurologic exam | 2 (6) | 2 (6) | 0 | |
| Smallness for gestational age (SGA) and hypotonicity, or hearing deficits and hypertonicity; structural abnormality in neuroimaging | 4 (13) | 4 (13) | 0 | |
| Abnormal funduscopic exam and irritability, dysphagia, or hypertonicity and abnormal reflexes | 4 (13) | 4 (13) | 0 | |
| Hearing deficits, hypertonicity, and abnormal reflexes | 3 (10) | 3 (10) | 0 | |
| Hypertonicity and hyperreflexia; knee fovea, redundant scalp | 4 (13) | 4 (13) | 0 | |
| Dysphagia and irritability | 1(3) | 1 (3) | 0 | |
| Image structural abnormalities | 3 (10) | 3 (10) | 0 | |
| Fetal death/stillbirth | 4 (13) | 3 (10) | 1 (3) | |
| Normal findings | 83 (73) | 23 (46) | 60 (94) | |
| DENV Exposure | ||||
|---|---|---|---|---|
| Maternal Characteristics | Total | Prior DENV Exposure (N = 93) | No Prior DENV Exposure (N = 21) | p |
| Age | N(%) | N(%) | N(%) | 0.179 |
| <20 | 16 (14.04) | 10 (10.8) | 6 (28.6) | |
| 20–29 | 47 (41.23) | 41 (44.1) | 6 (28.6) | |
| 30–39 | 45 (39.47) | 37 (39.8) | 8 (38.1) | |
| >40 | 6 (5.26) | 5 (5.4) | 1 (4.8) | |
| Gestational age at ZIKV infection with (weeks) | N(%) | N(%) | N(%) | 0.603 |
| <=26 | 75 (65.79) | 62 (66.7) | 13 (61.9) | |
| >=27 | 35 (30.7) | 27 (29) | 8 (38.1) | |
| Missing | 8 (7.02) | 4 (4.3) | 4 (19) | |
| Municipality | 0.052 | |||
| Rio de Janeiro | 50 (43.86) | 45 (48.4) | 5 (23.8) | |
| Manaus | 64 (56.14) | 48 (51.6) | 16 (76.2) | |
| Infant Outcomes | 0.181 | |||
| Altered | 31 (27.2) | 28 (30.1) | 3 (14.2) | |
| Non-altered | 83 (70.18) | 65 (69.9) | 18 (85.8) | |
| Infant Outcomes | ||||
|---|---|---|---|---|
| Maternal Characteristics | Total | Normal Findings | Abnormal Findings | p |
| Age | N(%) | N(%) | N(%) | 0.34 |
| <20 | 16 (14) | 10 (16) | 6 (12) | |
| 20–29 | 44 (38) | 28 (44) | 16 (31) | |
| 30–39 | 48 (42) | 22 (35) | 26 (51) | |
| >40 | 6 (5) | 3 (5) | 3 (6) | |
| Gestational age at the time of maternal ZIKV infection(weeks) | N(%) | N(%) | N(%) | <0.001 |
| <=26 | 75 (66) | 53 (66) | 22 (65) | |
| >=27 | 35 (31) | 27 (34) | 8 (24) | |
| Missing | 4 (4) | 0 | 4 (12) | |
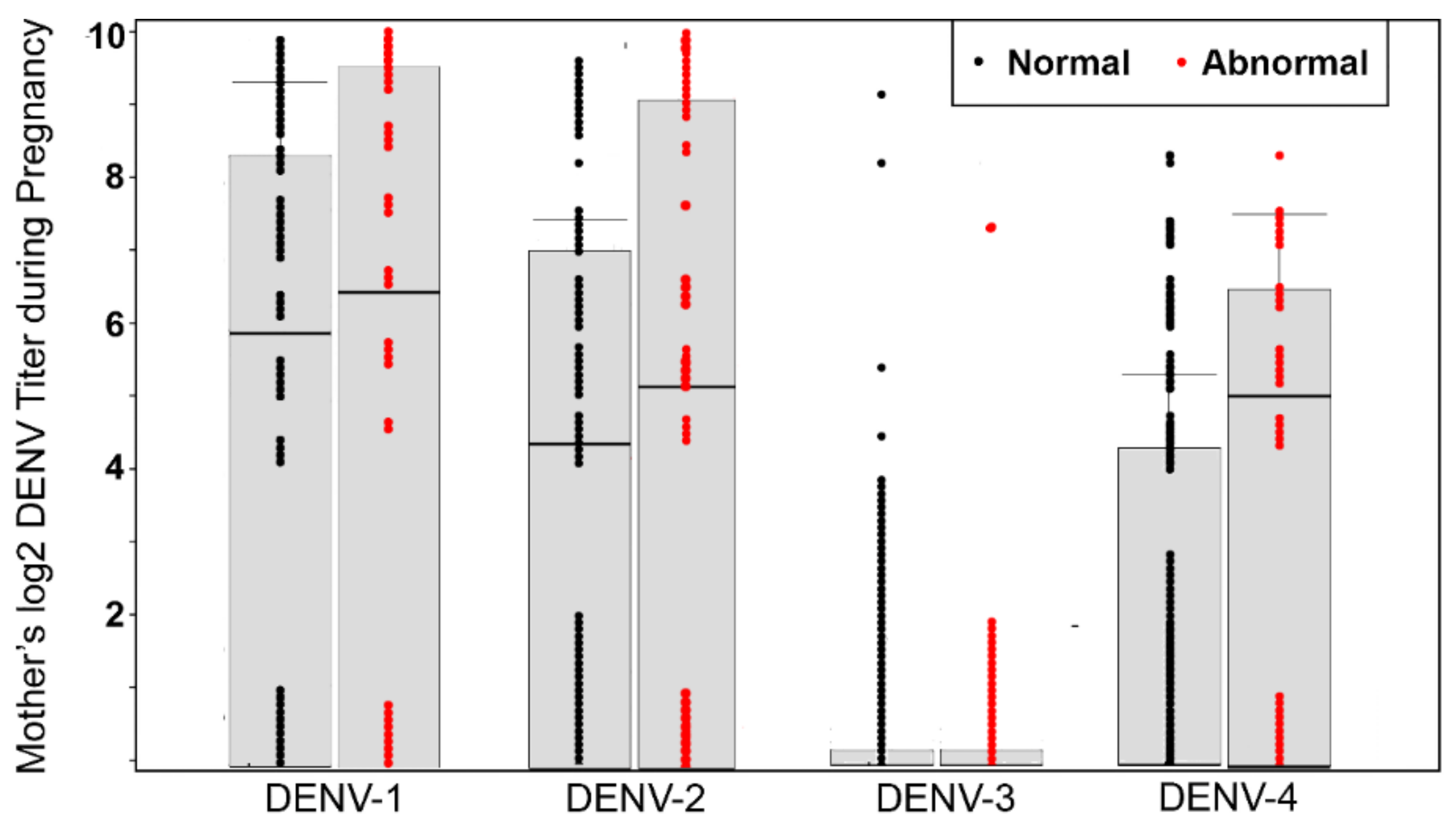
| Log2 DENV PRNT 80 (N = 114, median, IQR) | 0.474 | |||
| DENV-1 | 6.3 (0–8.1) | 5.9 (0–8.3) | 6.3 (0–9.3) | |
| DENV-2 | 4.4 (0–7.3) | 4.3 (0–7.1) | 5.3 (0–9.3) | |
| DENV-3 | 0 | 0 | 0 | |
| DENV-4 | 0 (0–5.4) | 0 (0–4.3) | 4.9 (0–6.3) | |
| Municipality | N (%) | N (%) | N (%) | <0.001 |
| Rio de Janeiro | 50 (43.9) | 23 (28.7) | 27 (79.4) | |
| Manaus | 64 (56.1) | 57 (71.3) | 7 (20.6) | |
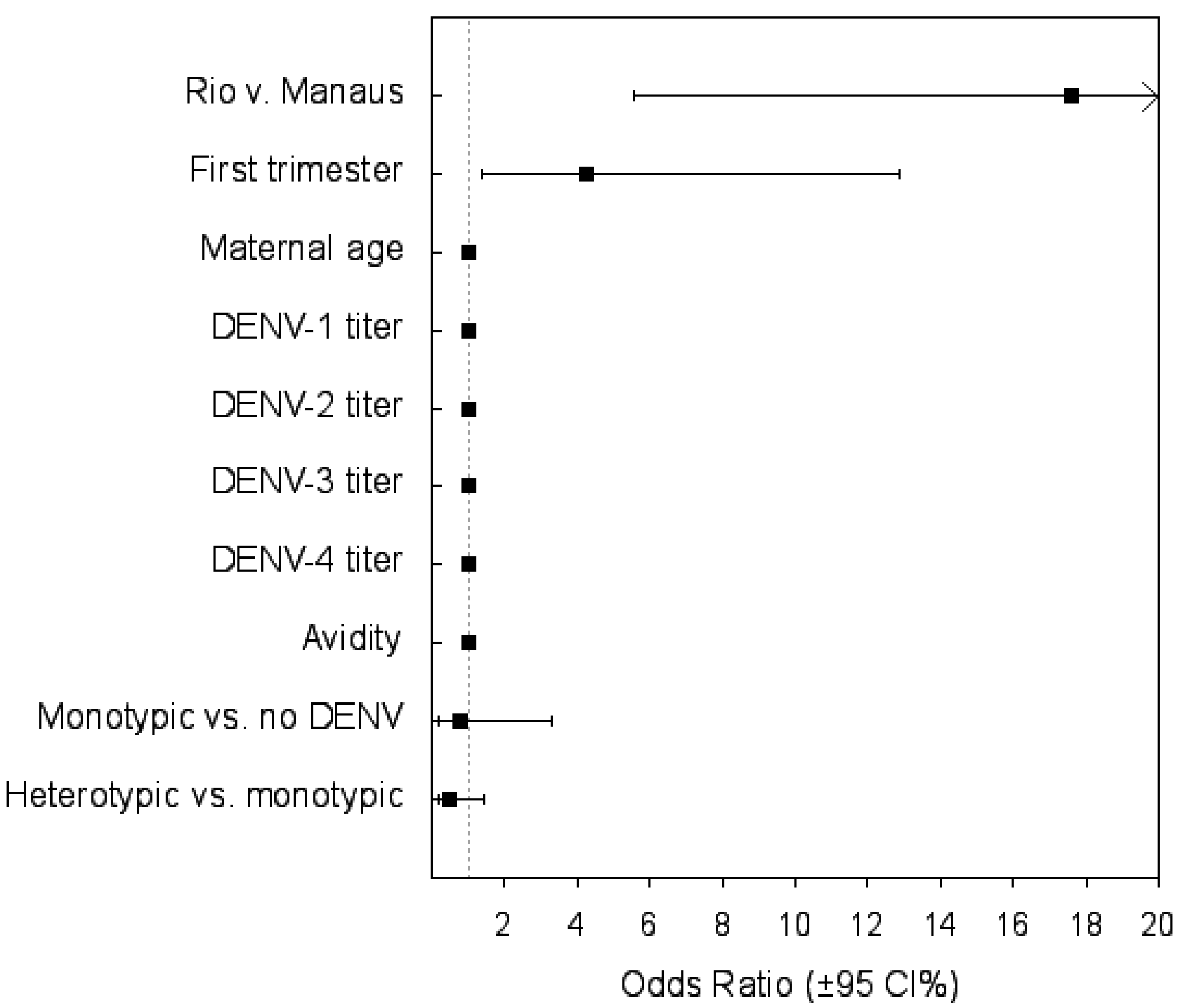
| Variables | Odds Ratio (95% CI) | p |
|---|---|---|
| Municipal-scale variable | ||
| Attack rate Rio vs. Manaus | 17.6 (5.55–55.88) | <0.001 |
| Maternal categorical variables | ||
| First trimester ZIKV infection | 4.26 (1.4–12.9) | 0.011 |
| Monotypic DENV infection vs. no DENV infection | 0.78 (0.18–3.31) | 0.73 |
| Heterotypic vs. monotypic DENV infection | 0.466 (0.154–1.413) | 0.18 |
| Maternal continuous variables | ||
| Maternal age | 1.0 (0.99–1) | 0.59 |
| DENV-1 titer | 0.99 (0.99–1) | 0.198 |
| DENV-2 titer | 0.99 (0.99–1) | 0.846 |
| DENV-3 titer | 0.99 (0.98–1.0) | 0.32 |
| DENV-4 titer | 1 (0.99–1.01) | 0.7 |
| Test of avidity | 0.99(0.96–1.02) | 0.523 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Damasceno, L.; Terzian, A.C.B.; Fuller, T.; Estofolete, C.F.; Andrade, A.; Kroon, E.G.; Zin, A.A.; Vasconcelos, Z.; Pereira, J.P., Jr.; Castilho, M.C.; et al. Why Did ZIKV Perinatal Outcomes Differ in Distinct Regions of Brazil? An Exploratory Study of Two Cohorts. Viruses 2021, 13, 736. https://doi.org/10.3390/v13050736
Damasceno L, Terzian ACB, Fuller T, Estofolete CF, Andrade A, Kroon EG, Zin AA, Vasconcelos Z, Pereira JP Jr., Castilho MC, et al. Why Did ZIKV Perinatal Outcomes Differ in Distinct Regions of Brazil? An Exploratory Study of Two Cohorts. Viruses. 2021; 13(5):736. https://doi.org/10.3390/v13050736
Chicago/Turabian StyleDamasceno, Luana, Ana Carolina B. Terzian, Trevon Fuller, Cassia F. Estofolete, Adriana Andrade, Erna G. Kroon, Andrea A. Zin, Zilton Vasconcelos, Jose P. Pereira, Jr., Márcia C. Castilho, and et al. 2021. "Why Did ZIKV Perinatal Outcomes Differ in Distinct Regions of Brazil? An Exploratory Study of Two Cohorts" Viruses 13, no. 5: 736. https://doi.org/10.3390/v13050736