
Recombinant Pseudorabies Virus with TK/gE Gene Deletion and Flt3L Co-Expression Enhances the Innate and Adaptive Immune Response via Activating Dendritic Cells

Abstract
:1. Introduction
2. Materials and Methods
2.1. Cells and Virus
2.2. Plasmid Construction
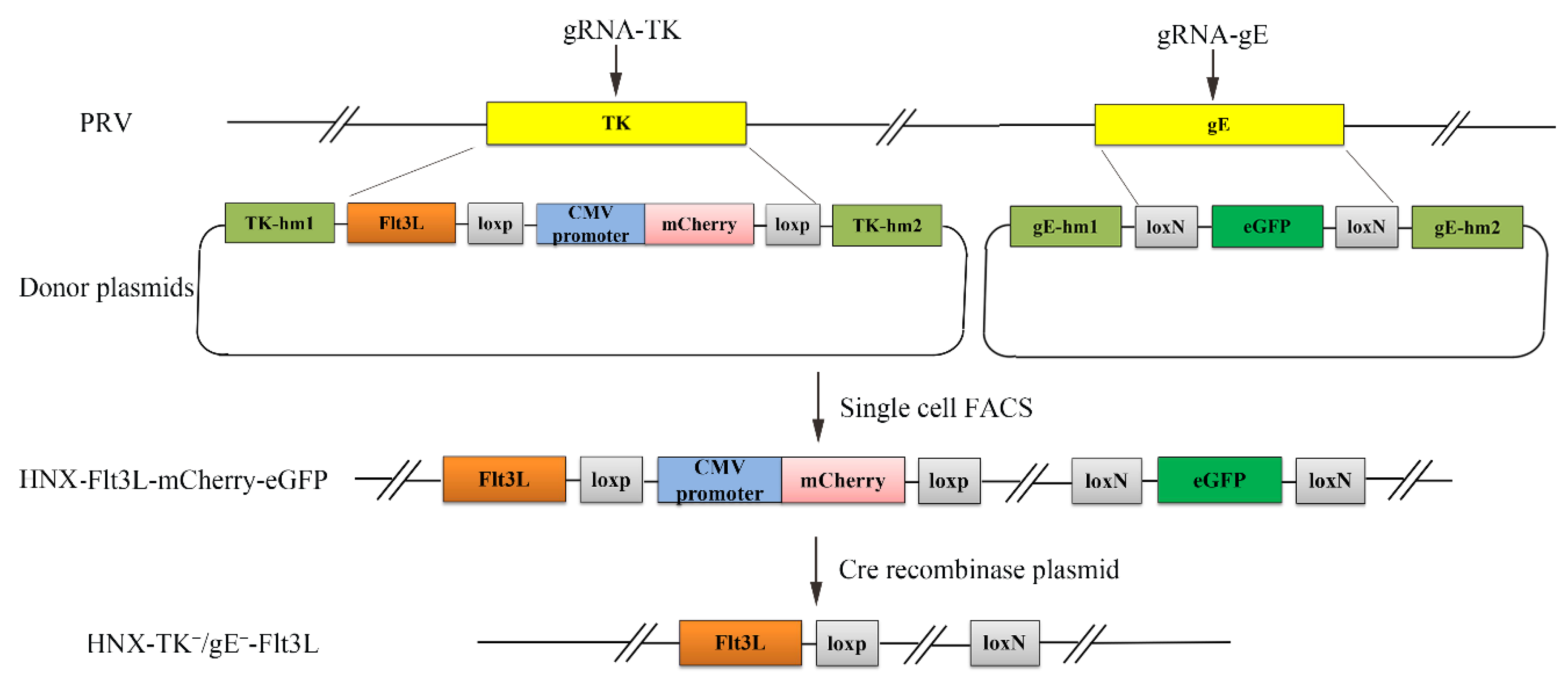
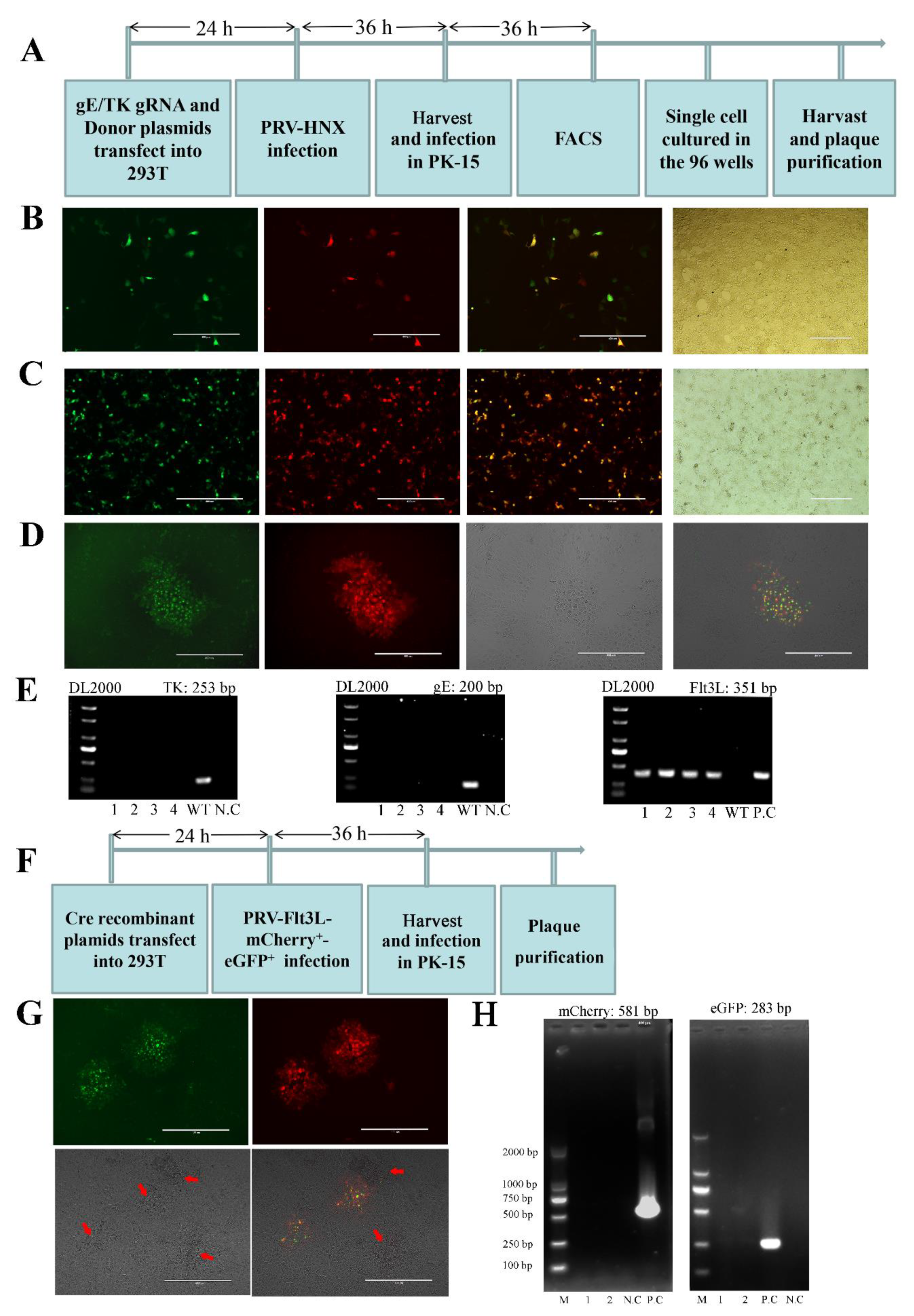
2.3. CRISPR/Cas9-Mediated Gene Insertion
2.4. Fluorescence-Activated Cell Sorting (FACS)
2.5. Virus Preparation, Propagation and Plaque Purification
2.6. Cre/Lox-Mediated Marker Genes Excision
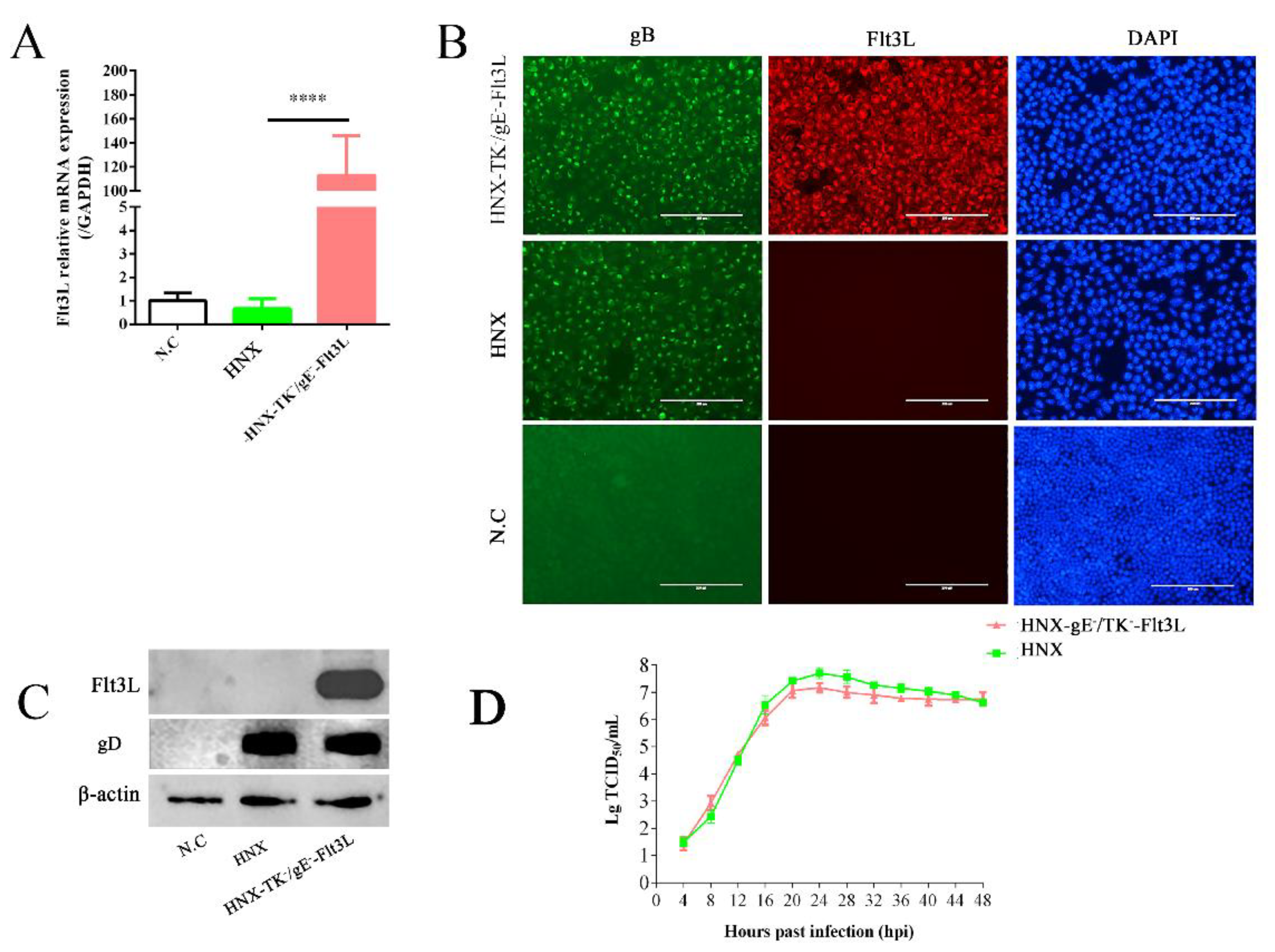
2.7. RNA Isolation and Quantitative Real-Time PCR (RT-qPCR)
2.8. Western Blot Analysis
2.9. Immunofluorescence Assay (IFA)
2.10. Replication Kinetics of the Rescued Recombinant PRV
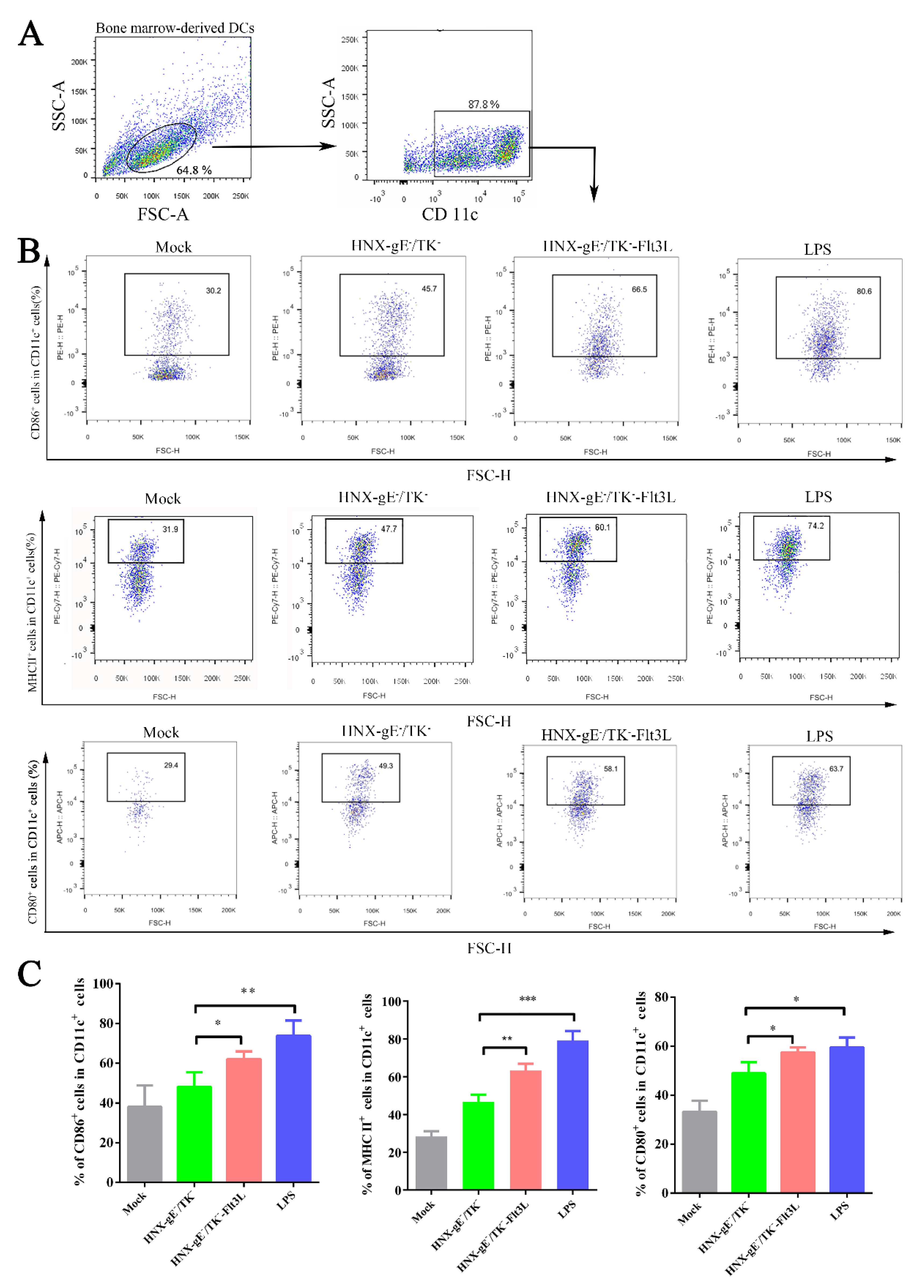
2.11. Collection of Bone Marrow-Derived DCs
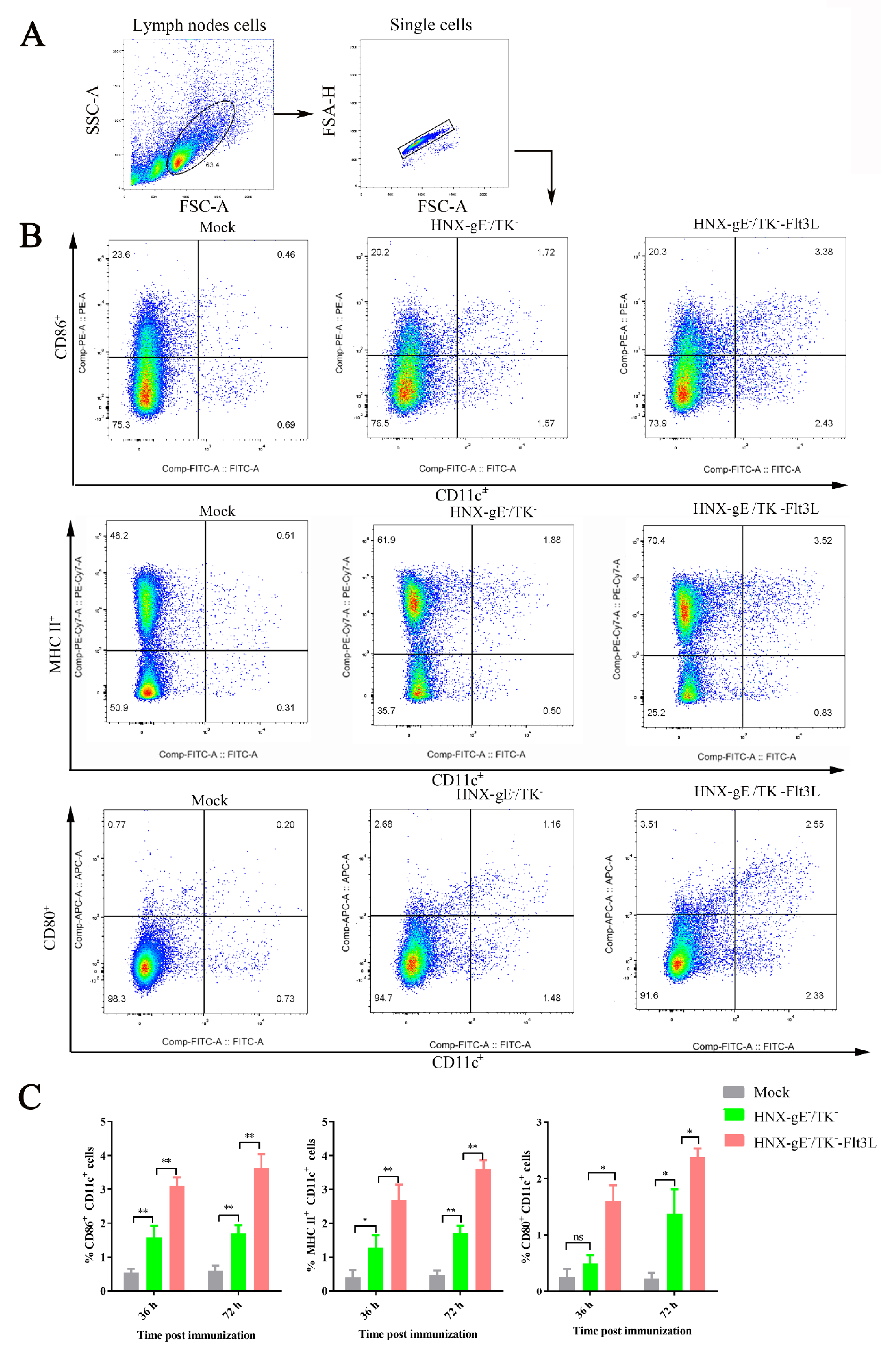
2.12. Sample Preparation and Flow Cytometry Analysis
2.13. Mice Immunization and Challenge Experiment
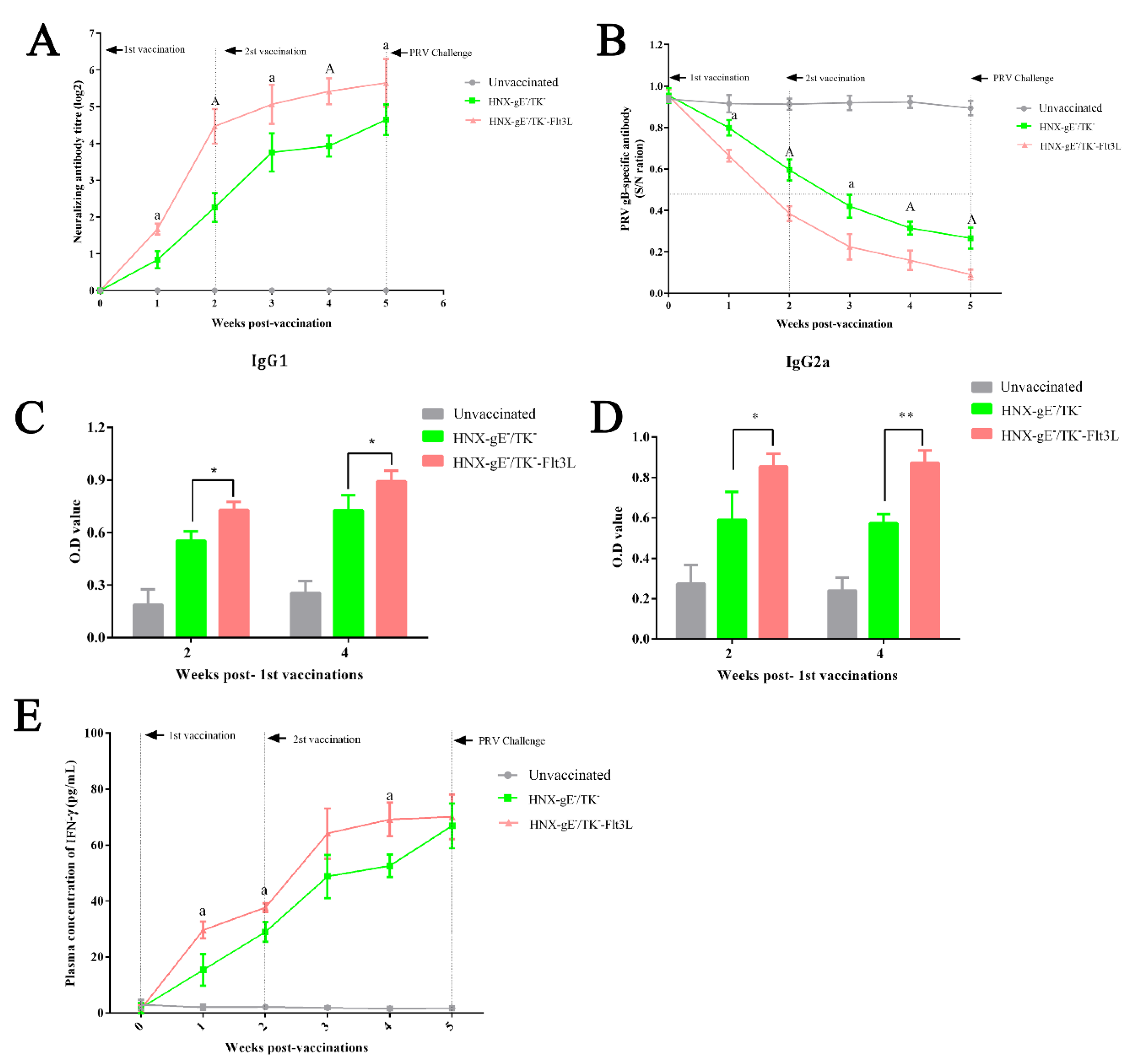
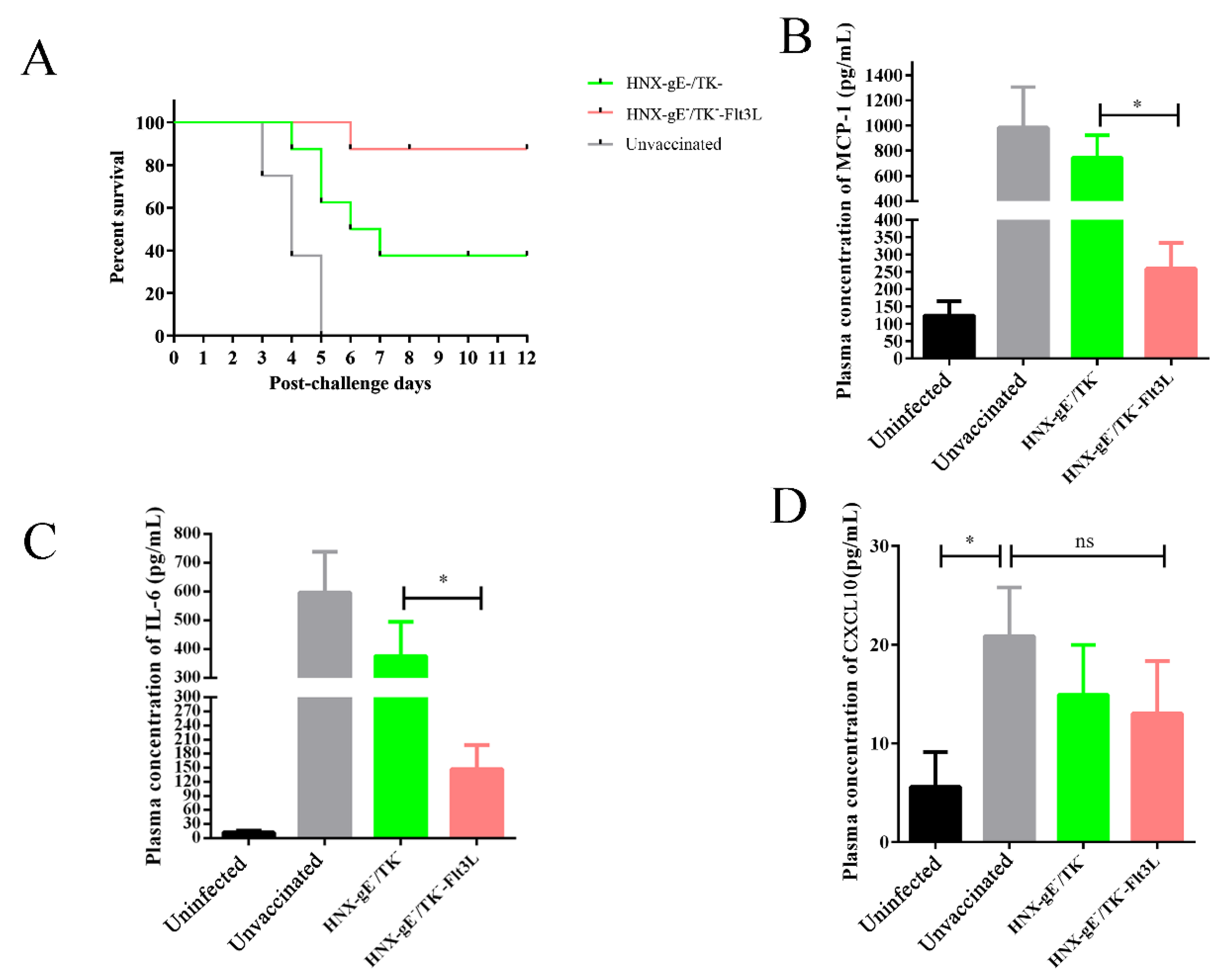
2.14. Antibody Detectionand Cytokine Detection
2.15. Neutralizing Antibody Assay
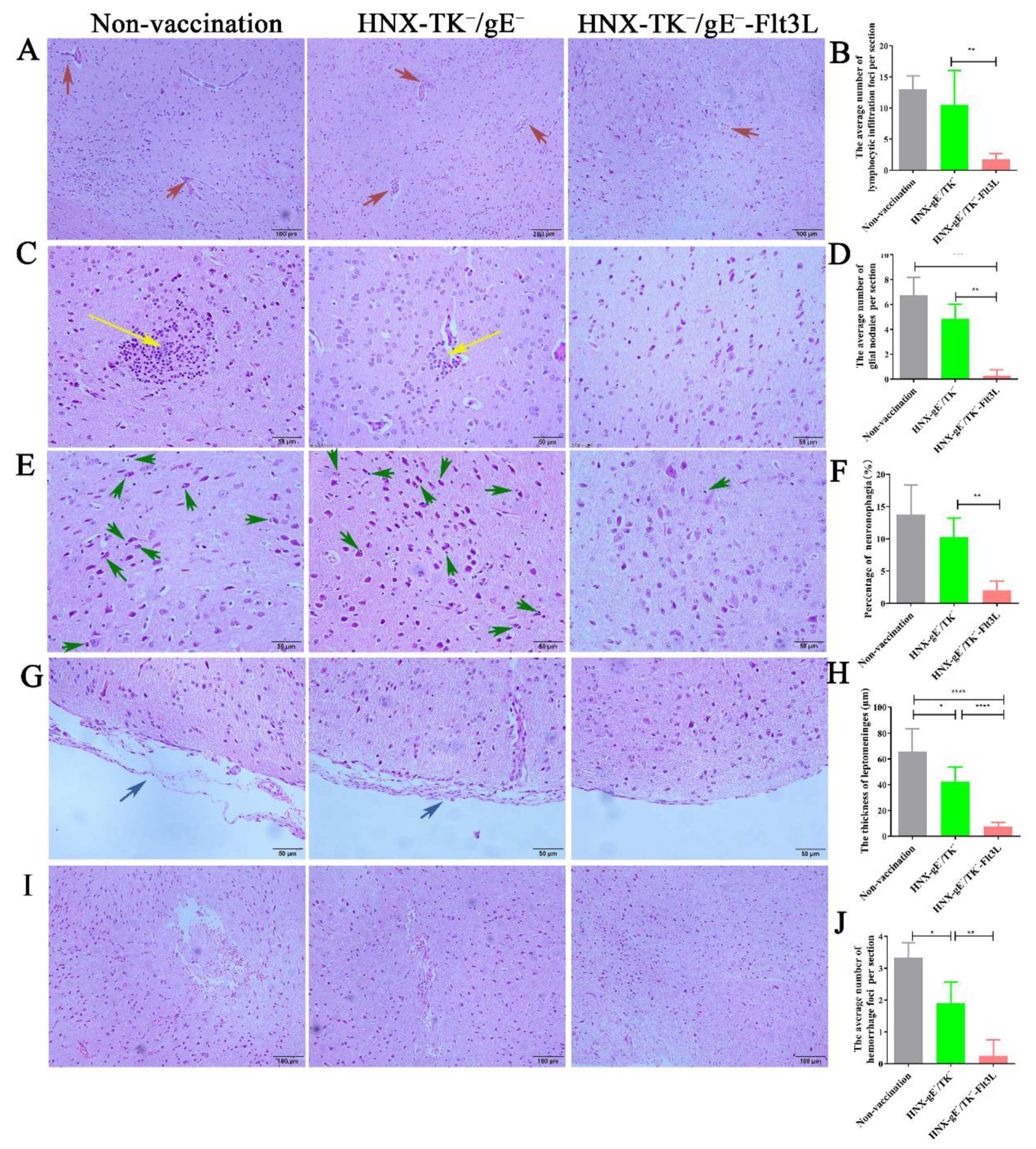
2.16. Histopathology
2.17. Statistical Analysis
3. Results
3.1. Construction of a Recombinant PRV by CRISPR/Cas9 and Cre/Lox System
3.2. Characterization of Recombinant HNX-TK−/gE−-Flt3L
3.3. Activation of Bone Marrow-Derived DCs by HNX-TK−/gE−-Flt3L In Vitro
3.4. Immune Responses after Immunization with HNX-TK−/gE−-Flt3L in Mice
3.5. Protective Effects of HNX-TK−/gE−-Flt3L against Lethal PRV Infection
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Klupp, B.G.; Hengartner, C.J.; Mettenleiter, T.C.; Enquist, L.W. Complete, annotated sequence of the pseudorabies virus genome. J. Virol. 2004, 78, 424–440. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pomeranz, L.E.; Reynolds, A.E.; Hengartner, C.J. Molecular biology of pseudorabies virus: Impact on neurovirology and veterinary medicine. Microbiol. Mol. Biol. Rev. 2005, 69, 462–500. [Google Scholar] [CrossRef] [Green Version]
- Tan, F.; Li, X.; Tian, K. Generating Recombinant Pseudorabies Virus for Use as a Vaccine Platform. Methods Mol. Biol. 2017, 1581, 79–96. [Google Scholar]
- Ben-Porat, T.; Rixon, F.J.; Blankenship, M.L. Analysis of the structure of the genome of pseudorabies virus. Virology 1979, 95, 285–294. [Google Scholar] [CrossRef]
- Hu, R.-M.; Zhou, Q.; Song, W.-B.; Sun, E.-C.; Zhang, M.-M.; He, Q.-G.; Chen, H.-C.; Wu, B.; Liu, Z.-F. Novel pseudorabies virus variant with defects in TK, gE and gI protects growing pigs against lethal challenge. Vaccine 2015, 33, 5733–5740. [Google Scholar] [CrossRef]
- Qian, P.; Zhi, X.; Wang, B.; Zhang, H.; Chen, H.; Li, X. Construction and immune efficacy of recombinant pseudorabies virus expressing PrM-E proteins of Japanese encephalitis virus genotype capital I.; Ukrainian. Virol. J. 2015, 12, 214. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Y.; Qiao, S.; Li, X.; Xie, W.; Guo, J.; Li, Q.; Liu, X.; Hou, J.; Xu, Y.; Wang, L.; et al. Molecular epidemiology of outbreak-associated pseudorabies virus (PRV) strains in central China. Virus Genes 2015, 50, 401–409. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; Guo, L.; Jia, X.; Wang, T.; Wang, J.; Sun, Z.; Wang, L.; Li, X.; Tan, F.; Tian, K. Construction of a triple gene-deleted Chinese Pseudorabies virus variant and its efficacy study as a vaccine candidate on suckling piglets. Vaccine 2015, 33, 2432–2437. [Google Scholar] [CrossRef] [PubMed]
- Ma, Z.; Han, Z.; Liu, Z.; Meng, F.; Wang, H.; Cao, L.; Li, Y.; Jiao, Q.; Liu, S.; Liu, M. Epidemiological investigation of porcine pseudorabies virus and its coinfection rate in Shandong Province in China from 2015 to 2018. J. Vet. Sci. 2020, 21, e36. [Google Scholar] [CrossRef] [PubMed]
- Zhou, H.; Pan, Y.; Liu, M.; Han, Z. Prevalence of Porcine Pseudorabies Virus and Its Coinfection Rate in Heilongjiang Province in China from 2013 to 2018. Viral Immunol. 2020, 33, 550–554. [Google Scholar] [CrossRef]
- Casades-Martí, L.; González-Barrio, D.; Royo-Hernández, L.; Díez-Delgado, I.; Ruiz-Fons, F. Dynamics of Aujeszky’s disease virus infection in wild boar in enzootic scenarios. Transbound. Emerg. Dis. 2020, 67, 388–405. [Google Scholar] [CrossRef]
- Pacini, M.I.; Forzan, M.; Cilia, G.; Bernardini, L.; Marzoli, F.; Pedonese, F.; Bandecchi, P.; Fratini, F.; Mazzei, M. Detection of Pseudorabies Virus in Wild Boar Foetus. Animals 2020, 10, 366. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Q.; Wang, X.; Xie, C.; Ding, S.; Yang, H.; Guo, S.; Li, J.; Qin, L.; Ban, F.; Wang, D.; et al. A Novel Human Acute Encephalitis Caused by Pseudorabies Virus Variant Strain. Clin. Infect. Dis. 2020. [Google Scholar] [CrossRef]
- Wang, D.; Tao, X.; Fei, M.; Chen, J.; Guo, W.; Li, P.; Wang, J. Human encephalitis caused by pseudorabies virus infection: A case report. J. NeuroVirol. 2020, 26, 442–448. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.; Guan, H.; Li, C.; Li, Y.; Wang, S.; Zhao, X.; Zhao, Y.; Liu, Y. Characteristics of human encephalitis caused by pseudorabies virus: A case series study. Int. J. Infect. Dis. 2019, 87, 92–99. [Google Scholar] [CrossRef] [Green Version]
- Moser, M.; Murphy, K.M. Dendritic cell regulation of TH1-TH2 development. Nat. Immunol. 2000, 1, 199–205. [Google Scholar] [CrossRef] [PubMed]
- Yuan, X.; Qin, X.; Wang, D.; Zhang, Z.; Tang, X.; Gao, X.; Chen, W.; Sun, L. Mesenchymal stem cell therapy induces FLT3L and CD1c+ dendritic cells in systemic lupus erythematosus patients. Nat. Commun. 2019, 10, 2498. [Google Scholar] [CrossRef]
- Ding, Y.; Wilkinson, A.; Idris, A.; Fancke, B.; O’Keeffe, M.; Khalil, D.; Ju, X.; Lahoud, M.H.; Caminschi, I.; Shortman, K.; et al. FLT3-Ligand Treatment of Humanized Mice Results in the Generation of Large Numbers of CD141+and CD1c+Dendritic Cells In Vivo. J. Immunol. 2014, 192, 1982–1989. [Google Scholar] [CrossRef] [Green Version]
- Waskow, C.; Liu, K.; Darrasse-Jèze, G.; Guermonprez, P.; Ginhoux, F.; Merad, M.; Shengelia, T.; Yao, K.; Nussenzweig, M.C. The receptor tyrosine kinase Flt3 is required for dendritic cell development in peripheral lymphoid tissues. Nat. Immunol. 2008, 9, 676–683. [Google Scholar] [CrossRef]
- Wen, Y.; Wang, H.; Wu, H.; Yang, F.; Tripp, R.A.; Hogan, R.J.; Fu, Z.F. Rabies Virus Expressing Dendritic Cell-Activating Molecules Enhances the Innate and Adaptive Immune Response to Vaccination. J. Virol. 2010, 85, 1634–1644. [Google Scholar] [CrossRef] [Green Version]
- Zhao, L.; Toriumi, H.; Wang, H.; Kuang, Y.; Guo, X.; Morimoto, K.; Fu, Z.F. Expression of MIP-1α (CCL3) by a Recombinant Rabies Virus Enhances Its Immunogenicity by Inducing Innate Immunity and Recruiting Dendritic Cells and B Cells. J. Virol. 2010, 84, 9642–9648. [Google Scholar] [CrossRef] [Green Version]
- Barry, K.C.; Hsu, J.; Broz, M.L.; Cueto, F.J.; Binnewies, M.; Combes, A.J.; Nelson, A.E.; Loo, K.; Kumar, R.; Rosenblum, M.D.; et al. A natural killer–dendritic cell axis defines checkpoint therapy–responsive tumor microenvironments. Nat. Med. 2018, 24, 1178–1191. [Google Scholar] [CrossRef] [PubMed]
- Borca, M.V.; Holinka, L.G.; Berggren, K.A.; Gladue, D.P. CRISPR-Cas9, a tool to efficiently increase the development of recombinant African swine fever viruses. Sci. Rep. 2018, 8, 3154. [Google Scholar] [CrossRef] [Green Version]
- Chang, P.; Yao, Y.; Tang, N.; Sadeyen, J.-R.; Sealy, J.; Clements, A.; Bhat, S.; Munir, M.; Bryant, J.E.; Iqbal, M. The Application of NHEJ-CRISPR/Cas9 and Cre-Lox System in the Generation of Bivalent Duck Enteritis Virus Vaccine against Avian Influenza Virus. Viruses 2018, 10, 81. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liang, X.; Sun, L.; Yu, T.; Pan, Y.; Wang, D.; Hu, X.; Fu, Z.; Zhenfang, F.; Cao, G. A CRISPR/Cas9 and Cre/Lox system-based express vaccine development strategy against re-emerging Pseudorabies virus. Sci. Rep. 2016, 6, 19176. [Google Scholar] [CrossRef] [Green Version]
- Jinek, M.; Chylinski, K.; Fonfara, I.; Hauer, M.; Doudna, J.A.; Charpentier, E. A Programmable dual-RNA-guided DNA endonuclease in adaptive bacterial immunity. Science 2012, 337, 816–821. [Google Scholar] [CrossRef] [PubMed]
- Mali, P.; Yang, L.; Esvelt, K.M.; Aach, J.; Guell, M.; DiCarlo, J.E.; Norville, J.E.; Church, G.M. RNA-Guided Human Genome Engineering via Cas9. Science 2013, 339, 823–826. [Google Scholar] [CrossRef] [Green Version]
- Ran, F.A.; Hsu, P.D.; Wright, J.; Agarwala, V.; Scott, D.A.; Zhang, F. Genome engineering using the CRISPR-Cas9 system. Nat. Protoc. 2013, 8, 2281–2308. [Google Scholar] [CrossRef] [Green Version]
- Cong, L.; Ran, F.A.; Cox, D.; Lin, S.; Barretto, R.; Habib, N.; Hsu, P.D.; Wu, X.; Jiang, W.; Marraffini, L.A.; et al. Multiplex Genome Engineering Using CRISPR/Cas Systems. Science 2013, 339, 819–823. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sternberg, N.; Hamilton, D. Bacteriophage P1 site-specific recombination. I. Recombination between loxP sites. J. Mol. Biol. 1981, 150, 467–486. [Google Scholar] [CrossRef]
- Ye, S.; Shao, K.; Li, Z.; Guo, N.; Zuo, Y.; Li, Q.; Lu, Z.; Chen, L.; He, Q.; Han, H. Antiviral Activity of Graphene Oxide: How Sharp Edged Structure and Charge Matter. ACS Appl. Mater. Interfaces 2015, 7, 21571–21579. [Google Scholar] [CrossRef]
- Gilboa, E. DC-based cancer vaccines. J. Clin. Investig. 2007, 117, 1195–1203. [Google Scholar] [CrossRef] [PubMed]
- Lutz, M.B.; Kukutsch, N.; Ogilvie, A.L.; Rößner, S.; Koch, F.; Romani, N.; Schuler, G. An advanced culture method for generating large quantities of highly pure dendritic cells from mouse bone marrow. J. Immunol. Methods 1999, 223, 77–92. [Google Scholar] [CrossRef]
- Zhang, Y.; Zhou, M.; Li, Y.; Luo, Z.; Chen, H.; Cui, M.; Fu, Z.F.; Zhao, L. Recombinant rabies virus with the glycoprotein fused with a DC-binding peptide is an efficacious rabies vaccine. Oncotarget 2017, 9, 831–841. [Google Scholar] [CrossRef] [Green Version]
- Maresch, C.; Lange, E.; Teifke, J.P.; Fuchs, W.; Klupp, B.; Müller, T.; Mettenleiter, T.C.; Vahlenkamp, T.W. Oral immunization of wild boar and domestic pigs with attenuated live vaccine protects against Pseudorabies virus infection. Vet. Microbiol. 2012, 161, 20–25. [Google Scholar] [CrossRef]
- Wang, J.; Song, Z.; Ge, A.; Guo, R.; Qiao, Y.; Xu, M.; Wang, Z.; Liu, Y.; Zheng, Y.; Fan, H.; et al. Safety and immunogenicity of an attenuated Chinese pseudorabies variant by dual deletion of TK&gE genes. BMC Vet. Res. 2018, 14, 287. [Google Scholar]
- Laval, K.; Vernejoul, J.B.; Van Cleemput, J.; Koyuncu, O.O.; Enquist, L.W. Virulent Pseudorabies Virus Infection Induces a Specific and Lethal Systemic Inflammatory Response in Mice. J. Virol. 2018, 92, e01614-18. [Google Scholar] [CrossRef] [Green Version]
- Freuling, C.M.; Muller, T.F.; Mettenleiter, T.C. Vaccines against pseudorabies virus (PrV). Vet. Microbiol. 2017, 206, 3–9. [Google Scholar] [CrossRef]
- Chen, M.; Chen, X.; Song, X.; Muhammad, A.; Jia, R.; Zou, Y.; Yin, L.; Li, L.; He, C.; Ye, G.; et al. The immune-adjuvant activity and the mechanism of resveratrol on pseudorabies virus vaccine in a mouse model. Int. Immunopharmacol. 2019, 76, 105876. [Google Scholar] [CrossRef] [PubMed]
- Bastola, R.; Seo, J.E.; Noh, G.; Keum, T.; Kim, J.H.; Shin, J.I.; Lee, S.; Lee, S. Determination of Mycoplasma hyopneumoniae-Specific IgG, IgG1, and IgG2a Titers in BALB/c Mice Induced by Mineral Oil-Based Oil-in-Water Emulsion Adjuvants Prepared Using a Self-Emulsifying Drug Delivery System. AAPS PharmSciTech 2019, 20, 31. [Google Scholar] [CrossRef]
- Gołoś, A.; Lutyńska, A. Aluminium-adjuvanted vaccines—A review of the current state of knowledge. Przeglad Epidemiol. 2015, 69, 731–734. [Google Scholar]
- Ma, R.; Zheng, H.; Liu, Q.; Wu, D.; Li, W.; Xu, S.; Cai, X.; Li, R. Exploring the interactions between engineered nanomaterials and immune cells at 3D nano-bio interfaces to discover potent nano-adjuvants. Nanomed. Nanotechnol. Biol. Med. 2019, 21, 102037. [Google Scholar] [CrossRef] [PubMed]
- McKee, A.S.; Munks, M.W.; Marrack, P. How Do Adjuvants Work? Important Considerations for New Generation Adjuvants. Immunity 2007, 27, 687–690. [Google Scholar] [CrossRef] [Green Version]
- Zheng, H.-H.; Wang, L.-Q.; Fu, P.-F.; Zheng, L.-L.; Chen, H.-Y.; Liu, F. Characterization of a recombinant pseudorabies virus expressing porcine parvovirus VP2 protein and porcine IL-6. Virol. J. 2020, 17, 19. [Google Scholar] [CrossRef]
- Mohammed, H.; Olivares-Fuster, O.; LaFrentz, S.; Arias, C.R. New attenuated vaccine against columnaris disease in fish: Choosing the right parental strain is critical for vaccine efficacy. Vaccine 2013, 31, 5276–5280. [Google Scholar] [CrossRef]
- Yu, T.; Chen, F.; Ku, X.; Fangzhou, C.; Zhu, Y.; Ma, H.; Li, S.; Wu, B.; He, Q. Growth characteristics and complete genomic sequence analysis of a novel pseudorabies virus in China. Virus Genes 2016, 52, 474–483. [Google Scholar] [CrossRef]
- He, W.; Auclert, L.Z.; Zhai, X.; Wong, G.; Zhang, C.; Zhu, H.; Xing, G.; Wang, S.; He, W.; Li, K.; et al. Interspecies Transmission, Genetic Diversity, and Evolutionary Dynamics of Pseudorabies Virus. J. Infect. Dis. 2018, 219, 1705–1715. [Google Scholar] [CrossRef] [PubMed]
- Zurkova, K.; Hainz, P.; Krystofova, J.; Kutinová, L.; Sanda, M.; Nemeckova, S. Attenuation of vaccinia virus by the expression of human Flt3 ligand. Virol. J. 2010, 7, 109. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Yang, J.; Li, M.; Cui, M.; Fu, Z.F.; Zhao, L.; Zhou, M. A Recombinant Rabies Virus Expressing Fms-like Tyrosine Kinase 3 Ligand (Flt3L) Induces Enhanced Immunogenicity in Mice. Virol. Sin. 2019, 34, 662–672. [Google Scholar] [CrossRef] [PubMed]
- Broom, L.J.; Kogut, M.H. Inflammation: Friend or foe for animal production? Poult. Sci. 2018, 97, 510–514. [Google Scholar] [CrossRef]
- Petousis-Harris, H. Vaccine injection technique and reactogenicity—Evidence for practice. Vaccine 2008, 26, 6299–6304. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Clinical Symptoms | Non Vaccinated | HNX-TK−/gE− | HNX-TK−/gE−-Flt3L |
|---|---|---|---|
| pruritus | 8/8 + + + | 2/8 +, 5/8 + + | 1/8 + + |
| ruffled fur | 8/8 + + + | 7/8 + + | 2/8 +, 1/8 + + |
| hyperkinesia | 8/8 + + + | 7/8 + + | 3/8 + + |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yao, L.; Hu, Q.; Chen, S.; Zhou, T.; Yu, X.; Ma, H.; H. Ghonaim, A.; Wu, H.; Sun, Q.; Fan, S.; et al. Recombinant Pseudorabies Virus with TK/gE Gene Deletion and Flt3L Co-Expression Enhances the Innate and Adaptive Immune Response via Activating Dendritic Cells. Viruses 2021, 13, 691. https://doi.org/10.3390/v13040691
Yao L, Hu Q, Chen S, Zhou T, Yu X, Ma H, H. Ghonaim A, Wu H, Sun Q, Fan S, et al. Recombinant Pseudorabies Virus with TK/gE Gene Deletion and Flt3L Co-Expression Enhances the Innate and Adaptive Immune Response via Activating Dendritic Cells. Viruses. 2021; 13(4):691. https://doi.org/10.3390/v13040691
Chicago/Turabian StyleYao, Lun, Qiao Hu, Siqi Chen, Tong Zhou, Xuexiang Yu, Hailong Ma, Ahmed. H. Ghonaim, Hao Wu, Qi Sun, Shengxian Fan, and et al. 2021. "Recombinant Pseudorabies Virus with TK/gE Gene Deletion and Flt3L Co-Expression Enhances the Innate and Adaptive Immune Response via Activating Dendritic Cells" Viruses 13, no. 4: 691. https://doi.org/10.3390/v13040691