
Virulence and Infectivity of UC, MD, and L Strains of Infectious Hematopoietic Necrosis Virus (IHNV) in Four Populations of Columbia River Basin Chinook Salmon


Abstract
:1. Introduction
2. Materials and Methods
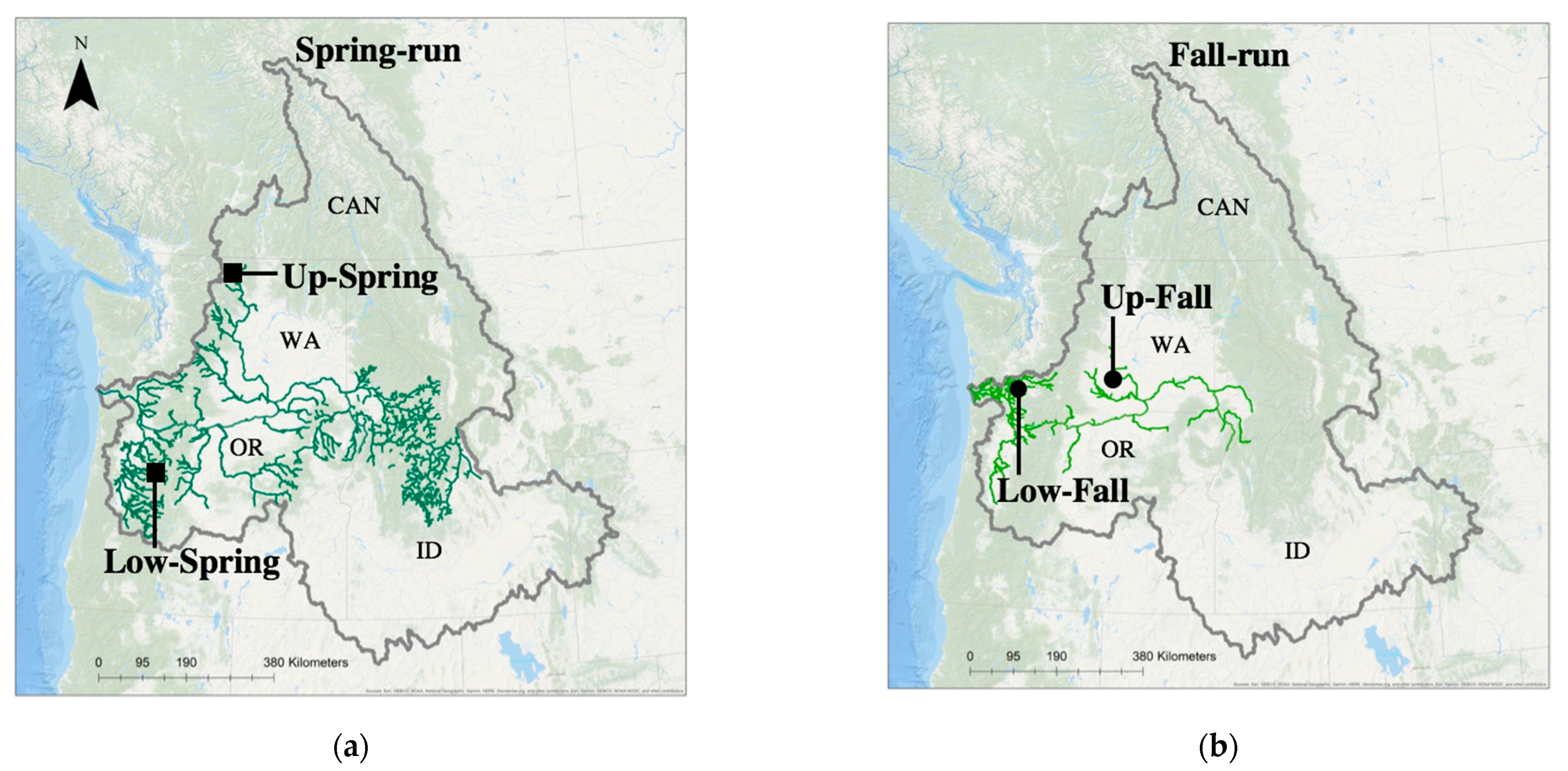
2.1. Experimental Challenges: Host Populations
2.2. Experimental Challenges: Virus Exposures
2.3. Measures of Viral Load
2.3.1. RNA Extraction and cDNA Synthesis
2.3.2. Viral RNA Quantification via Reverse Transcriptase Real-Time PCR
2.3.3. Statistical Analyses
Survival of Experimental Host Populations
Infection of Experimental Host Populations
The Influence of Exposure Dose, Viral Strain, and Host Population on Infection Frequency
- is the odds ratio associated with the ith fish, the expected number of infected individuals per uninfected individual;
- is the model’s y-intercept, capturing the reference level’s influence on the infection outcome;
- is the log10 of the exposure dose for the ith fish;
- is the effect size of exposure dose on the infection outcome;
- is the dummy variable for the hth host population for the ith fish;
- is the offset from the y-intercept associated with the hth host population;
- is the dummy variable for the sth viral strain for the ith fish;
- is the offset from the y-intercept associated with the sth viral strain.
2.3.4. The Influence of Exposure Dose, Viral Strain, and Host Population on Viral Load
2.4. Analysis of IHNV Field Surveillance Data
2.5. Abundance of Spring-, Summer- and Fall-Run Chinook Salmon across the CRB
3. Results
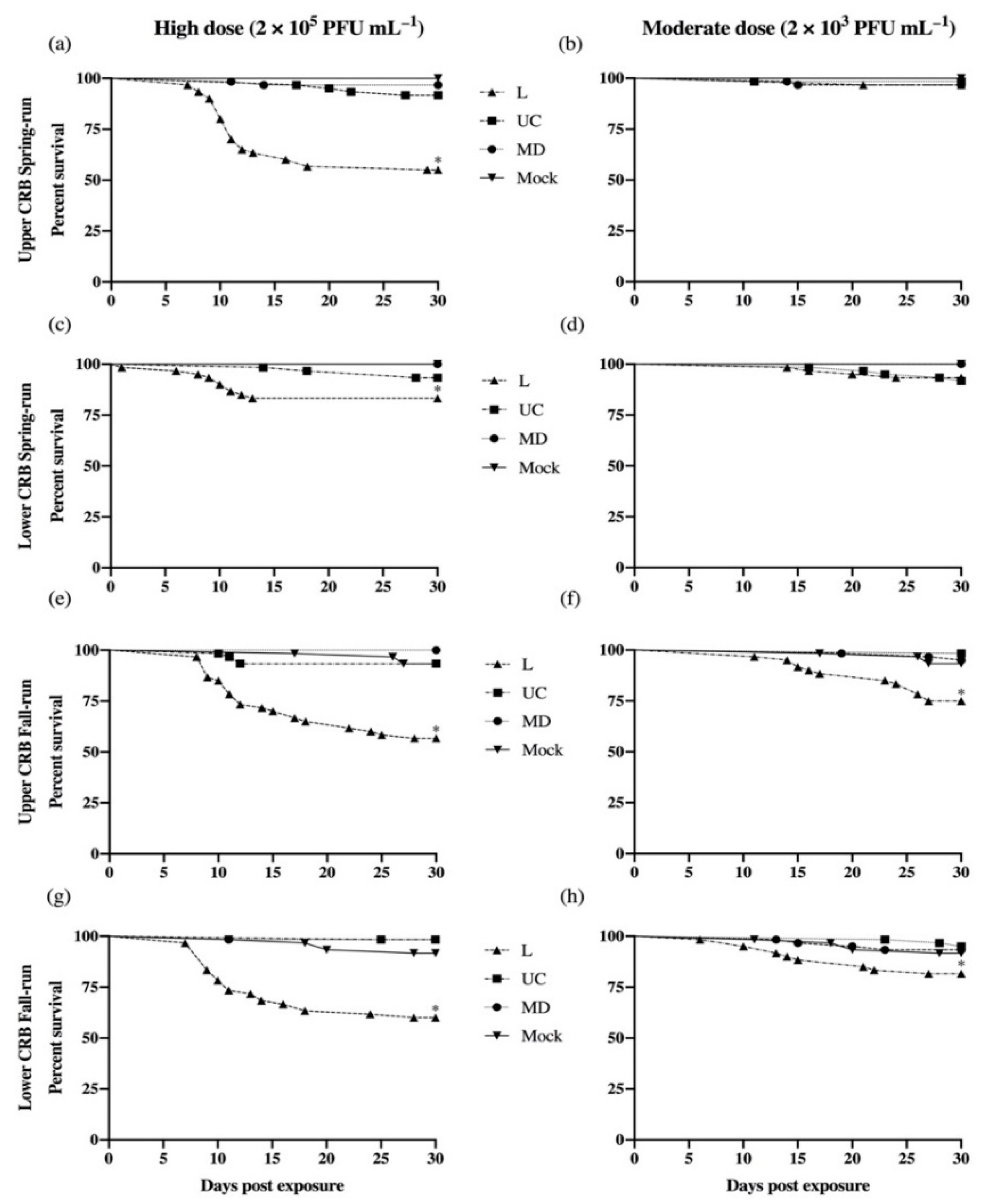
3.1. Survival of Experimental Populations of Columbia River Basin Chinook Salmon
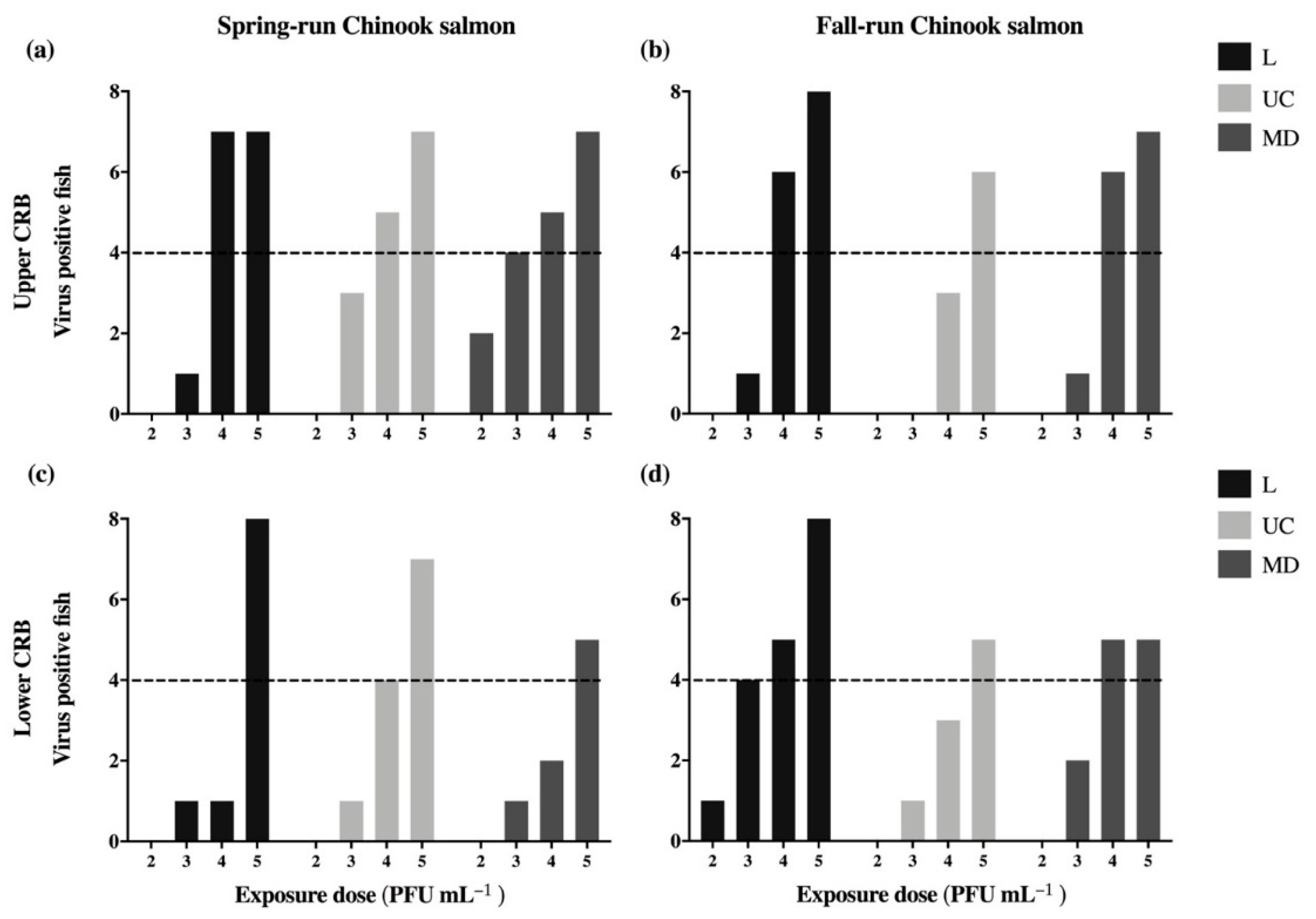
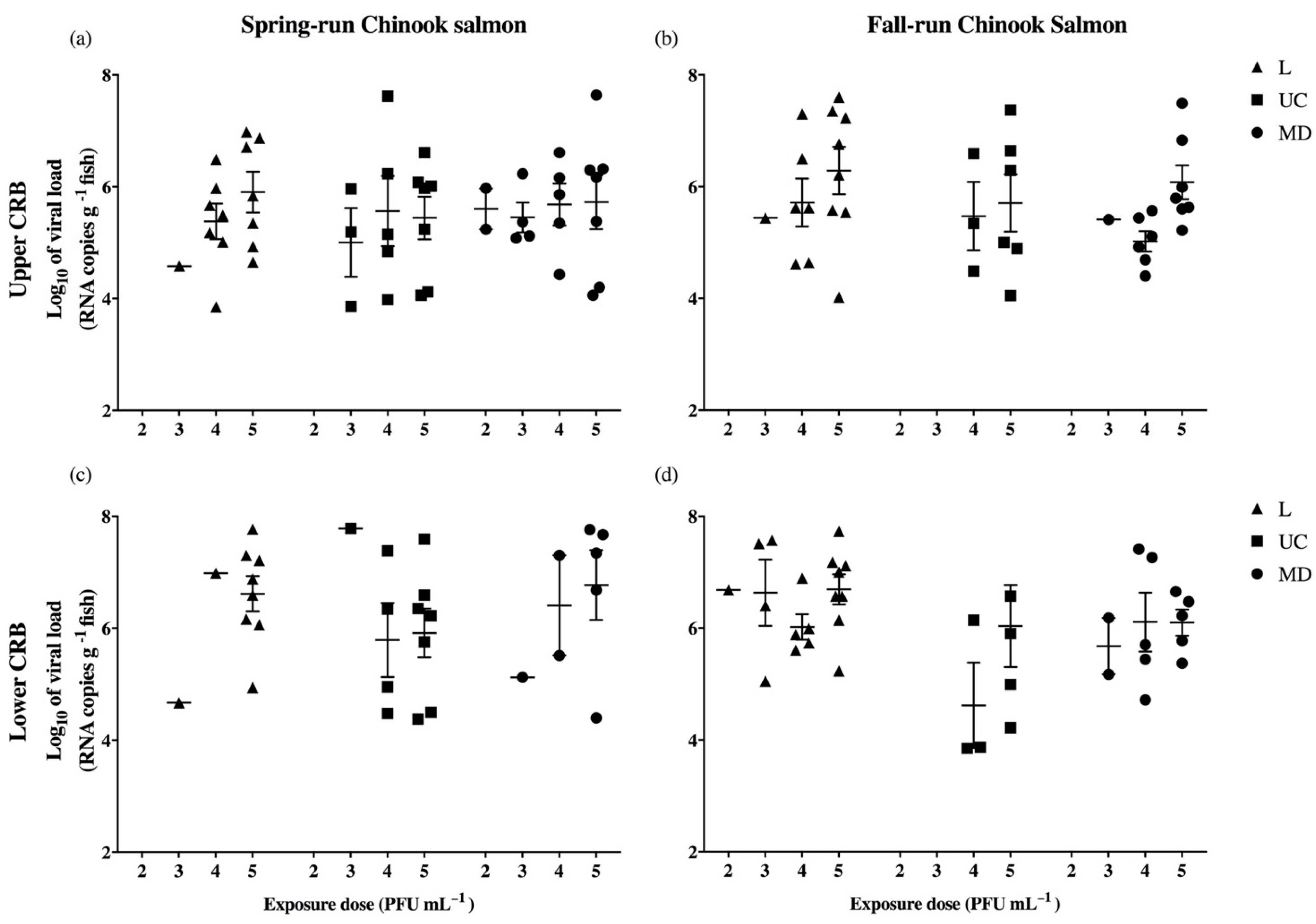
3.2. Infection of Experimental Populations of Columbia River Basin Chinook Salmon
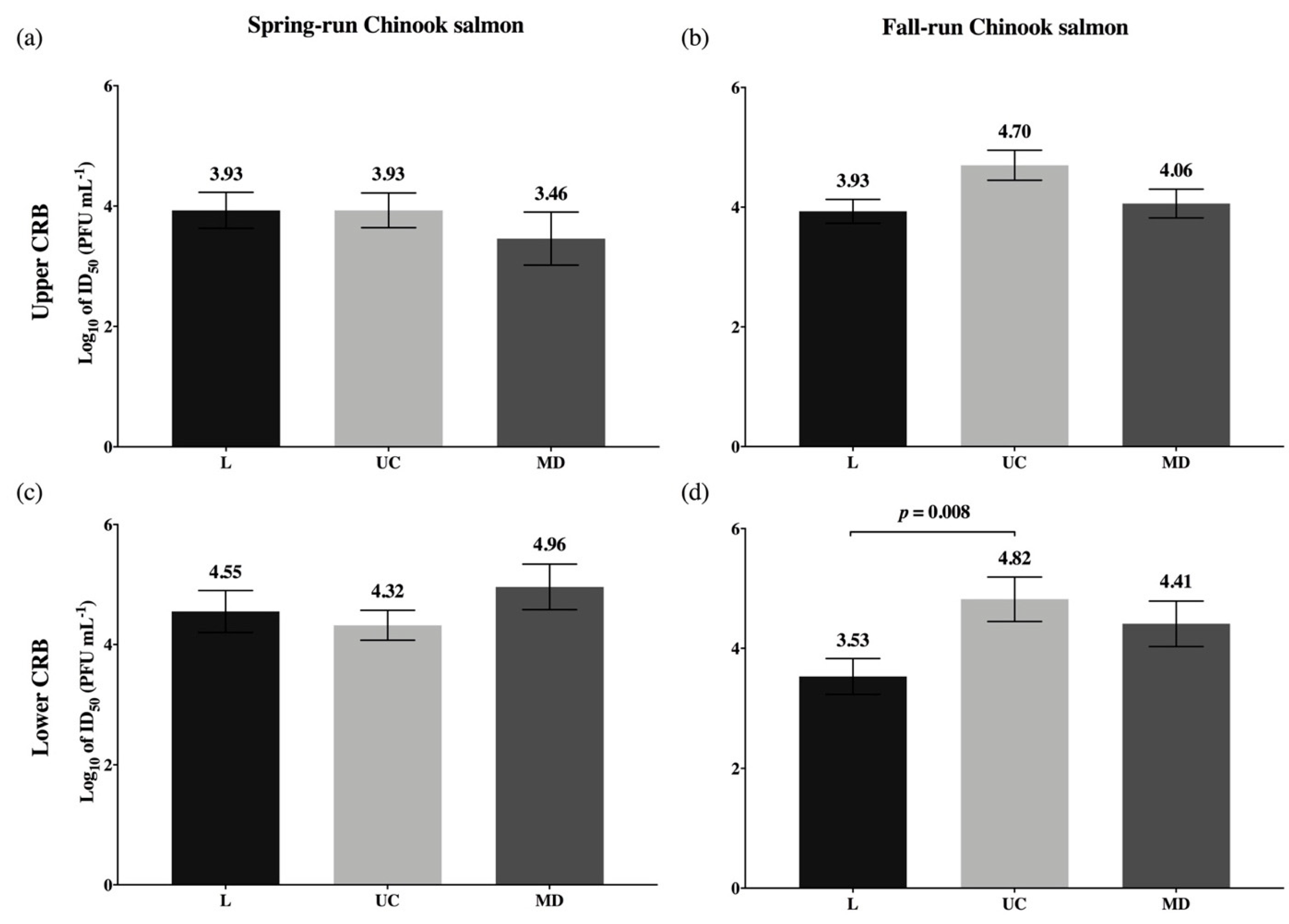
3.3. Viral Dosages Needed to Infect 50 Percent of a Host Population (ID50)
3.4. The Influence of Exposure Factors on Infection Status and Viral Load
3.5. The Influence of the Host Population on Infection Status and Viral Load
3.6. Persistence of IHNV Infection in Juvenile Columbia River Basin Chinook Salmon
3.7. Field Occurrence Patterns of UC and MD IHNV Infection in CRB Chinook Salmon
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kurath, G.; Garver, K.A.; Troyer, R.M.; Emmenegger, E.J.; Einer-Jensen, K.; Anderson, E.D. Phylogeography of infectious haematopoietic necrosis virus in North America. J. Gen. Virol. 2003, 84, 803–814. [Google Scholar] [CrossRef] [PubMed]
- Kurath, G. Fish novirhabdoviruses. In Rhabdoviruses: Molecular Taxonomy, Evolution, Genomics, Ecology, Host-Vector Interactions, Cytopathology, and Control; Dietzgen, R.G., Kuzman, I.V., Eds.; Caister Academic Press: Norfolk, UK, 2012; pp. 89–117. [Google Scholar]
- Hernandez, D.G.; Purcell, M.K.; Friedman, C.S.; Kurath, G. Susceptibility of ocean-and stream-type Chinook salmon to isolates of the L, U, and M genogroups of infectious hematopoietic necrosis virus (IHNV). Dis. Aquat. Org. 2016, 121, 15–28. [Google Scholar] [CrossRef] [PubMed]
- Breyta, R.; Brito, I.; Kurath, G.; LaDeau, S. Infectious hematopoietic necrosis virus virological and genetic surveillance 2000–2012. Ecology 2017, 98, 283. [Google Scholar] [CrossRef] [Green Version]
- Myers, J.M.; Kope, R.G.; Bryant, B.J.; Teel, D.; Lierheimer, L.J.; Wainwright, T.C.; Grant, W.S.; Waknitz, F.W.; Neely, K.; Lindley, S.T.; et al. Status Review of Chinook Salmon from Washington, Idaho, Oregon, and California; Memo NMFS-NWFSC-35; U.S. Department of Commerce, NOAA Tech: Washington, DC, USA, 1998.
- Brannon, E.L.; Powell, M.S.; Quinn, T.P.; Talbot, A. Population Structure of Columbia River Basin Chinook Salmon and Steelhead Trout. Rev. Fish. Sci. 2004, 12, 99–232. [Google Scholar] [CrossRef]
- Waples, R.S.; Teel, D.J.; Myers, J.M.; Marshall, A.R. Life—History divergence in Chinook salmon: Historic contingency and parallel evolution. Evolution 2004, 58, 386–403. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Waples, R.S.; Pess, G.R.; Beechie, T. Evolutionary history of Pacific salmon in dynamic environments. Evol. Appl. 2008, 1, 189–206. [Google Scholar] [CrossRef]
- Narum, S.R.; Hess, J.E.; Matala, A.P. Examining Genetic Lineages of Chinook Salmon in the Columbia River Basin. Trans. Am. Fish. Soc. 2010, 139, 1465–1477. [Google Scholar] [CrossRef]
- StreamNet GIS Data. Metadata for Pacific Northwest Chinook Salmon Fish Distribution Spatial Data Set; StreamNet: Portland, OR, USA, 2019; Available online: https://www.streamnet.org/data/interactive-maps-and-gis-data/ (accessed on 3 March 2019).
- Wolf, K. Infectious hematopoietic necrosis virus. In Fish Viruses and Fish Viral Diseases; Wolf, K., Ed.; Cornell University Press: Ithaca, NY, USA, 1988; pp. 83–114. [Google Scholar]
- Bootland, L.M.; Leong, J.C. Infectious haematopoietic necrosis virus. In Fish Diseases and Disorders; Woo, P.T.K., Leatherland, J.F., Bruno, D.W., Eds.; CAB International: Poole, UK, 2011; Volume 3, pp. 66–109. [Google Scholar]
- International Committee for Virus Taxonomy, Virus Taxonomy 2019 Release. Available online: https://talk.ictvonline.org/taxonomy (accessed on 3 January 2019).
- Garver, K.; Troyer, R.; Kurath, G. Two distinct phylogenetic clades of infectious hematopoietic necrosis virus overlap within the Columbia River basin. Dis. Aquat. Org. 2003, 55, 187–203. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Breyta, R.; Black, A.; Kaufman, J.; Kurath, G. Spatial and temporal heterogeneity of infectious hematopoietic necrosis virus in Pacific Northwest salmonids. Infect. Genet. Evol. 2016, 45, 347–358. [Google Scholar] [CrossRef] [Green Version]
- Kelley, G.O.; Bendorf, C.M.; Yun, S.C.; Kurath, G.; Hedrick, R.P. Genotypes and phylogeographical relation-ships of infectious hematopoietic necrosis virus in California, USA. Dis. Aquat. Org. 2007, 77, 29–40. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Black, A.; Breyta, R.; Bedford, T.; Kurath, G. Geography and host species shape the evolutionary dynamics of U genogroup infectious hematopoietic necrosis virus. Virus Evol. 2016, 2, vew034. [Google Scholar] [CrossRef] [Green Version]
- Columbia River DART Database (Data Access in Real Time), Columbia Basin Research. University of Washington, School of Aquatic & Fishery Sciences. Seattle (WA). Available online: http://www.cbr.washington.edu/dart (accessed on 8 March 2019).
- LaPatra, S.E. Strain Differentiation and Detection of Infectious Hematopoietic Necrosis Virus. Ph.D. Thesis, Oregon State University, Corvallis, Orgeon, 1989. [Google Scholar]
- Foott, J.S.; Free, D.; McDowell, T.; Arkush, K.D.; Hedrick, R.P. Infectious hematopoietic necrosis virus transmission and disease among juvenile Chinook salmon exposed in culture compared to environmentally relevant conditions. San Fr. Estuary Watershed Sci. 2006, 4, 1–16. [Google Scholar] [CrossRef] [Green Version]
- Bendorf, A.C.M. Phylogenetic Analysis and Virulence Characteristics for Chinook Salmon, Rainbow Trout, and Steelhead of L Genogroup Infectious Hematopoietic Necrosis Virus (IHNV) From The Feather River, California. Ph.D. Thesis, University of California, Davis, CA, USA, 2010. [Google Scholar]
- Wertheimer, A.C.; Winton, J.R. Differences in Susceptibility Among Three Stocks of Chinook Salmon, Oncorhynchus Tshawytscha, to Two Isolates of Infectious Hematopoietic Necrosis Virus. NOAA Technical Memorandum NMFS F/NWC-22, 1982. Available online: repository.library.noaa.gov (accessed on 18 March 2019).
- LaPatra, S.E.; Fryer, J.L.; Rohovec, J.S. Virulence comparison of different electrophero-types of infectious hematopoietic necrosis virus. Dis. Aquat. Org. 1993, 16, 115–120. [Google Scholar] [CrossRef]
- Taylor, E.B. Phenotypic Correlates of Life-History Variation in Juvenile Chinook Salmon, Oncorhynchus tshawytscha. J. Anim. Ecol. 1990, 59, 455. [Google Scholar] [CrossRef]
- Healey, M.C. The life history of Chinook salmon (Oncorhynchus tshawytscha). In Life History of Pacific Salmon; Groot, C., Margolis, L., Eds.; University of British Columbia Press: Vancouver, BC, Canada, 1991; pp. 311–393. [Google Scholar]
- World Organization for Animal Health (OIE). Recommendations for Surface Disinfection of Salmonid Eggs; Aquatic Animal Health Code; World Organization for Animal Health (OIE): Paris, France, 2019; Chapter 4.4. [Google Scholar]
- Garver, K.A.; Batts, W.N.; Kurath, G. Virulence Comparisons of Infectious Hematopoietic Necrosis Virus U and M Genogroups in Sockeye Salmon and Rainbow Trout. J. Aquat. Anim. Health 2006, 18, 232–243. [Google Scholar] [CrossRef] [PubMed]
- Breyta, R.; Jones, A.; Kurath, G. Differential susceptibility in steelhead trout populations to an emergent MD strain of infectious hematopoietic necrosis virus. Dis. Aquat. Org. 2014, 112, 17–28. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Breyta, R.; McKenney, D.; Tesfaye, T.; Ono, K.; Kurath, G. Increasing virulence, but not infectivity, associated with serially emergent virus strains of a fish rhabdovirus. Virus Evol. 2016, 2, vev018. [Google Scholar] [CrossRef] [Green Version]
- Wargo, A.R.; Garver, K.A.; Kurath, G. Virulence correlates with fitness in vivo for two M group genotypes of Infectious hematopoietic necrosis virus (IHNV). Virology 2010, 404, 51–58. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Purcell, M.; Thompson, R.; Garver, K.; Hawley, L.; Batts, W.; Sprague, L.; Sampson, C.; Winton, J.; Purcell, M. Universal reverse-transcriptase real-time PCR for infectious hematopoietic necrosis virus (IHNV). Dis. Aquat. Org. 2013, 106, 103–115. [Google Scholar] [CrossRef] [Green Version]
- McKenney, D.G.; Kurath, G.; Wargo, A.R. Characterization of infectious dose and lethal dose of two strains of infectious hematopoietic necrosis virus (IHNV). Virus Res. 2016, 214, 80–89. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2018; Available online: https://www.R-project.org (accessed on 8 March 2018).
- Crawley, M.J. The R Book; John Wiley and Sons: Hoboken, NJ, USA, 2012. [Google Scholar]
- Venables, W.N.; Ripley, B.D. Modern Applied Statistics with S-PLUS; Springer Science & Business Media: Berlin, Germany, 2013. [Google Scholar]
- StreamNet GIS Data. Columbia River Watershed Boundary Spatial Data Set; StreamNet: Portland, OR, USA, 2019; Available online: https://www.streamnet.org/data/interactive-maps-and-gis-data/ (accessed on 3 March 2019).
- Molecular Epidemiology of Aquatic Pathogens IHNV Database 2020. Available online: http://gis.nacse.org (accessed on 8 March 2019).
- Peñaranda, M.M.D.; Purcell, M.K.; Kurath, G. Differential virulence mechanisms of infectious hematopoietic necrosis virus in rainbow trout (Oncorhynchus mykiss) include host entry and virus replication kinetics. J. Gen. Virol. 2009, 90, 2172–2182. [Google Scholar] [CrossRef] [PubMed]
- Purcell, M.K.; Garver, K.A.; Conway, C.; Elliott, D.G.; Kurath, G. Infectious haematopoietic necrosis virus genogroup—Specific virulence mechanisms in sockeye salmon, Oncorhynchus nerka (Walbaum), from Redfish Lake, Idaho. J. Fish Dis. 2009, 32, 619–631. [Google Scholar] [CrossRef]
- Workenhe, S.T.; Rise, M.L.; Kibenge, M.J.; Kibenge, F.S. The fight between the teleost fish immune response and aquatic viruses. Mol. Immunol. 2010, 47, 2525–2536. [Google Scholar] [CrossRef]
- Purcell, M.K.; Laing, K.J.; Winton, J.R. Immunity to Fish Rhabdoviruses. Viruses 2012, 4, 140–166. [Google Scholar] [CrossRef] [Green Version]
- Winans, G.A. Genetic Variability in Chinook Salmon Stocks from the Columbia River Basin. N. Am. J. Fish. Manag. 1989, 9, 47–52. [Google Scholar] [CrossRef]
- Abdul-Aziz, O.I.; Mantua, N.J.; Myers, K.W. Potential climate change impacts on thermal habitats of Pacific salmon (Oncorhynchus spp.) in the North Pacific Ocean and adjacent seas. Can. J. Fish. Aquat. Sci. 2011, 68, 1660–1680. [Google Scholar] [CrossRef]
- Shelton, A.O.; Sullaway, G.H.; Ward, E.J.; Feist, B.E.; Somers, K.A.; Tuttle, V.J.; Satterthwaite, W.H. Redistribution of salmon populations in the northeast Pacific ocean in response to climate. Fish Fish. 2020, 1–15. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| (a) | ||||
|---|---|---|---|---|
| Columbia River Basin Geographic Region a | Adult Migration Timing | Juvenile Life-History | Phylogenetic Lineage b,c | Chinook Salmon Population c |
| Upper CRB | Spring-run | Stream-type | Interior Columbia River Spring (3) | Methow River (28) |
| Upper CRB | Fall-run | Ocean-type | Interior Columbia River Fall (2) | Hanford Reach (14) |
| Lower CRB | Spring-run | Stream-type | Willamette River Spring (1) | North Santiam River (8) |
| Lower CRB | Fall-run | Ocean-type | Lower Columbia River Fall (1) | Cowlitz River (1) |
| (b) | |||||
|---|---|---|---|---|---|
| Virus Strain | Genogroup, Subgroup | midG Type d | Host | Isolation Site | Year of Isolation |
| FR0031 | L | mG011L | Chinook salmon | Feather River Hatchery, CA | 2000 |
| QTS07 | MD | mG110M | Steelhead trout | Salmon River Hatchery, WA | 2007 |
| RB1 | UC | mG001U | Steelhead trout | Round Butte Hatchery, OR | 1975 |
| (a) | ||
|---|---|---|
| Host Population | Total # Fish Positive/Total # Exposed, for All Virus Strains | Log10 of Total Virus (Viral RNA Copies) |
| Up-Spring | 48/96 | 8.15 |
| Up-Fall | 38/96 | 8.27 |
| Low-Spring | 30/96 | 8.61 |
| Low-Fall | 39/96 | 8.92 |
| (b) | ||
|---|---|---|
| Virus Strain | Total # Fish Positive/Total # Exposed, for All Host Populations | Log10 of Total Virus (Viral RNA Copies) |
| L | 58/128 | 8.65 |
| UC | 45/128 | 8.91 |
| MD | 52/128 | 8.49 |
| mock | 0/30 | na |
| L | UC | MD | |||||
|---|---|---|---|---|---|---|---|
| (FR0031) | (RB1) | (QTS07) | |||||
| Host Population | Dose a | Mean Log Viral Load b | SEM | Mean Log Viral Load b | SEM | Mean Log Viral Load b | SEM |
| Up-Spring | 2 × 102 | - | - | - | - | 5.61 | 0.365 |
| 2 × 103 | 4.58 | na | 5.00 | 0.613 | 5.45 | 0.268 | |
| 2 × 104 | 5.38 | 0.316 | 4.56 | 0.628 | 5.68 | 0.374 | |
| 2 × 105 | 5.90 | 0.364 | 4.44 | 0.38 | 5.72 | 0.482 | |
| Mock | - | - | - | - | - | - | |
| Up-Fall | 2 × 102 | - | - | - | - | - | - |
| 2 × 103 | 5.44 | na | - | - | 5.41 | na | |
| 2 × 104 | 5.72 | 0.429 | 5.47 | 0.61 | 5.02 | 0.18 | |
| 2 × 105 | 6.29 | 0.426 | 5.71 | 0.513 | 6.08 | 0.30 | |
| Mock | - | - | - | - | - | - | |
| Low-Spring | 2 × 102 | - | - | - | - | - | - |
| 2 × 103 | 4.67 | na | 7.78 | na | 5.12 | na | |
| 2 × 104 | 6.98 | na | 5.79 | 0.662 | 6.41 | 0.9 | |
| 2 × 105 | 6.61 | 0.315 | 5.91 | 0.434 | 6.77 | 0.6 | |
| Mock | - | - | - | - | - | - | |
| Low-Fall | 2 × 102 | 6.68 | na | - | - | - | - |
| 2 × 103 | 6.63 | 0.592 | 8.43 | na | 5.68 | 0.505 | |
| 2 × 104 | 6.02 | 0.228 | 4.62 | 0.76 | 6.11 | 0.527 | |
| 2 × 105 | 6.69 | 0.269 | 6.04 | 0.734 | 6.10 | 0.234 | |
| IHNV Genogroup, Subgroup | L | UC | MD |
|---|---|---|---|
| (Experimental Virus Strain) | (FR0031) | (RB1) | (QTS07) |
| L | |||
| (FR0031) | - | 0.042 | 0.330 |
| UC | |||
| (RB1) | 0.030 | - | 0.280 |
| MD | |||
| (QTS07) | 0.315 | 0.237 | - |
| Exposure Dose | 0.001 * (infection status); 0.115 (log10 viral load) | ||
| Chinook Salmon Population | Up-Spring | Up-Fall | Low-Spring | Low-Fall |
|---|---|---|---|---|
| Up–Spring | - | 0.576 | 0.005 * | 0.008 b,c |
| Up–Fall | 0.056 | - | 0.027 c | 0.049 c |
| Low–Spring | 0.001 * | 0.119 | - | 0.707 |
| Low–Fall | 0.085 | 0.847 | 0.080 | - |
| L (FR0031) | UC (RB1) | MD (QTS07) | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Host Population | Num.pos. /Tested | Mean Log Viral Load | SEM a | Num.pos. /Tested | Mean Log Viral Load | SEM a | Num.pos./Tested | Mean Log Viral Load | SEM a | Total Fish pos. | Log10 Total Virus Quantity |
| Up-Spring | 4/8 | 6.67 | 0.56 | 4/8 | 5.77 | 0.031 | 2/8 | 5.34 | 0.23 | 10 | 8.02 |
| Up-Fall | 4/8 | 6.59 | 1.2 | 2/8 | 4.38 | 0.36 | 1/8 | 5.96 | na | 7 | 9.15 |
| Low-Spring | 2/8 | 5.87 | 0.97 | 1/8 | 5.83 | na | 1/8 | 5.18 | na | 4 | 6.89 |
| Low-Fall | 5/8 | 5.22 | 0.51 | 3/8 | 4.99 | 0.16 | 1/8 | 5.87 | na | 9 | 7.19 |
| Total fish pos. | 15 | 10 | 5 | ||||||||
| Log10 total virus quantity | 9.18 | 6.75 | 6.36 | ||||||||
| Adult Migration Timing (Juvenile Life-History) | Total Cohorts Tested a,b | Virus pos. Cohorts | % Virus Pos. | Pos. Cohorts Genotyped | U Pos. Cohorts | % U Pos. | M Pos. Cohorts | % M Pos. |
|---|---|---|---|---|---|---|---|---|
| Spring-run | ||||||||
| (stream-type) | 777 | 209 | 27% | 126 | 103 | 82% | 23 | 18% |
| Fall-run | ||||||||
| (ocean-type) | 172 | 43 | 25% | 26 | 23 | 88% | 3 | 12% |
| Summer-run | ||||||||
| (stream/ocean-type) c | 62 | 8 | 13% | 2 | 1 | 50% | 1 | 50% |
| Trait | Measure | Variation Observed | Data |
|---|---|---|---|
| SURVIVAL | |||
| Mortality after high dose virus exposure | |||
| Significant difference: Low-Spring is less susceptible to L virus than 3 other host populations. | Figure 2 | ||
| Mortality after moderate dose virus exposure | |||
| Significant difference: Low-Spring and Up-Spring are less susceptible to L virus than Up-Fall. | Figure 2 | ||
| INFECTION | |||
| Primary data for total # fish infected | |||
| Non-statistical observation: Low-Spring has the lowest and Up-Spring the highest # fish infected. | Table 2 | ||
| Primary data for total viral loads | |||
| Non-statistical observation: Low-Spring & Low-Fall have higher viral loads than Up-Spring & Up-Fall. | Table 2 | ||
| ID50 values | |||
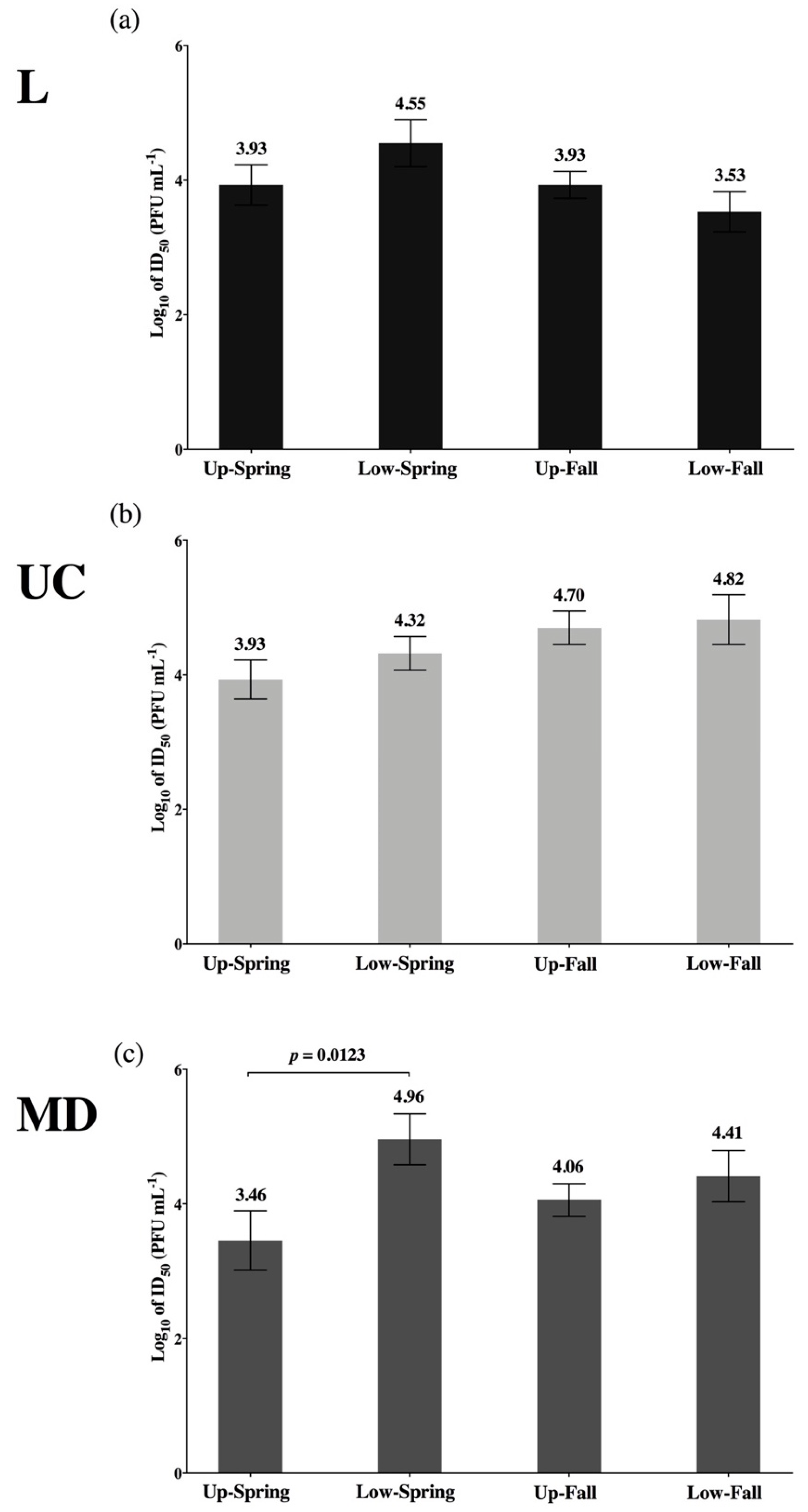
| Significant difference: Low-Spring is less susceptible than Up-Spring for MD virus only. Consistent trends: Low-Spring is also least susceptible to L virus; Up-Spring is most susceptible to UC virus. | Figure 6 | ||
| Infection status on day 3 post-exposure (logistic regression models) | |||
| Significant difference at p < 0.0083: Low-Spring is less susceptible to infection than Up-Spring (p = 0.001). | Table 5 | ||
| Viral load on day 3 post-exposure (generalized linear models) | |||
| Significant difference at p < 0.0083: Low-Spring has higher viral loads than Up-Spring (p = 0.005). Additional trends at p < 0.05: Low-Spring has higher viral loads than Up-Fall (p = 0.027); Low-Fall has higher viral loads than Up-Spring (p = 0.00849); Low-Fall has higher viral loads than Up-Fall (p = 0.05) | Table 5 | ||
| Persistence of detectable infection 30 days after virus exposure | Table 6 | ||
| Non-statistical observations: Low-Spring has the smallest number of fish infected; Low-Spring & Low-Fall lower total viral loads than Up-Spring & Up-Fall. | |||
| Trait | Measure | Variation Observed | Data |
|---|---|---|---|
| VIRULENCE | |||
| Mortality after high dose virus exposure | |||
| Significant differences: L virus causes higher mortality than UC or MD, in all host populations | Figure 2 | ||
| Mortality after moderate dose virus exposure | |||
| No significant differences | Figure 2 | ||
| INFECTION | |||
| Primary data for total # fish infected | |||
| Non-statistical observation: L virus infects more fish than UC or MD | Table 2 | ||
| Primary data for total viral loads | |||
| Non-statistical observation: UC virus has higher viral loads than L or MD | Table 2 | ||
| ID50 values | |||
| Significant difference: L virus is more infectious (lower ID50) than UC in Low-OT population only. Consistent trend: L virus is also more infectious (lower ID50) than UC in Up-OT population. | Figure 5 | ||
| Infection status on day 3 post-exposure (logistic regression models) | |||
| No significant difference at p < 0.0166. Trend at p < 0.05: L virus is more infectious than UC (p = 0.03). | Table 4 | ||
| Viral load on day 3 post-exposure (generalized linear models) | |||
| No significant difference at p < 0.0166. Trend at p < 0.05: UC virus has higher viral load than L (p = 0.042). | Table 4 | ||
| Persistence of detectable infection 30 days after virus exposure | Table 6 | ||
| Non-statistical observations: L virus has highest number of detectable persistence infections, MD has the lowest; L virus has higher total viral load than UC or MD. | |||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hernandez, D.G.; Brown, W.; Naish, K.A.; Kurath, G. Virulence and Infectivity of UC, MD, and L Strains of Infectious Hematopoietic Necrosis Virus (IHNV) in Four Populations of Columbia River Basin Chinook Salmon. Viruses 2021, 13, 701. https://doi.org/10.3390/v13040701
Hernandez DG, Brown W, Naish KA, Kurath G. Virulence and Infectivity of UC, MD, and L Strains of Infectious Hematopoietic Necrosis Virus (IHNV) in Four Populations of Columbia River Basin Chinook Salmon. Viruses. 2021; 13(4):701. https://doi.org/10.3390/v13040701
Chicago/Turabian StyleHernandez, Daniel G., William Brown, Kerry A. Naish, and Gael Kurath. 2021. "Virulence and Infectivity of UC, MD, and L Strains of Infectious Hematopoietic Necrosis Virus (IHNV) in Four Populations of Columbia River Basin Chinook Salmon" Viruses 13, no. 4: 701. https://doi.org/10.3390/v13040701