
Survival Strategies of Streptococcus pyogenes in Response to Phage Infection

Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacterial Strains
2.2. Phage Propagation
2.3. Bacteriophage A1‒Membrane Vesicle Interaction Assays
2.4. Electron Microscopy and Image Analysis
2.5. One-Step Growth Analysis
2.6. Spectrophotometric Phage A1 Lysis Profile
2.7. Phage A1 Efficiency of Plating on M1 Serotype S. pyogenes
2.8. Lysogeny and CRISPR Acquisition Testing
2.9. Genomic DNA Isolation
2.10. Genome Sequencing and Bioinformatic Analysis
2.11. Phylogenetic Analysis
2.12. Cell Line Maintanance and Differentiation
2.13. Infection Assay
2.14. Antibiotic Susceptibility
2.15. Statistics
3. Results
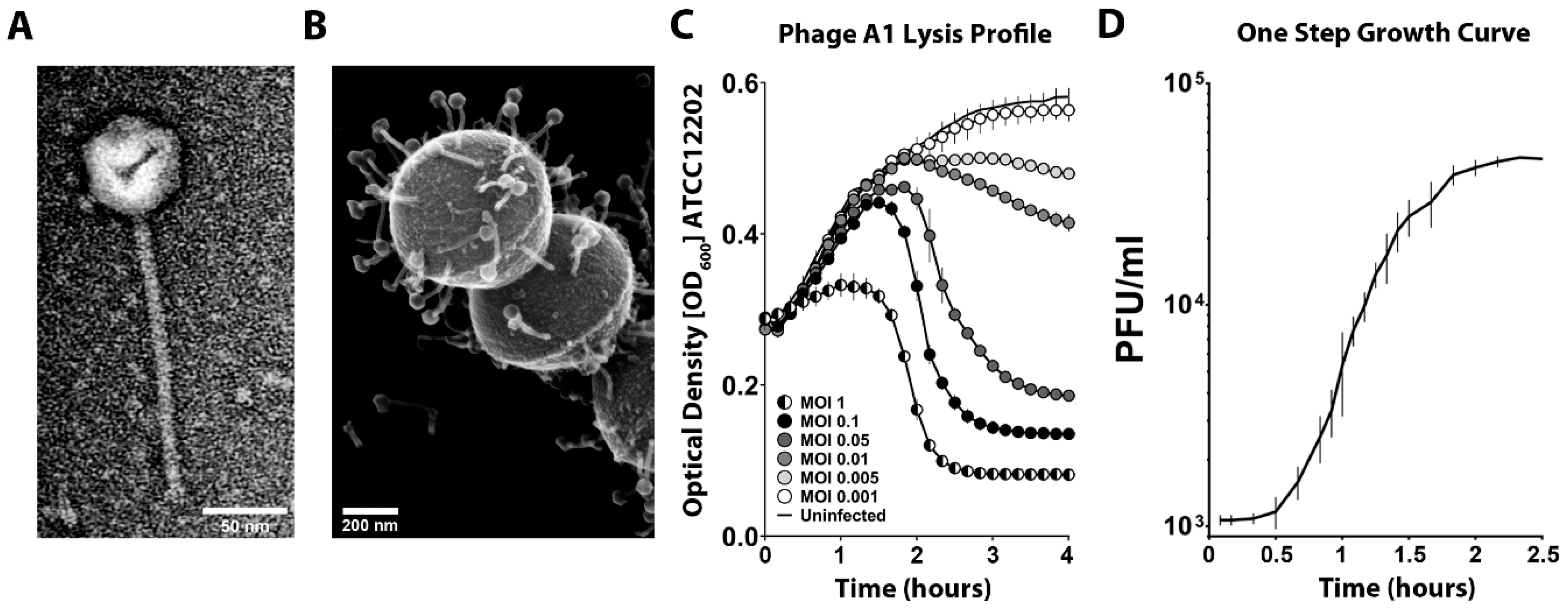
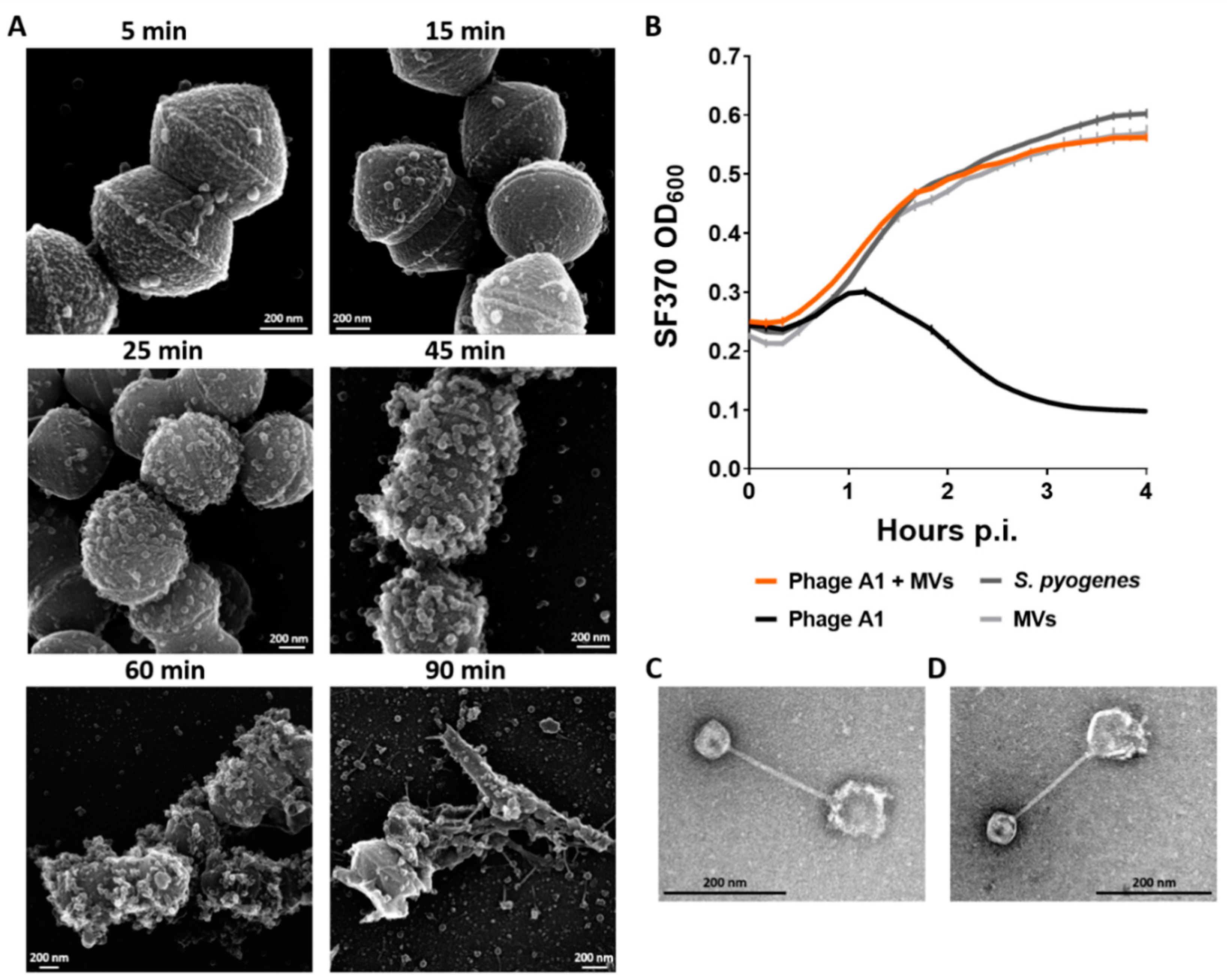
3.1. Virion Morphology and Physiological Characterization of S. pyogenes Bacteriophage A1
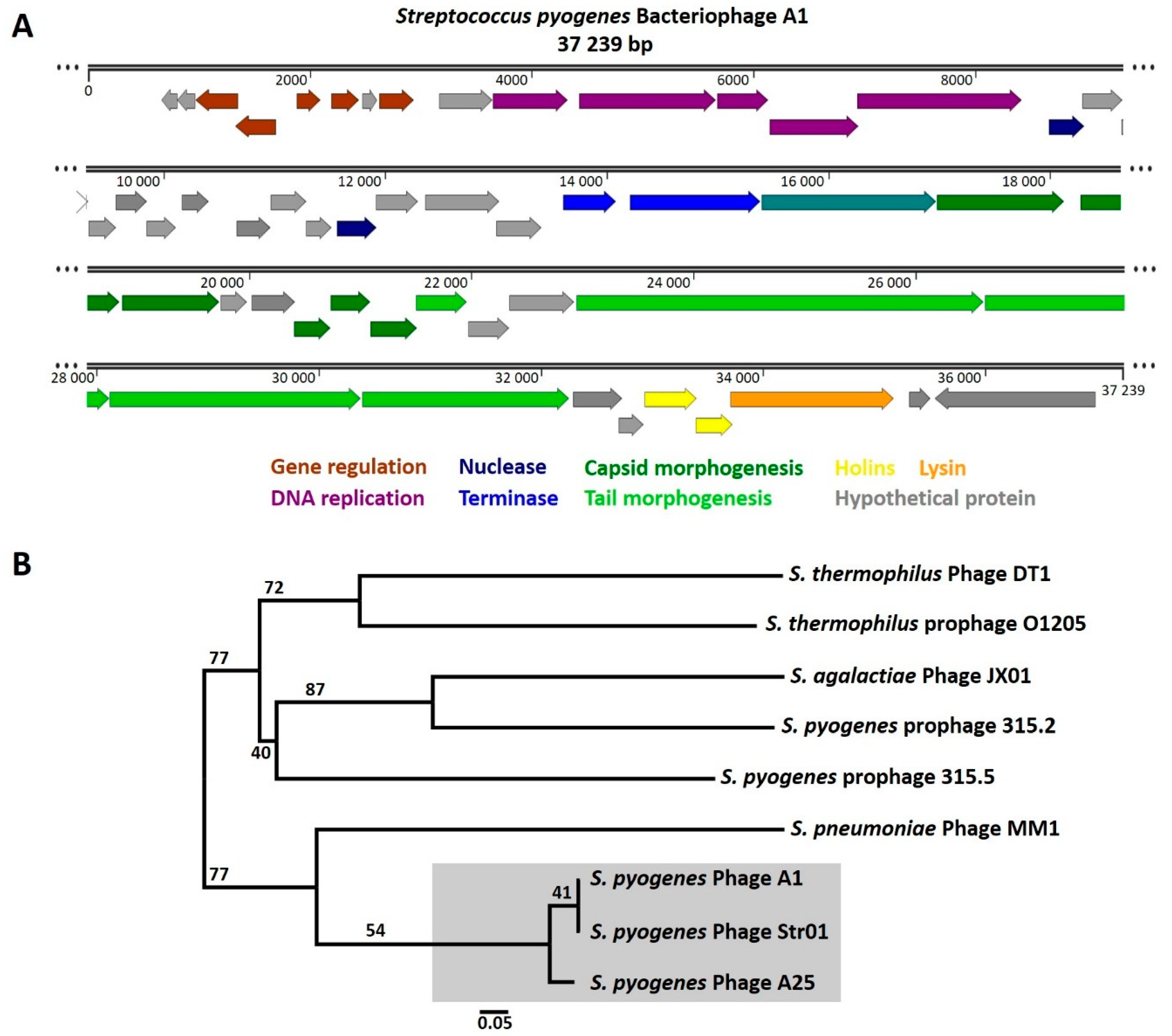
3.2. Genome Sequence and Phylogeny of S. pyogenes Bacteriophage A1
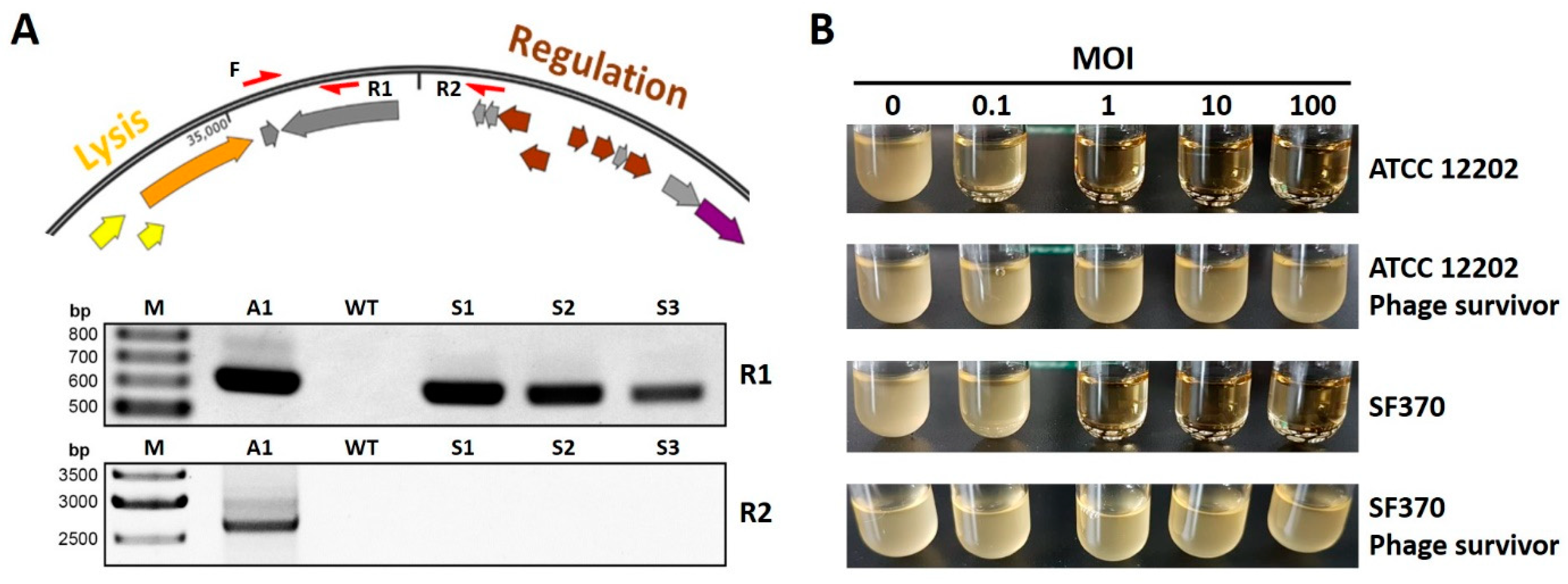
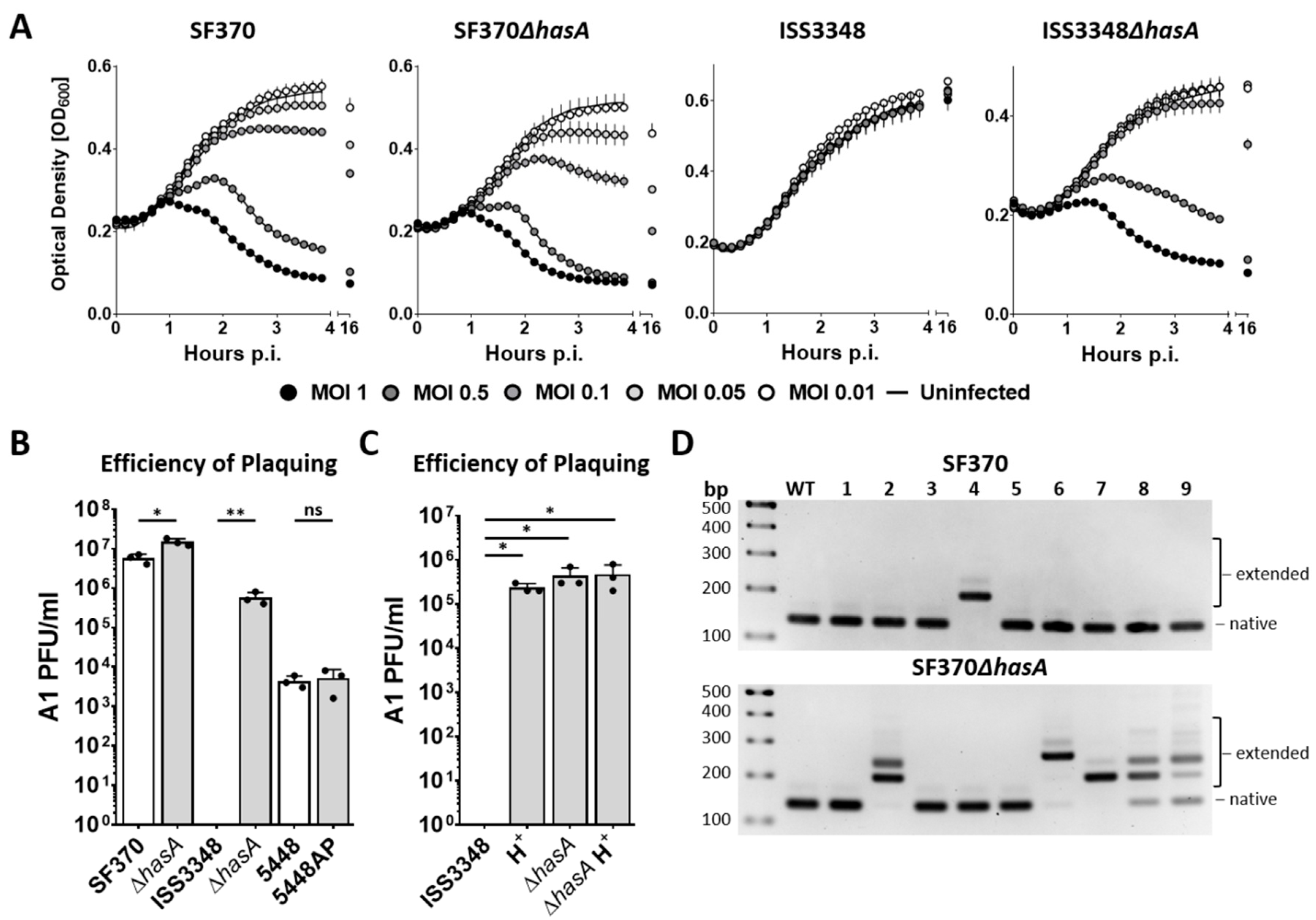
3.3. Phage A1 Is Potentially Temperate and Phage A1 Survivors Acquire Phage Resistance
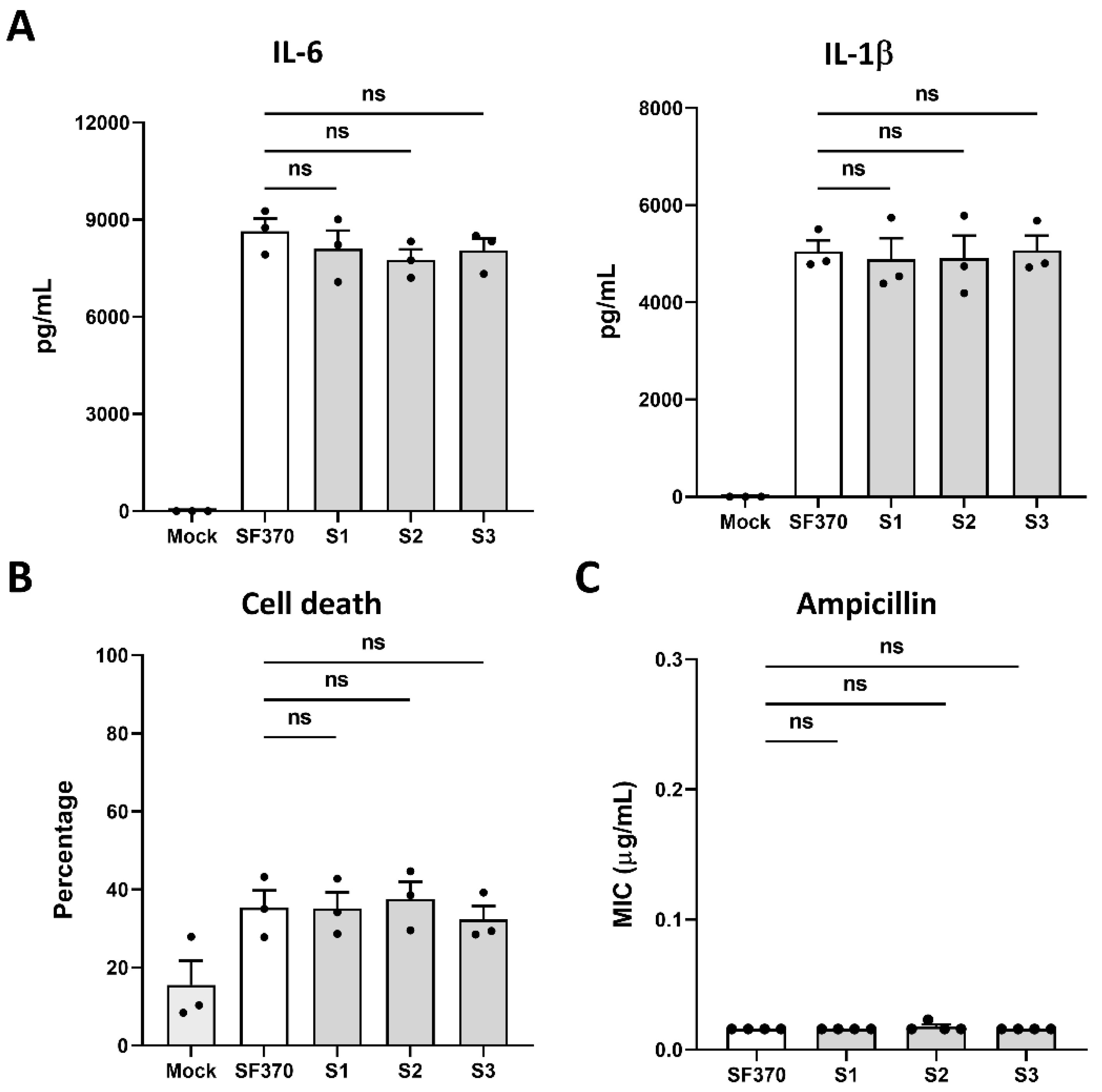
3.4. Phage A1-Surviving Strains Do Not Alter the Host Immune Response or Tolerance to Antibiotics
3.5. CRISPR-Cas-Mediated Spacer Acquisition Is Increased in the Absence of a Capsule
3.6. Membrane Vesicles of S. pyogenes Bind Phages and Limit Phage Infection
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ferretti, J.; Köhler, W. History of Streptococcal Research; University of Oklahoma Health Sciences Center: Oklahoma City, OK, USA, 2016. [Google Scholar]
- Vela, A.I.; Villalón, P.; Sáez-Nieto, J.A.; Chacon, G.; Dominguez, L.; Fernández-Garayzábal, J.F. Characterization of Streptococcus pyogenes from Animal Clinical Specimens, Spain. Emerg. Infect. Dis. 2017, 23, 2013–2016. [Google Scholar] [CrossRef] [Green Version]
- Martin, J. The Streptococcus pyogenes Carrier State; University of Oklahoma Health Sciences Center: Oklahoma City, OK, USA, 2016. [Google Scholar]
- Cunningham, M.W. Pathogenesis of group a streptococcal infections. Clin. Microbiol. Rev. 2000, 13, 470–511. [Google Scholar] [CrossRef]
- Henningham, A.; Barnett, T.C.; Maamary, P.G.; Walker, M.J. Pathogenesis of group A streptococcal infections. Discov. Med. 2012, 13, 329–342. [Google Scholar]
- Carapetis, J.R.; Steer, A.C.; Mulholland, E.K.; Weber, M. The global burden of group A streptococcal diseases. Lancet Infect. Dis. 2005, 5, 685–694. [Google Scholar] [CrossRef]
- Cattoir, V. Mechanisms of Antibiotic Resistance. In Virulence Mechanisms of Bacterial Pathogens; American Society for Microbiology: Washington, DC, USA, 2016; pp. 481–511. [Google Scholar]
- Brook, I. Penicillin Failure in the Treatment of Streptococcal Pharyngo-Tonsillitis. Curr. Infect. Dis. Rep. 2013, 15, 232–235. [Google Scholar] [CrossRef] [PubMed]
- Cisek, A.A.; Dąbrowska, I.; Gregorczyk, K.P.; Wyżewski, Z. Phage Therapy in Bacterial Infections Treatment: One Hundred Years After the Discovery of Bacteriophages. Curr. Microbiol. 2017, 74, 277–283. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koskella, B.; Meaden, S. Understanding Bacteriophage Specificity in Natural Microbial Communities. Viruses 2013, 5, 806–823. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fischetti, V.A. Streptococcal M protein: Molecular design and biological behavior. Clin. Microbiol. Rev. 1989, 2, 285–314. [Google Scholar] [CrossRef]
- Maxted, W.R. The indirect bactericidal test as a means of identifying antibody to the M antigen of Streptococcus pyogenes. Br. J. Exp. Pathol. 1956, 37, 415–422. [Google Scholar] [PubMed]
- Perez-Casal, J.; Caparon, M.; Scott, J. Introduction of the emm6 gene into an emm-deleted strain of Streptococcus pyogenes restores its ability to resist phagocytosis. Res. Microbiol. 1992, 143, 549–558. [Google Scholar] [CrossRef]
- Cleary, P.P.; Johnson, Z. Possible dual function of M protein: Resistance to bacteriophage A25 and resistance to phagocytosis by human leukocytes. Infect. Immun. 1977, 16, 280–292. [Google Scholar] [CrossRef] [Green Version]
- Wessels, M.R. Cell Wall and Surface Molecules: Capsule; University of Oklahoma Health Sciences Center: Oklahoma City, OK, USA, 2016. [Google Scholar]
- Ashbaugh, C.D.; Warren, H.B.; Carey, V.J.; Wessels, M.R. Molecular analysis of the role of the group A streptococcal cysteine protease, hyaluronic acid capsule, and M protein in a murine model of human invasive soft-tissue infection. J. Clin. Investig. 1998, 102, 550–560. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wessels, M.R.; Moses, A.E.; Goldberg, J.B.; Dicesare, T.J. Hyaluronic acid capsule is a virulence factor for mucoid group A streptococci. Proc. Natl. Acad. Sci. USA 1991, 88, 8317–8321. [Google Scholar] [CrossRef] [Green Version]
- Maxted, W.R. Enhancement of Streptococcal Bacteriophage Lysis by Hyaluronidase. Nat. Cell Biol. 1952, 170, 1020–1021. [Google Scholar] [CrossRef]
- DeAngelis, P.; Papaconstantinou, J.; Weigel, P. Isolation of a Streptococcus pyogenes gene locus that directs hyaluronan biosynthesis in acapsular mutants and in heterologous bacteria. J. Biol. Chem. 1993, 268, 14568–14571. [Google Scholar] [CrossRef]
- Dougherty, B.A.; Van De Rijn, I. Molecular characterization of hasA from an operon required for hyaluronic acid synthesis in group A streptococci. J. Biol. Chem. 1994, 269, 169–175. [Google Scholar] [CrossRef]
- Heath, A.; DiRita, V.J.; Barg, N.L.; Engleberg, N.C. A Two-Component Regulatory System, CsrR-CsrS, Represses Expression of Three Streptococcus pyogenes Virulence Factors, Hyaluronic Acid Capsule, Streptolysin S, and Pyrogenic Exotoxin B. Infect. Immun. 1999, 67, 5298–5305. [Google Scholar] [CrossRef] [Green Version]
- Fiebig, A.; Loof, T.G.; Babbar, A.; Itzek, A.; Koehorst, J.J.; Schaap, P.J.; Nitsche-Schmitz, D.P. Comparative Genomics of Streptococcus pyogenes M1 isolates differing in virulence and propensity to cause systemic infection in mice. Int. J. Med Microbiol. 2015, 305, 532–543. [Google Scholar] [CrossRef]
- McShan, W.M.; McCullor, K.A.; Nguyen, S.V. The Bacteriophages of Streptococcus pyogenes. Microbiol. Spectr. 2019, 7, 158–176. [Google Scholar] [CrossRef] [PubMed]
- Maxted, B.W.R. The Influence of Bacteriophage on Streptococcus pyogenes. Microbiology 1955, 12, 484–495. [Google Scholar] [CrossRef] [Green Version]
- Wannamaker, L.W.; Almquist, S.; Skjold, S. Intergroup phage reactions and transduction between group C and group A streptococci. J. Exp. Med. 1973, 137, 1338–1353. [Google Scholar] [CrossRef] [PubMed]
- Malke, H. Characteristics of transducing group A streptococcal bacteriophages A5 and A25. Arch. Virol. 1970, 29, 44–49. [Google Scholar] [CrossRef]
- Malke, H. Transduction of Streptococcus pyogenes K 56 by Temperature-Sensitive Mutants of the Transducing Phage A25. Z. Nat. B 1969, 24, 1556–1561. [Google Scholar] [CrossRef]
- Fischetti, V.A.; Barron, B.A.; Zabriskie, J.B. Studies on streptococcal bacteriophages. J. Exp. Med. 1968, 127, 475–488. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cleary, P.P.; Wannamaker, L.W.; Fisher, M.; Laible, N. Studies of the receptor for phage A25 in group A streptococci: The role of peptidoglycan in reversible adsorption. J. Exp. Med. 1977, 145, 578–593. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McCullor, K.; Postoak, B.; Rahman, M.; King, C.; McShan, W.M. Genomic Sequencing of High-Efficiency Transducing Streptococcal Bacteriophage A25: Consequences of Escape from Lysogeny. J. Bacteriol. 2018, 200, e00358-18. [Google Scholar] [CrossRef] [Green Version]
- Ferretti, J.J.; McShan, W.M.; Ajdic, D.; Savic, D.J.; Savic, G.; Lyon, K.; Primeaux, C.; Sezate, S.; Suvorov, A.N.; Kenton, S.; et al. Complete genome sequence of an M1 strain of Streptococcus pyogenes. Proc. Natl. Acad. Sci. USA 2001, 98, 4658–4663. [Google Scholar] [CrossRef] [Green Version]
- Abbot, E.L.; Smith, W.D.; Siou, G.P.S.; Chiriboga, C.; Smith, R.J.; Wilson, J.A.; Hirst, B.H.; Kehoe, M.A. Pili mediate specific adhesion of Streptococcus pyogenes to human tonsil and skin. Cell. Microbiol. 2007, 9, 1822–1833. [Google Scholar] [CrossRef]
- Kazmi, S.U.; Kansal, R.; Aziz, R.K.; Hooshdaran, M.; Norrby-Teglund, A.; Low, D.E.; Halim, A.-B.; Kotb, M. Reciprocal, Temporal Expression of SpeA and SpeB by Invasive M1T1 Group A Streptococcal Isolates In Vivo. Infect. Immun. 2001, 69, 4988–4995. [Google Scholar] [CrossRef] [Green Version]
- Aziz, R.K.; Pabst, M.J.; Jeng, A.; Kansal, R.; Low, D.E.; Nizet, V.; Kotb, M. Invasive M1T1 group A Streptococcus undergoes a phase-shift in vivo to prevent proteolytic degradation of multiple virulence factors by SpeB. Mol. Microbiol. 2003, 51, 123–134. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Resch, U.; Tsatsaronis, J.A.; Le Rhun, A.; Stuebiger, G.; Rohde, M.; Kasvandik, S.; Holzmeister, S.; Tinnefeld, P.; Wai, S.N.; Charpentier, E. A Two-Component Regulatory System Impacts Extracellular Membrane-Derived Vesicle Production in Group A Streptococcus. mBio 2016, 7, e00207-16. [Google Scholar] [CrossRef] [Green Version]
- Kropinski, A.M. Practical Advice on the One-Step Growth Curve. In Methods in Molecular Biology; Springer Science and Business Media LLC: Berlin, Germany, 2018; Volume 1681, pp. 41–47. [Google Scholar]
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A flexible trimmer for Illumina sequence data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef] [Green Version]
- Bankevich, A.; Nurk, S.; Antipov, D.; Gurevich, A.A.; Dvorkin, M.; Kulikov, A.S.; Lesin, V.M.; Nikolenko, S.I.; Pham, S.; Prjibelski, A.D.; et al. SPAdes: A New Genome Assembly Algorithm and Its Applications to Single-Cell Sequencing. J. Comput. Biol. 2012, 19, 455–477. [Google Scholar] [CrossRef] [Green Version]
- Seemann, T. Prokka: Rapid Prokaryotic Genome Annotation. Bioinformatics 2014, 30, 2068–2069. [Google Scholar] [CrossRef] [PubMed]
- Meier-Kolthoff, J.P.; Göker, M. VICTOR: Genome-based phylogeny and classification of prokaryotic viruses. Bioinformations 2017, 33, 3396–3404. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rapino, F.; Robles, E.F.; Richter-Larrea, J.A.; Kallin, E.M.; Martinez-Climent, J.A.; Graf, T. C/EBPα Induces Highly Efficient Macrophage Transdifferentiation of B Lymphoma and Leukemia Cell Lines and Impairs Their Tumorigenicity. Cell Rep. 2013, 3, 1153–1163. [Google Scholar] [CrossRef] [Green Version]
- Athukoralage, J.S.; McMahon, S.A.; Zhang, C.; Grüschow, S.; Graham, S.; Krupovic, M.; Whitaker, R.J.; Gloster, T.M.; White, M.F. An anti-CRISPR viral ring nuclease subverts type III CRISPR immunity. Nat. Cell Biol. 2020, 577, 572–575. [Google Scholar] [CrossRef]
- Dale, D.C.; Boxer, L.; Liles, W.C. The phagocytes: Neutrophils and monocytes. Blood 2008, 112, 935–945. [Google Scholar] [CrossRef] [PubMed]
- Banks, D.J.; Porcella, S.F.; Barbian, K.D.; Martin, J.M.; Musser, J.M. Structure and Distribution of an Unusual Chimeric Genetic Element Encoding Macrolide Resistance in Phylogenetically Diverse Clones of Group A Streptococcus. J. Infect. Dis. 2003, 188, 1898–1908. [Google Scholar] [CrossRef] [PubMed]
- Alberti, S.; Ashbaugh, C.D.; Wessels, M.R. Structure of the has operon promoter and regulation of hyaluronic acid capsule expression in group A Streptococcus. Mol. Microbiol. 1998, 28, 343–353. [Google Scholar] [CrossRef]
- Graham, M.R.; Smoot, L.M.; Migliaccio, C.A.L.; Virtaneva, K.; Sturdevant, D.E.; Porcella, S.F.; Federle, M.J.; Adams, G.J.; Scott, J.R.; Musser, J.M. Virulence control in group A Streptococcus by a two-component gene regulatory system: Global expression profiling and in vivo infection modeling. Proc. Natl. Acad. Sci. USA 2002, 99, 13855–13860. [Google Scholar] [CrossRef] [Green Version]
- Heler, R.; Samai, P.; Modell, J.W.; Weiner, C.; Goldberg, G.W.; Bikard, D.; Marraffini, L.A. Cas9 specifies functional viral targets during CRISPR–Cas adaptation. Nat. Cell Biol. 2015, 519, 199–202. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Manning, A.J.; Kuehn, M.J. Contribution of bacterial outer membrane vesicles to innate bacterial defense. BMC Microbiol. 2011, 11, 258. [Google Scholar] [CrossRef] [Green Version]
- Dunne, M.; Hupfeld, M.; Klumpp, J.; Loessner, M.J. Molecular Basis of Bacterial Host Interactions by Gram-Positive Targeting Bacteriophages. Viruses 2018, 10, 397. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Harhala, M.; Barylski, J.; Humińska-Lisowska, K.; Lecion, D.; Wojciechowicz, J.; Lahutta, K.; Kuś, M.; Kropinski, A.M.; Nowak, S.; Nowicki, G.; et al. Two novel temperate bacteriophages infecting Streptococcus pyogenes: Their genomes, morphology and stability. PLoS ONE 2018, 13, e0205995. [Google Scholar] [CrossRef]
- Broussard, G.W.; Oldfield, L.M.; Villanueva, V.M.; Lunt, B.L.; Shine, E.E.; Hatfull, G.F. Integration-Dependent Bacteriophage Immunity Provides Insights into the Evolution of Genetic Switches. Mol. Cell 2013, 49, 237–248. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kaplan, E.L.; Wotton, J.T.; Johnson, D.R. Dynamic epidemiology of group A streptococcal serotypes associated with pharyngitis. Lancet 2001, 358, 1334–1337. [Google Scholar] [CrossRef]
- Ikebe, T.; Wada, A.; Inagaki, Y.; Sugama, K.; Suzuki, R.; Tanaka, D.; Tamaru, A.; Fujinaga, Y.; Abe, Y.; Shimizu, Y.; et al. Dissemination of the Phage-Associated Novel Superantigen Gene speL in Recent Invasive and Noninvasive Streptococcus pyogenes M3/T3 Isolates in Japan. Infect. Immun. 2002, 70, 3227–3233. [Google Scholar] [CrossRef] [Green Version]
- Banks, D.J.; Lei, B.; Musser, J.M. Prophage Induction and Expression of Prophage-EncodedVirulence Factors in Group A Streptococcus Serotype M3 StrainMGAS315. Infect. Immun. 2003, 71, 7079–7086. [Google Scholar] [CrossRef] [Green Version]
- Bondy-Denomy, J.; Qian, J.; Westra, E.R.; Buckling, A.; Guttman, D.S.; Davidson, A.R.; Maxwell, K.L. Prophages mediate defense against phage infection through diverse mechanisms. ISME J. 2016, 10, 2854–2866. [Google Scholar] [CrossRef]
- Vojtek, I.; Pirzada, Z.A.; Henriques-Normark, B.; Mastny, M.; Janapatla, R.P.; Charpentier, E. Lysogenic Transfer of Group AStreptococcusSuperantigen Gene among Streptococci. J. Infect. Dis. 2008, 197, 225–234. [Google Scholar] [CrossRef] [Green Version]
- Weeks, C.R.; Ferretti, J.J. The gene for type a streptococcal exotoxin (erythrogenic toxin) is located in bacteriophage T12. Infect. Immun. 1984, 46, 531–536. [Google Scholar] [CrossRef] [Green Version]
- Monteiro, R.; Pires, D.P.; Costa, A.R.; Azeredo, J. Phage Therapy: Going Temperate? Trends Microbiol. 2019, 27, 368–378. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Park, J.Y.; Moon, B.Y.; Park, J.W.; Thornton, J.A.; Park, Y.H.; Seo, K.S. Genetic engineering of a temperate phage-based delivery system for CRISPR/Cas9 antimicrobials against Staphylococcus aureus. Sci. Rep. 2017, 7, srep44929. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fischetti, V.A. Development of Phage Lysins as Novel Therapeutics: A Historical Perspective. Viruses 2018, 10, 310. [Google Scholar] [CrossRef] [Green Version]
- Dugar, G.; Herbig, A.; Förstner, K.U.; Heidrich, N.; Reinhardt, R.; Nieselt, K.; Sharma, C.M. High-Resolution Transcriptome Maps Reveal Strain-Specific Regulatory Features of Multiple Campylobacter jejuni Isolates. PLoS Genet. 2013, 9, e1003495. [Google Scholar] [CrossRef] [PubMed]
- Dugar, G.; Leenay, R.T.; Eisenbart, S.K.; Bischler, T.; Aul, B.U.; Beisel, C.L.; Sharma, C.M. CRISPR RNA-Dependent Binding and Cleavage of Endogenous RNAs by the Campylobacter jejuni Cas9. Mol. Cell 2018, 69, 893–905.e7. [Google Scholar] [CrossRef] [Green Version]
- Sacher, J.C.; Flint, A.; Butcher, J.; Blasdel, B.; Reynolds, H.M.; Lavigne, R.; Stintzi, A.; Szymanski, C.M. Transcriptomic Analysis of the Campylobacter jejuni Response to T4-Like Phage NCTC 12673 Infection. Viruses 2018, 10, 332. [Google Scholar] [CrossRef] [Green Version]
- Heler, R.; Wright, A.V.; Vucelja, M.; Doudna, J.A.; Marraffini, L.A. Spacer Acquisition Rates Determine the Immunological Diversity of the Type II CRISPR-Cas Immune Response. Cell Host Microbe 2019, 25, 242–249.e3. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nuñez, J.K.; Bai, L.; Harrington, L.B.; Hinder, T.L.; Doudna, J.A. CRISPR Immunological Memory Requires a Host Factor for Specificity. Mol. Cell 2016, 62, 824–833. [Google Scholar] [CrossRef] [Green Version]
- Tsatsaronis, J.A.; Franch-Arroyo, S.; Resch, U.; Charpentier, E. Extracellular Vesicle RNA: A Universal Mediator of Microbial Communication? Trends Microbiol. 2018, 26, 401–410. [Google Scholar] [CrossRef] [PubMed]
- Reyes-Robles, T.; Dillard, R.S.; Cairns, L.S.; Silva-Valenzuela, C.A.; Housman, M.; Ali, A.; Wright, E.R.; Camilli, A. Vibrio cholerae Outer Membrane Vesicles Inhibit Bacteriophage Infection. J. Bacteriol. 2018, 200. [Google Scholar] [CrossRef] [PubMed] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strain | Description | Origin |
|---|---|---|
| ATCC 12202 | Streptococcus pyogenes Rosenbach (ATCC® 12202™) | ATCC database |
| SF370 | Streptococcus pyogenes Rosenbach (ATCC® 700294™) | Ferretti et al. 2001 [31] and Abbot et al. 2007 [32] |
| SF370ΔhasA | SF370 with complete deletion of the hasA coding sequence | Abbot et al. 2007 [32] |
| 5448 | Wild-type clinical STSS/Necrotizing Fasciitis isolate | M. Walker, University of Queensland, Australia [33] |
| 5448AP | Animal passaged variant of 5448 containing a 1-bp insertion in covS | M. Walker, University of Queensland, Australia [34] |
| ISS3348 | Clinical isolate; 30-bp deletion in covS | G. Teti, University of Messina, Italy |
| 3348ΔhasA | ISS3348 with complete deletion of the hasA coding sequence | Resch et al. 2016 [35] |
| Phage Parameter | Bacteriophage A1 (nm ± SD *) |
|---|---|
| Head | 60 ± 2 |
| Tail length | 187 ± 4 |
| Tail width | 10 ± 1 |
| Baseplate width | 16 ± 2 |
| Total length | 249 ± 3 |
| Genomic Location (Gp Number) | Identified Protein (Species) | Direction | Accession Number | E Value | Identity % |
|---|---|---|---|---|---|
| 667..801 (Gp01) | Hypothetical phage protein (S. pyogenes Phage 10270.2) | <= | ABF33863.1 | 1e−21 | 100 |
| 808..960 (Gp02) | Hypothetical protein (S. pyogenes) | <= | WP_011054825.1 | 5e−27 | 100 |
| 971..1348 (Gp03) | ImmA/IrrE family metallo-endopeptidase (S. pyogenes) | <= | WP_011054824.1 | 2e−86 | 100 |
| 1332..1691 (Gp04) | XRE family transcriptional regulator (S. pyogenes Phage Str01) | <= | APZ81910.1 | 2e−79 | 100 |
| 1881..2099 (Gp05) | Cro family anti-repressor (S. pyogenes Phage 10270.2) | => | ABF33867.1 | 2e−44 | 100 |
| 2194..2445 (Gp06) | Putative DNA-binding antitoxin (S. pyogenes GA40634) | => | EPZ46884.1 | 4e−46 | 98.65 |
| 2476..2610 (Gp07) | Hypothetical protein (S. pyogenes Phage 10270.2) | => | ABF33611.1 | 5e−24 | 100 |
| 2626..2940 (Gp08) | Helix-turn-helix transcriptional regulator (S. pyogenes) | => | WP_011528544.1 | 2e−69 | 100 |
| 3167..3649 (Gp09) | Hypothetical Gp157 family protein (S. pyogenes) | => | WP_011528545.1 | 2e−108 | 100 |
| 3650..4330 (Gp10) | AAA family ATPase (S. pyogenes MGAS1882) | => | AFC68359.1 | 2e−164 | 99.56 |
| 4432..5661 (Gp11) | DEAD/DEAH box helicase (Streptococcus sp.) | => | WP_011528546.1 | 0 | 100 |
| 5677..6135 (Gp12) | DUF669 domain-containing protein (Streptococcus sp.) | => | WP_002995969.1 | 2e−107 | 100 |
| 6147..6950 (Gp13) | Bifunctional DNA primase/polymerase (S. pyogenes) | => | WP_011528547.1 | 0 | 99.25 |
| 6940..8421 (Gp14) | DNA primase (S. pyogenes) | => | WP_020905118.1 | 0 | 99.8 |
| 8666..8986 (Gp15) | VRR-NUC domain-containing protein (S. pyogenes) | => | WP_002995960.1 | 7e−71 | 100 |
| 8970..9326 (Gp16) | Hypothetical protein (S. pyogenes) | => | WP_011018138.1 | 4e−79 | 100 |
| 9323..9574 (Gp17) | Hypothetical protein (S. pyogenes) | => | WP_011528549.1 | 4e−54 | 100 |
| 9568..9852 (Gp18) | Hypothetical DUF3310-containing protein (S. pyogenes) | => | WP_011017568.1 | 5e−64 | 100 |
| 9849..10,118 (Gp19) | Hypothetical protein (S. dysgalactiae) | => | SUN67275.1 | 1e−58 | 98.88 |
| 10,170..10,409 (Gp20) | Hypothetical TIGR01671 family protein (S. pyogenes GA40634) | => | ESA49192.1 | 6e−33 | 98.28 |
| 10,660..10,971 (Gp21) | Hypothetical DUF1372-containing protein (S. dysgalactiae) | => | WP_081281150.1 | 9e−62 | 92.31 |
| 10,971..11,294 (Gp22) | Hypothetical protein (S. pyogenes Phage Str01) | => | APZ81913.1 | 2e−70 | 100 |
| 11,287..11,517 (Gp23) | Hypothetical protein (S. pyogenes Phage A25) | => | YP_009191526.1 | 2e−47 | 100 |
| 11,565..11,924 (Gp24) | Endodeoxyribonuclease RusA (S. dysgalactiae) | => | WP_065359284.1 | 7e−81 | 97.48 |
| 11,917..12,297 (Gp25) | Hypothetical protein (S. pyogenes Phage Str01) | => | APZ81908.1 | 7e−86 | 100 |
| 12,364..13,035 (Gp26) | Hypothetical DUF4417-containing protein (S. dysgalactiae) | => | WP_155778398.1 | 3e−164 | 98.21 |
| 13,005..13,415 (Gp27) | Hypothetical protein (S. pyogenes Phage Str01) | => | APZ81907.1 | 2e−87 | 100 |
| 13,608..14,087 (Gp28) | Terminase (small subunit) (S. dysgalactiae) | => | WP_003058573.1 | 6e−111 | 100 |
| 14,209..15,387 (Gp29) | Terminase (large subunit) (S. dysgalactiae) | => | WP_003058577.1 | 0 | 100 |
| 15,399..16,976 (Gp30) | Portal protein (S. dysgalactiae) | => | WP_003058556.1 | 0 | 100 |
| 16,979..18,127 (Gp31) | Minor capsid protein (S. dysgalactiae) | => | WP_003058578.1 | 0 | 100 |
| 18,274..18,837 (Gp32) | Scaffolding protein (S. pyogenes Phage A25) | => | YP_009191535.1 | 2e−129 | 100 |
| 18,856..19,734 (Gp33) | Major capsid protein (Streptococcus canis) | => | WP_164406187.1 | 0 | 97.26 |
| 19,745..19,981 (Gp34) | Hypothetical protein (S. dysgalactiae) | => | WP_003058596.1 | 3e−46 | 100 |
| 20,025..20,417 (Gp35) | Hypothetical protein (S. dysgalactiae) | => | WP_003058548.1 | 9e−91 | 100 |
| 20,407..20,733 (Gp36) | Minor capsid protein (S. pyogenes Phage Str01) | => | APZ81883.1 | 8e−70 | 100 |
| 20,733..21,092 (Gp37) | Capsid protein (S. pyogenes Phage A25) | => | YP_009191540.1 | 4e−82 | 100 |
| 21,092..21,511 (Gp38) | Capsid protein (S. dysgalactiae) | => | WP_155783005.1 | 4e−96 | 100 |
| 21,504..21,962 (Gp39) | Major tail shaft protein (S. pyogenes Phage A25) | => | YP_009191542.1 | 6e−107 | 100 |
| 21,977..22,345 (Gp40) | Hypothetical protein (S. dysgalactiae) | => | WP_155783003.1 | 1e−81 | 100 |
| 22,345..22,929 (Gp41) | Hypothetical Gp15 protein (Streptococcus equi subsp. zooepidemicus) | => | KIS17966.1 | 4e−88 | 68.39% |
| 22,946..26,617 (Gp42) | Tail tape measure protein (S. pyogenes Phage Str01) | => | APZ81886.1 | 0 | 100 |
| 26,626..28,116 (Gp43) | Tail endopeptidase (S. pyogenes Phage Str01) | => | APZ81874.1 | 0 | 100 |
| 28,120..30,381 (Gp44) | Minor structural tail protein (S. pyogenes Phage A25) | => | ALF02720.1 | 0 | 100 |
| 30,393..32,258 (Gp45) | Structural tail protein (S. pyogenes Phage A25) | => | ALF02721.1 | 0 | 100 |
| 32,291..32,737 (Gp46) | Hypothetical DUF1366-containing protein (S. dysgalactiae) | => | WP_003058584.1 | 5e−97 | 99.3 |
| 32,703..32,930 (Gp47) | Hypothetical protein (S. pyogenes Phage Str01) | => | APZ81920.1 | 4e−45 | 100 |
| 32,938..33,405 (Gp48) | Class I Holin (S. pyogenes Phage A25) | => | YP_009191551.1 | 2e−106 | 98.71 |
| 33,398..33,733 (Gp49) | Class III Holin (S. dysgalactiae) | => | WP_003058565.1 | 6e−74 | 100 |
| 33,708..35,180 (Gp50) | Lysin (S. pyogenes Phage A25) | => | YP_009191553.1 | 0 | 100 |
| 35,318..35,509 (Gp51) | Hypothetical Paratox protein (S. pyogenes) | => | WP_136110174.1 | 2e−06 | 51.22 |
| 35,552..36,991 (Gp52) | Hypothetical Recombinase family protein (S. bovimastitidis) | <= | WP_071794279.1 | 0 | 82.88 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Beerens, D.; Franch-Arroyo, S.; Sullivan, T.J.; Goosmann, C.; Brinkmann, V.; Charpentier, E. Survival Strategies of Streptococcus pyogenes in Response to Phage Infection. Viruses 2021, 13, 612. https://doi.org/10.3390/v13040612
Beerens D, Franch-Arroyo S, Sullivan TJ, Goosmann C, Brinkmann V, Charpentier E. Survival Strategies of Streptococcus pyogenes in Response to Phage Infection. Viruses. 2021; 13(4):612. https://doi.org/10.3390/v13040612
Chicago/Turabian StyleBeerens, Dior, Sandra Franch-Arroyo, Timothy J. Sullivan, Christian Goosmann, Volker Brinkmann, and Emmanuelle Charpentier. 2021. "Survival Strategies of Streptococcus pyogenes in Response to Phage Infection" Viruses 13, no. 4: 612. https://doi.org/10.3390/v13040612