
Prevalence and Phylogenetic Analysis of Parvovirus (B19V) among Blood Donors with Different Nationalities Residing in Qatar

 , , ,
, , , 
 , and
, and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Ethical Approval and Sample Collection
2.2. DNA Extraction
2.3. B19V DNA Detection by Quantitative Real-Time PCR
2.4. Qualitative ELISA Testing of B19V Antibodies
2.5. Immune Status Classification of Blood Donors
2.6. Amplification of the NS1-VP1u Region by Semi-Nested PCR
2.7. Sanger Sequencing of the NS1-VP1u Region
2.8. Phylogenetic Analysis
2.9. Statistical Analysis
3. Results
3.1. Demographic Characteristics of the Donors
3.2. B19V Viremia Rates among Healthy Blood Donors
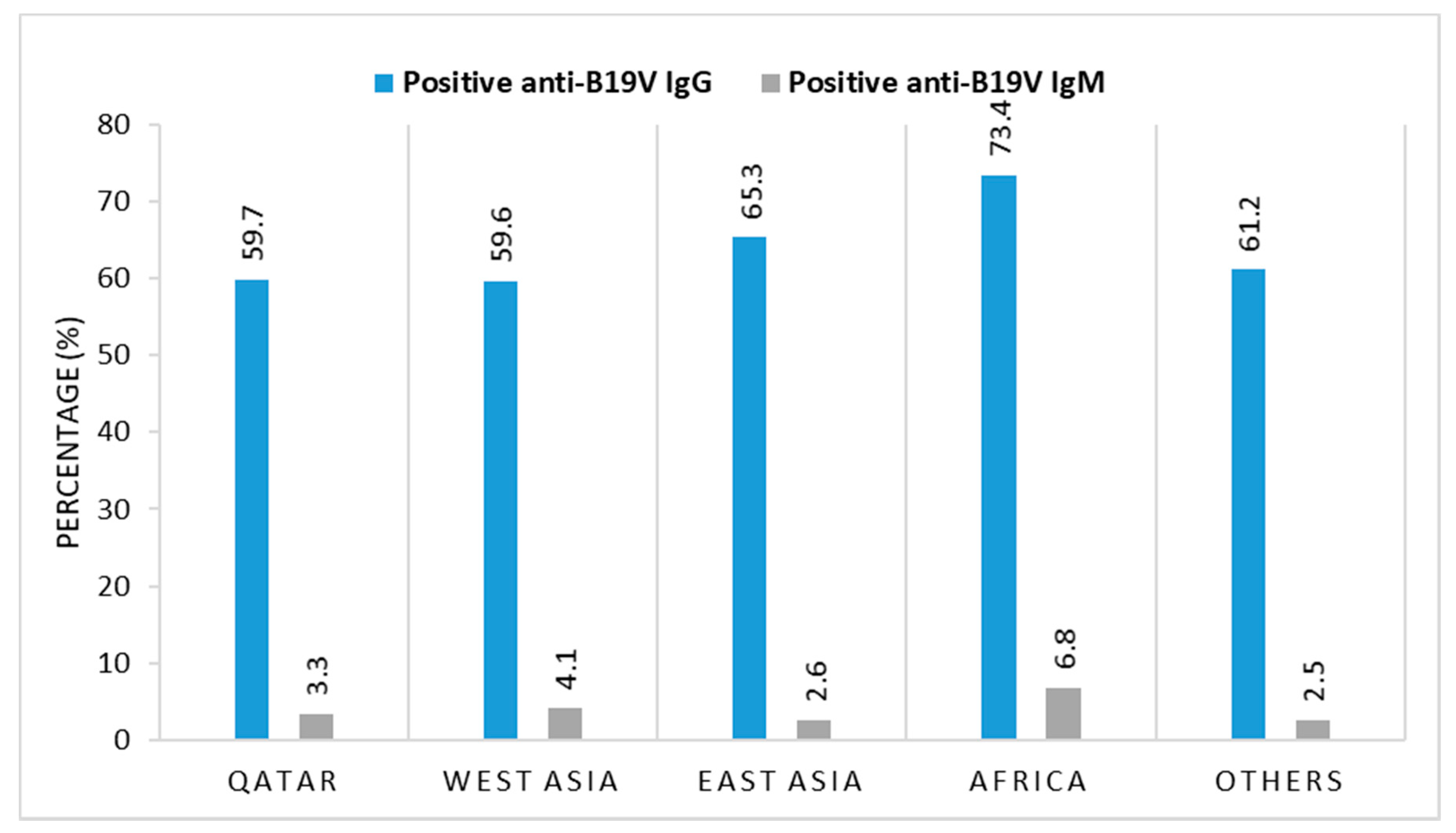
3.3. B19V Seroprevalence and Correlation with Gender, Geographic Origin, and Age
3.4. Identifying the B19V Immune Status of the Donors
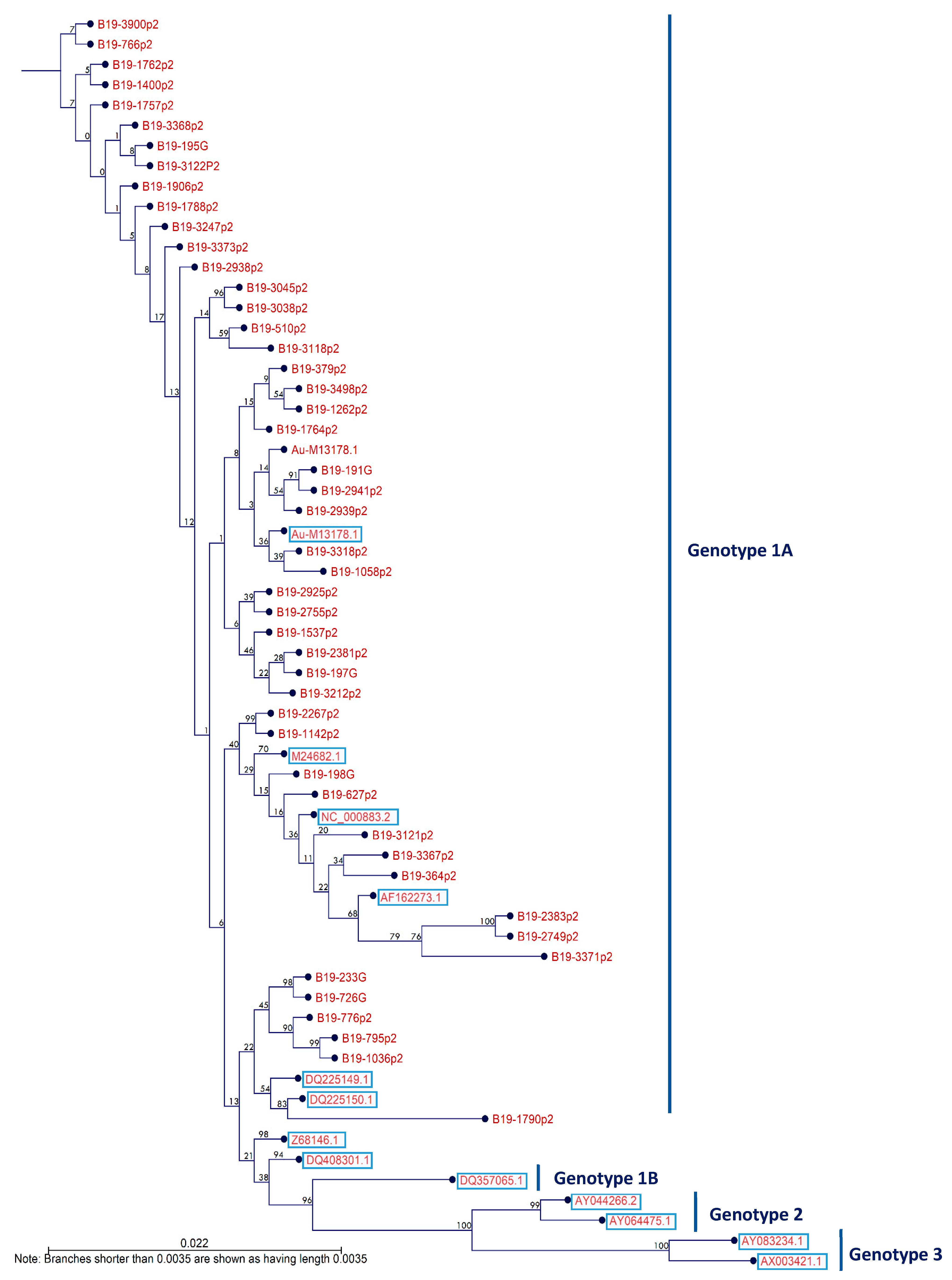
3.5. Circulating B19V Genotypes among Blood Donors
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviation
| B19 | Human parvovirus (B19) |
| EDTA | Ethylenediamine tetraacetic acid |
| ELISA | Enzyme-linked immunosorbent assay |
| HMC | Hamad Medical Corporation |
| IRB | Institutional review board |
| MENA | Middle East and North Africa |
| qQPCR | Quantitave real-time polymerase chain reaction |
| NS | Nonstructural protein |
References
- Servant, A.; Laperche, S.; Lallemand, F.; Marinho, V.; De Saint Maur, G.; Meritet, J.F.; Garbarg-Chenon, A. Genetic diversity within human erythroviruses: Identification of three genotypes. J. Virol. 2002, 76, 9124–9134. [Google Scholar] [CrossRef]
- Heegaard, E.D.; Brown, K.E. Human parvovirus B19. Clin. Microbiol. Rev. 2002, 15, 485–505. [Google Scholar] [CrossRef] [PubMed]
- Obeid, O.E. Molecular and serological assessment of parvovirus B19 infections among sickle cell anemia patients. J. Infect. Dev. Ctries. 2011, 5, 535–539. [Google Scholar] [CrossRef]
- Anderson, M.J.; Higgins, P.G.; Davis, L.R.; Willman, J.S.; Jones, S.E.; Kidd, I.M.; Pattison, J.R.; Tyrrell, D.A. Experimental parvoviral infection in humans. J. Infect. Dis. 1985, 152, 257–265. [Google Scholar] [CrossRef] [PubMed]
- Woolf, A.D.; Campion, G.V.; Chishick, A.; Wise, S.; Cohen, B.J.; Klouda, P.T.; Caul, O.; Dieppe, P.A. Clinical manifestations of human parvovirus B19 in adults. Arch. Intern. Med. 1989, 149, 1153–1156. [Google Scholar] [CrossRef]
- Toppinen, M.; Norja, P.; Aaltonen, L.M.; Wessberg, S.; Hedman, L.; Söderlund-Venermo, M.; Hedman, K. A new quantitative PCR for human parvovirus B19 genotypes. J. Virol. Methods 2015, 218, 40–45. [Google Scholar] [CrossRef]
- Ekman, A.; Hokynar, K.; Kakkola, L.; Kantola, K.; Hedman, L.; Bondén, H.; Gessner, M.; Aberham, C.; Norja, P.; Miettinen, S.; et al. Biological and immunological relations among human parvovirus B19 genotypes 1 to 3. J. Virol. 2007, 81, 6927–6935. [Google Scholar] [CrossRef]
- Erdman, D.D.; Durigon, E.L.; Wang, Q.-Y.; Anderson, L.J. Genetic diversity of human parvovirus B19: Sequence analysis of the VP1/VP2 gene from multiple isolates. J. Gen. Virol. 1996, 77, 2767–2774. [Google Scholar] [CrossRef]
- Gallinella, G.; Venturoli, S.; Gentilomi, G.; Musiani, M.; Zerbini, M. Extent of sequence variability in a genomic region coding for capsid proteins of B19 parvovirus. Arch. Virol. 1995, 140, 1119–1125. [Google Scholar] [CrossRef]
- Nguyen, Q.T.; Wong, S.; Heegaard, E.D.; Brown, K.E. Identification and characterization of a second novel human erythrovirus variant, A6. Virology 2002, 301, 374–380. [Google Scholar] [CrossRef]
- Shade, R.O.; Blundell, M.C.; Cotmore, S.F.; Tattersall, P.; Astell, C.R. Nucleotide sequence and genome organization of human parvovirus B19 isolated from the serum of a child during aplastic crisis. J. Virol. 1986, 58, 921–936. [Google Scholar] [CrossRef]
- Dorsch, S.; Liebisch, G.; Kaufmann, B.; von Landenberg, P.; Hoffmann, J.H.; Drobnik, W.; Modrow, S. The VP1 unique region of parvovirus B19 and its constituent phospholipase A2-like activity. J. Virol. 2002, 76, 2014–2018. [Google Scholar] [CrossRef] [PubMed]
- Zadori, Z.; Szelei, J.; Lacoste, M.C.; Li, Y.; Gariepy, S.; Raymond, P.; Allaire, M.; Nabi, I.R.; Tijssen, P. A viral phospholipase A2 is required for parvovirus infectivity. Dev. Cell 2001, 1, 291–302. [Google Scholar] [CrossRef]
- Craig Corcoran, D.H. Genetic Variants of Human Parvovirus B19 in South Africa: Cocirculation of Three Genotypes and Identification of a Novel Subtype of Genotype 1. J. Clin. Biol. 2010, 48, 137–142. [Google Scholar] [CrossRef] [PubMed]
- Hubschen, J.M.; Mihneva, Z.; Mentis, A.F.; Schneider, F.; Aboudy, Y.; Grossman, Z.; Rudich, H.; Kasymbekova, K.; Sarv, I.; Nedeljkovic, J.; et al. Phylogenetic analysis of human parvovirus b19 sequences from eleven different countries confirms the predominance of genotype 1 and suggests the spread of genotype 3b. J. Clin. Microbiol. 2009, 47, 3735–3738. [Google Scholar] [CrossRef]
- Parsyan, A.; Szmaragd, C.; Allain, J.P.; Candotti, D. Identification and genetic diversity of two human parvovirus B19 genotype 3 subtypes. J. Gen. Virol. 2007, 88, 428–431. [Google Scholar] [CrossRef] [PubMed]
- Candotti, D.; Etiz, N.; Parsyan, A.; Allain, J.P. Identification and characterization of persistent human erythrovirus infection in blood donor samples. J. Virol. 2004, 78, 12169–12178. [Google Scholar] [CrossRef]
- Parsyan, A.; Candotti, D. Human erythrovirus B19 and blood transfusion–an update. Transfus. Med. 2007, 17, 263–278. [Google Scholar] [CrossRef]
- Willkommen, H.; Schmidt, I.; Lower, J. Safety issues for plasma derivatives and benefit from NAT testing. Biologicals 1999, 27, 325–331. [Google Scholar] [CrossRef]
- Soucie, J.M.; De Staercke, C.; Monahan, P.E.; Recht, M.; Chitlur, M.B.; Gruppo, R.; Hooper, W.C.; Kessler, C.; Kulkarni, R.; Manco-Johnson, M.J.; et al. Evidence for the transmission of parvovirus B19 in patients with bleeding disorders treated with plasma-derived factor concentrates in the era of nucleic acid test screening. Transfusion 2013, 53, 1217–1225. [Google Scholar] [CrossRef]
- Liumbruno, G.M.; Franchini, M. Solvent/detergent plasma: Pharmaceutical characteristics and clinical experience. J. Thromb. Thrombolysis 2015, 39, 118–128. [Google Scholar] [CrossRef]
- Blumel, J.; Schmidt, I.; Effenberger, W.; Seitz, H.; Willkommen, H.; Brackmann, H.H.; Lower, J.; Eis-Hubinger, A.M. Parvovirus B19 transmission by heat-treated clotting factor concentrates. Transfusion 2002, 42, 1473–1481. [Google Scholar] [CrossRef] [PubMed]
- Solheim, B.G.; Rollag, H.; Svennevig, J.L.; Arafa, O.; Fosse, E.; Bergerud, U. Viral safety of solvent/detergent-treated plasma. Transfusion 2000, 40, 84–90. [Google Scholar] [CrossRef]
- Lefrere, J.J.; Mariotti, M. Use of digoxigenin-labelled probes for the detection of B19 parvovirus DNA in batches of blood products. Cell. Mol. Biol. 1995, 41, 985–988. [Google Scholar]
- Lefrere, J.J.; Mariotti, M.; De la Croix, I.; Lerable, J.; Thauvin, M.; Burnouf, T.; Follea, G. Albumin batches and B19 parvovirus DNA. Transfusion 1995, 35, 389–391. [Google Scholar] [CrossRef]
- Lefrere, J.J.; Mariotti, M.; Thauvin, M. B19 parvovirus DNA in solvent/detergent-treated anti-haemophilia concentrates. Lancet 1994, 343, 211–212. [Google Scholar] [CrossRef]
- McOmish, F.; Yap, P.L.; Jordan, A.; Hart, H.; Cohen, B.J.; Simmonds, P. Detection of parvovirus B19 in donated blood: A model system for screening by polymerase chain reaction. J. Clin. Microbiol. 1993, 31, 323–328. [Google Scholar] [CrossRef] [PubMed]
- Rollag, H.; Solheim, B.G.; Svennevig, J.L. Viral safety of blood derivatives by immune neutralization. Vox Sang. 1998, 74 (Suppl. 1), 213–217. [Google Scholar] [CrossRef] [PubMed]
- Saldanha, J.; Minor, P. Detection of human parvovirus B19 DNA in plasma pools and blood products derived from these pools: Implications for efficiency and consistency of removal of B19 DNA during manufacture. Br. J. Haematol. 1996, 93, 714–719. [Google Scholar] [CrossRef]
- Yaegashi, N.; Niinuma, T.; Chisaka, H.; Watanabe, T.; Uehara, S.; Okamura, K.; Moffatt, S.; Sugamura, K.; Yajima, A. The incidence of, and factors leading to, parvovirus B19-related hydrops fetalis following maternal infection; report of 10 cases and meta-analysis. J. Infect. 1998, 37, 28–35. [Google Scholar] [CrossRef]
- Juhl, D.; Hennig, H. Parvovirus B19: What is the Relevance in Transfusion Medicine? Front. Med. 2018, 5, 4. [Google Scholar] [CrossRef] [PubMed]
- Wakamatsu, C.; Takakura, F.; Kojima, E.; Kiriyama, Y.; Goto, N.; Matsumoto, K.; Oyama, M.; Sato, H.; Okochi, K.; Maeda, Y. Screening of blood donors for human parvovirus B19 and characterization of the results. Vox Sang. 1999, 76, 14–21. [Google Scholar] [CrossRef] [PubMed]
- Schmidt, M.; Themann, A.; Drexler, C.; Bayer, M.; Lanzer, G.; Menichetti, E.; Lechner, S.; Wessin, D.; Prokoph, B.; Allain, J.P.; et al. Blood donor screening for parvovirus B19 in Germany and Austria. Transfusion 2007, 47, 1775–1782. [Google Scholar] [CrossRef] [PubMed]
- Oh, D.J.; Lee, Y.L.; Kang, J.W.; Kwon, S.Y.; Cho, N.S. Investigation of the Prevalence of Human Parvovirus B19 DNA in Korean Plasmapheresis Donors. Korean J. Lab. Med. 2010, 30, 58–64. [Google Scholar] [CrossRef]
- Thomas, I.; Di Giambattista, M.; Gerard, C.; Mathys, E.; Hougardy, V.; Latour, B.; Branckaert, T.; Laub, R. Prevalence of human erythrovirus B19 DNA in healthy Belgian blood donors and correlation with specific antibodies against structural and non-structural viral proteins. Vox Sang. 2003, 84, 300–307. [Google Scholar] [CrossRef]
- Yoto, Y.; Kudoh, T.; Haseyama, K.; Suzuki, N.; Oda, T.; Katoh, T.; Takahashi, T.; Sekiguchi, S.; Chiba, S. Incidence of human parvovirus B19 DNA detection in blood donors. Br. J. Haematol. 1995, 91, 1017–1018. [Google Scholar] [CrossRef]
- Kleinman, S.H.; Glynn, S.A.; Lee, T.-H.; Tobler, L.H.; Schlumpf, K.S.; Todd, D.S.; Qiao, H.; Yu, M.-Y.W.; Busch, M.P. A linked donor-recipient study to evaluate parvovirus B19 transmission by blood component transfusion. Blood 2009, 114, 3677–3683. [Google Scholar] [CrossRef]
- Average Age of Expatriates Living in the Gulf Cooperation Council as of March 2018, by Country; Statista, Germany. 2018. Available online: https://www.statista.com/statistics/806066/gcc-average-age-of-expats-by-country/ (accessed on 15 January 2021).
- Nasrallah, G.K.; Al Absi, E.S.; Ghandour, R.; Ali, N.H.; Taleb, S.; Hedaya, L.; Ali, F.; Huwaidy, M.; Husseini, A. Seroprevalence of hepatitis E virus among blood donors in Qatar (2013–2016). Transfusion 2017, 57, 1801–1807. [Google Scholar] [CrossRef]
- Al-Qahtani, A.A.; Alabsi, E.S.; AbuOdeh, R.; Thalib, L.; El Zowalaty, M.E.; Nasrallah, G.K. Prevalence of anelloviruses (TTV, TTMDV, and TTMV) in healthy blood donors and in patients infected with HBV or HCV in Qatar. Virol. J. 2016, 13, 208. [Google Scholar] [CrossRef]
- Amimo, J.O.; El Zowalaty, M.E.; Githae, D.; Wamalwa, M.; Djikeng, A.; Nasrallah, G.K. Metagenomic analysis demonstrates the diversity of the fecal virome in asymptomatic pigs in East Africa. Arch. Virol. 2016, 161, 887–897. [Google Scholar] [CrossRef]
- AbuOdeh, R.O.; Al-Absi, E.; Ali, N.H.; Khalili, M.; Al-Mawlawi, N.; Hadwan, T.A.; Althani, A.A.; Nasrallah, G.K. Detection and phylogenetic analysis of human pegivirus (GBV-C) among blood donors and patients infected with hepatitis B virus (HBV) in Qatar. J. Med. Virol. 2015, 87, 2074–2081. [Google Scholar] [CrossRef]
- AbuOdeh, R.; Al-Mawlawi, N.; Al-Qahtani, A.A.; Bohol, M.F.F.; Al-Ahdal, M.N.; Hasan, H.A.; AbuOdeh, L.; Nasrallah, G.K. Detection and genotyping of torque teno virus (TTV) in healthy blood donors and patients infected with HBV or HCV in Qatar. J. Med. Virol. 2015, 87, 1184–1191. [Google Scholar] [CrossRef]
- Siritantikorn, S.; Kaewrawang, S.; Siritanaratkul, N.; Theamboonlers, A.; Poovorawan, Y.; Kantakamalakul, W.; Wasi, C. The prevalence and persistence of human parvovirus B19 infection in thalassemic patients. Asian Pac. J. Allergy Immunol. 2007, 25, 169–174. [Google Scholar]
- Toan, N.L.; Duechting, A.; Kremsner, P.G.; Song le, H.; Ebinger, M.; Aberle, S.; Binh, V.Q.; Duy, D.N.; Torresi, J.; Kandolf, R.; et al. Phylogenetic analysis of human parvovirus B19, indicating two subgroups of genotype 1 in Vietnamese patients. J. Gen. Virol. 2006, 87, 2941–2949. [Google Scholar] [CrossRef]
- Manaresi, E.; Gallinella, G.; Zuffi, E.; Bonvicini, F.; Zerbini, M.; Musiani, M. Diagnosis and quantitative evaluation of parvovirus B19 infections by real-time PCR in the clinical laboratory. J. Med. Virol. 2002, 67, 275–281. [Google Scholar] [CrossRef] [PubMed]
- Oakley, G. Final Diagnosis–Acute Parvovirus Infection. Available online: https://path.upmc.edu/cases/case522/dx.html (accessed on 16 February 2020).
- Jia, J.; Ma, Y.; Zhao, X.; Huangfu, C.; Zhong, Y.; Fang, C.; Fan, R.; Lv, M.; Zhang, J. Existence of various human parvovirus B19 genotypes in Chinese plasma pools: Genotype 1, genotype 3, putative intergenotypic recombinant variants and new genotypes. Virol. J. 2016, 13, 155. [Google Scholar] [CrossRef] [PubMed]
- Blundell, M.C.; Beard, C.; Astell, C.R. In vitro identification of a B19 parvovirus promoter. Virology 1987, 157, 534–538. [Google Scholar] [CrossRef]
- Grabarczyk, P.; Kalinska, A.; Kara, M.; Wieczorek, R.; Ejduk, A.; Sulkowska, E.; Golebiowska-Staroszczyk, S.; Matysiak, M.; Baylis, S.A.; Brojer, E. Identification and characterization of acute infection with parvovirus B19 genotype 2 in immunocompromised patients in Poland. J. Med. Virol. 2011, 83, 142–149. [Google Scholar] [CrossRef] [PubMed]
- Rinckel, L.A.; Buno, B.R.; Gierman, T.M.; Lee, D.C. Discovery and analysis of a novel parvovirus B19 Genotype 3 isolate in the United States. Transfusion 2009, 49, 1488–1492. [Google Scholar] [CrossRef]
- Jordan, J.; Tiangco, B.; Kiss, J.; Koch, W. Human parvovirus B19: Prevalence of viral DNA in volunteer blood donors and clinical outcomes of transfusion recipients. Vox Sang. 1998, 75, 97–102. [Google Scholar] [CrossRef]
- Weimer, T.; Streichert, S.; Watson, C.; Groner, A. High-titer screening PCR: A successful strategy for reducing the parvovirus B19 load in plasma pools for fractionation. Transfusion 2001, 41, 1500–1504. [Google Scholar] [CrossRef]
- Satake, M.; Hoshi, Y.; Taira, R.; Momose, S.Y.; Hino, S.; Tadokoro, K. Symptomatic parvovirus B19 infection caused by blood component transfusion. Transfusion 2011, 51, 1887–1895. [Google Scholar] [CrossRef]
- Patou, G.; Pillay, D.; Myint, S.; Pattison, J. Characterization of a nested polymerase chain reaction assay for detection of parvovirus B19. J. Clin. Microbiol. 1993, 31, 540–546. [Google Scholar] [CrossRef]
- Grabarczyk, P.; Korzeniowska, J.; Liszewski, G.; Kalinska, A.; Sulkowska, E.; Krug-Janiak, M.; Kopacz, A.; Letowska, M.; Brojer, E. [Parvovirus B19 DNA testing in Polish blood donors, 2004–2010]. Prz. Epidemiol. 2012, 66, 7–12. [Google Scholar]
- Sakata, H.; Matsubayashi, K.; Ihara, H.; Sato, S.; Kato, T.; Wakisaka, A.; Tadokoro, K.; Yu, M.Y.; Baylis, S.A.; Ikeda, H.; et al. Impact of chemiluminescent enzyme immunoassay screening for human parvovirus B19 antigen in Japanese blood donors. Transfusion 2013, 53, 2556–2566. [Google Scholar] [CrossRef]
- Letaïef, M.V.G.; Boukef, K.; Yacoub, S.; Muylle, L.; Mertens, G. Higher prevalence of parvovirus B19 in Belgian as compared to Tunisian blood donors: Differential implications for prevention of transfusional transmission. Transfus. Sci. 1997, 18, 523–530. [Google Scholar] [CrossRef]
- Hassan Omer, A.; Jamal Al deen Adam, A.; Ali Mohamed, F.; Hashim Hamed, F.; Hussain Edr, H.; Ahmed Al sedig, M.; Hashim Ahmed, R.; Ismail Mohammed, Y.; Mohammed Hamid, O. Seroprevalance of Human Parvovirus B19 among Blood Donor Volunteers from Sudanese Blood Bank in Khartoum State 2017. J. Immuno. Biol. 2017, 3, 1–4. [Google Scholar] [CrossRef]
- Kishore, J.; Srivastava, M.; Choudhary, N. Standardization of B19 IgG ELISA to study the seroepidemiology of parvovirus B19 in North Indian voluntary blood donors. Asian J. Transfus. Sci. 2010, 4, 86–90. [Google Scholar] [CrossRef] [PubMed]
- Kooistra, K.; Mesman, H.J.; de Waal, M.; Koppelman, M.H.G.M.; Zaaijer, H.L. Epidemiology of high-level parvovirus B19 viraemia among Dutch blood donors, 2003–2009. Vox Sang. 2010, 100, 261–266. [Google Scholar] [CrossRef]
- Zaaijer, H.L.; Koppelman, M.H.; Farrington, C.P. Parvovirus B19 viraemia in Dutch blood donors. Epidemiol. Infect. 2004, 132, 1161–1166. [Google Scholar] [CrossRef] [PubMed]
- Slavov, S.N.; Haddad, S.K.; Silva-Pinto, A.C.; Amarilla, A.A.; Alfonso, H.L.; Aquino, V.H.; Covas, D.T. Molecular and phylogenetic analyses of human Parvovirus B19 isolated from Brazilian patients with sickle cell disease and beta-thalassemia major and healthy blood donors. J. Med. Virol. 2012, 84, 1652–1665. [Google Scholar] [CrossRef]
- Munoz, S.; Alonso, M.A.; Fernandez, M.J.; Munoz, J.L.; Garcia-Rodriguez, J.A. Seroprevalence versus Parvovirus B19 in blood donors. Enferm. Infecc. Microbiol. Clin. 1998, 16, 161–162. [Google Scholar] [PubMed]
- Johargy, A.K. Seroprevalence of erythrovirus B19 IgG among saudi blood donors in makkah, Saudi Arabia. J. Fam. Community Med. 2009, 16, 111–114. [Google Scholar]
- Manaresi, E.; Gallinella, G.; Morselli Labate, A.M.; Zucchelli, P.; Zaccarelli, D.; Ambretti, S.; Delbarba, S.; Zerbini, M.; Musiani, M. Seroprevalence of IgG against conformational and linear capsid antigens of parvovirus B19 in Italian blood donors. Epidemiol. Infect. 2004, 132, 857–862. [Google Scholar] [CrossRef]
- Kumar, S.; Gupta, R.M.; Sen, S.; Sarkar, R.S.; Philip, J.; Kotwal, A.; Sumathi, S.H. Seroprevalence of human parvovirus B19 in healthy blood donors. Med. J. Armed Forces India 2013, 69, 268–272. [Google Scholar] [CrossRef] [PubMed]
- Ou, S.H.; Xie, J.Z.; Zhang, Y.L.; Ni, H.Y.; Song, X.Y. Prevalence of Parvovirus B19 Infection in Chinese Xiamen Area Blood Donors. Zhongguo Shi Yan Xue Ye Xue Za Zhi 2016, 24, 1572–1576. [Google Scholar] [CrossRef]
- Zadsar, M.; Aghakhani, A.; Banifazl, M.; Kazemimanesh, M.; Tabatabaei Yazdi, S.M.; Mamishi, S.; Bavand, A.; Sadat Larijani, M.; Ramezani, A. Seroprevalence, molecular epidemiology and quantitation of parvovirus B19 DNA levels in Iranian blood donors. J. Med. Virol. 2018, 90, 1318–1322. [Google Scholar] [CrossRef] [PubMed]
- Anderson, L.J. Role of parvovirus B19 in human disease. Pediatr. Infect. Dis. J. 1987, 6, 711–718. [Google Scholar] [CrossRef] [PubMed]
- Lin, K.H.; You, S.L.; Chen, C.J.; Wang, C.F.; Yang, C.S.; Yamazaki, S. Seroepidemiology of human parvovirus B19 in Taiwan. J. Med. Virol. 1999, 57, 169–173. [Google Scholar] [CrossRef]
- De Freitas, R.B.; Wong, D.; Boswell, F.; de Miranda, M.F.; Linhares, A.C.; Shirley, J.; Desselberger, U. Prevalence of human parvovirus (B19) and rubella virus infections in urban and remote rural areas in northern Brazil. J. Med. Virol. 1990, 32, 203–208. [Google Scholar] [CrossRef]
- Koch, W.C.; Adler, S.P. Human parvovirus B19 infections in women of childbearing age and within families. Pediatr. Infect. Dis. J. 1989, 8, 83–87. [Google Scholar]
- Kelly, H.A.; Siebert, D.; Hammond, R.; Leydon, J.; Kiely, P.; Maskill, W. The age-specific prevalence of human parvovirus immunity in Victoria, Australia compared with other parts of the world. Epidemiol. Infect. 2000, 124, 449–457. [Google Scholar] [CrossRef] [PubMed]
- Nascimento, J.P.; Buckley, M.M.; Brown, K.E.; Cohen, B.J. The prevalence of antibody to human parvovirus B19 in Rio de Janeiro, Brazil. Rev. Inst. Med. Trop. Sao Paulo 1990, 32, 41–45. [Google Scholar] [CrossRef] [PubMed]
- Al-Danani, D.A.-D.; Saudi, M.H.; Al-Shamahy, H. Seroprevalence of parvovirus B19 IgG among blood donors in Aden, Yemen and Alexandria, Egypt. J. Chin. Clin. Med. 2008, 3, 173–176. [Google Scholar]
- Elisha, M.; Yakubu, R.K.; Rufai, O.A. Risk of Transfusion-Transmitted Human Parvovirus B19 Infection in Anyigba and Lokoja, Kogi State—Nigeria. Iosr J. Pharm. 2013, 3, 66–70. [Google Scholar]
- Goral, S.; Yenicesu, I.; Bozdayi, G.; Duyan Camurdan, A.; Altay Kocak, A. Parvovirus B19 seroprevalence in Turkish blood donors. Turk. J. Med. Sci. 2018, 48, 956–960. [Google Scholar] [CrossRef]
- Wasfy, S.; Nishikawa, J.; Petric, M. Seroprevalence of immunoglobulin G antibody to parvovirus B19 in Ontario. Can. J. Infect. Dis. J. Can. Des Mal. Infect. 1996, 7, 313–316. [Google Scholar] [CrossRef] [PubMed]
- Van Rijckevorsel, G.G.; Sonder, G.J.; Schim van der Loeff, M.F.; van den Hoek, J.A. Population-based study on the seroprevalence of parvovirus B19 in Amsterdam. J. Med. Virol. 2009, 81, 1305–1309. [Google Scholar] [CrossRef] [PubMed]
- Mendelson, E.; Aboudy, Y.; Smetana, Z.; Tepperberg, M.; Grossman, Z. Laboratory assessment and diagnosis of congenital viral infections: Rubella, cytomegalovirus (CMV), varicella-zoster virus (VZV), herpes simplex virus (HSV), parvovirus B19 and human immunodeficiency virus (HIV). Reprod. Toxicol. 2006, 21, 350–382. [Google Scholar] [CrossRef] [PubMed]
- Kleinman, S.H.; Glynn, S.A.; Lee, T.H.; Tobler, L.; Montalvo, L.; Todd, D.; Kiss, J.E.; Shyamala, V.; Busch, M.P. Prevalence and quantitation of parvovirus B19 DNA levels in blood donors with a sensitive polymerase chain reaction screening assay. Transfusion 2007, 47, 1756–1764. [Google Scholar] [CrossRef]
- Liefeldt, L.; Plentz, A.; Klempa, B.; Kershaw, O.; Endres, A.S.; Raab, U.; Neumayer, H.H.; Meisel, H.; Modrow, S. Recurrent high level parvovirus B19/genotype 2 viremia in a renal transplant recipient analyzed by real-time PCR for simultaneous detection of genotypes 1 to 3. J. Med. Virol. 2005, 75, 161–169. [Google Scholar] [CrossRef]
- Enders, M.; Schalasta, G.; Baisch, C.; Weidner, A.; Pukkila, L.; Kaikkonen, L.; Lankinen, H.; Hedman, L.; Soderlund-Venermo, M.; Hedman, K. Human parvovirus B19 infection during pregnancy--value of modern molecular and serological diagnostics. J. Clin. Virol. 2006, 35, 400–406. [Google Scholar] [CrossRef]
- Maple, P.A.; Hedman, L.; Dhanilall, P.; Kantola, K.; Nurmi, V.; Soderlund-Venermo, M.; Brown, K.E.; Hedman, K. Identification of past and recent parvovirus B19 infection in immunocompetent individuals by quantitative PCR and enzyme immunoassays: A dual-laboratory study. J. Clin. Microbiol. 2014, 52, 947–956. [Google Scholar] [CrossRef]
- De Jong, E.P.; de Haan, T.R.; Kroes, A.C.; Beersma, M.F.; Oepkes, D.; Walther, F.J. Parvovirus B19 infection in pregnancy. J. Clin. Virol. 2006, 36, 1–7. [Google Scholar] [CrossRef]
- Bredl, S.; Plentz, A.; Wenzel, J.J.; Pfister, H.; Most, J.; Modrow, S. False-negative serology in patients with acute parvovirus B19 infection. J. Clin. Virol. 2011, 51, 115–120. [Google Scholar] [CrossRef]
- Henriques, I.; Monteiro, F.; Meireles, E.; Cruz, A.; Tavares, G.; Ferreira, M.; Araújo, F. Prevalence of Parvovirus B19 and Hepatitis A virus in Portuguese blood donors. Transfus. Apher. Sci. 2005, 33, 305–309. [Google Scholar] [CrossRef]
- Ivanova, S.K.; Mihneva, Z.G.; Toshev, A.K.; Kovaleva, V.P.; Andonova, L.G.; Muller, C.P.; Hübschen, J.M. Insights into epidemiology of human parvovirus B19 and detection of an unusual genotype 2 variant, Bulgaria, 2004 to 2013. Eurosurveillance 2016, 21, 30116. [Google Scholar] [CrossRef]
- Hokynar, K.; Brunstein, J.; Soderlund-Venermo, M.; Kiviluoto, O.; Partio, E.K.; Konttinen, Y.; Hedman, K. Integrity and full coding sequence of B19 virus DNA persisting in human synovial tissue. J. Gen. Virol. 2000, 81, 1017–1025. [Google Scholar] [CrossRef] [PubMed]
- Lukashov, V.V.; Goudsmit, J. Evolutionary relationships among parvoviruses: Virus-host coevolution among autonomous primate parvoviruses and links between adeno-associated and avian parvoviruses. J. Virol. 2001, 75, 2729–2740. [Google Scholar] [CrossRef] [PubMed]
- Heegaard, E.D.; Panum Jensen, I.; Christensen, J. Novel PCR assay for differential detection and screening of erythrovirus B19 and erythrovirus V9. J. Med. Virol. 2001, 65, 362–367. [Google Scholar] [CrossRef] [PubMed]
- Slavov, S.N.; Otaguiri, K.K.; Covas, D.T.; Kashima, S. Prevalence and Viral Load of Human Parvovirus B19 (B19V) among Blood Donors in South-East Brazil. Indian J. Hematol. Blood Transfus. 2016, 32, 323–325. [Google Scholar] [CrossRef]
- Hemauer, A.; von Poblotzki, A.; Gigler, A.; Cassinotti, P.; Siegl, G.; Wolf, H.; Modrow, S. Sequence variability among different parvovirus B19 isolates. J. Gen. Virol. 1996, 77 Pt 8, 1781–1785. [Google Scholar] [CrossRef]
- Gallinella, G.; Venturoli, S.; Manaresi, E.; Musiani, M.; Zerbini, M. B19 virus genome diversity: Epidemiological and clinical correlations. J. Clin. Virol. 2003, 28, 1–13. [Google Scholar] [CrossRef]
- Servant-Delmas, A.; Lefrère, J.J.; Morinet, F.; Pillet, S. Advances in Human B19 Erythrovirus Biology. J. Virol. 2010, 84, 9658–9665. [Google Scholar] [CrossRef]
- Suzuki, M.; Yoto, Y.; Ishikawa, A.; Tsutsumi, H. Analysis of Nucleotide Sequences of Human Parvovirus B19 Genome Reveals Two Different Modes of Evolution, a Gradual Alteration and a Sudden Replacement: A Retrospective Study in Sapporo, Japan, from 1980 to 2008. J. Virol. 2009, 83, 10975–10980. [Google Scholar] [CrossRef]
- Schneider, B.; Hone, A.; Tolba, R.H.; Fischer, H.P.; Blumel, J.; Eis-Hubinger, A.M. Simultaneous persistence of multiple genome variants of human parvovirus B19. J. Gen. Virol. 2008, 89, 164–176. [Google Scholar] [CrossRef] [PubMed]
- Da Costa, A.C.; Bendit, I.; de Oliveira, A.C.S.; Kallas, E.G.; Sabino, E.C.; Sanabani, S.S. Investigation of human parvovirus B19 occurrence and genetic variability in different leukaemia entities. Clin. Microbiol. Infect. 2013, 19, E31–E43. [Google Scholar] [CrossRef] [PubMed]
- Molenaar-de Backer, M.W.A.; Lukashov, V.V.; van Binnendijk, R.S.; Boot, H.J.; Zaaijer, H.L. Global co-existence of two evolutionary lineages of parvovirus B19 1a, different in genome-wide synonymous positions. PLoS ONE 2012, 7, e43206. [Google Scholar] [CrossRef]
- Hokynar, K.; Norja, P.; Laitinen, H.; Palomaki, P.; Garbarg-Chenon, A.; Ranki, A.; Hedman, K.; Soderlund-Venermo, M. Detection and differentiation of human parvovirus variants by commercial quantitative real-time PCR tests. J. Clin. Microbiol. 2004, 42, 2013–2019. [Google Scholar] [CrossRef]
- Sanabani, S.; Neto, W.K.; Pereira, J.; Sabino, E.C. Sequence Variability of Human Erythroviruses Present in Bone Marrow of Brazilian Patients with Various Parvovirus B19-Related Hematological Symptoms. J. Clin. Microbiol. 2006, 44, 604–606. [Google Scholar] [CrossRef]
- Doerig, C.; Hirt, B.; Antonietti, J.P.; Beard, P. Nonstructural protein of parvoviruses B19 and minute virus of mice controls transcription. J. Virol. 1990, 64, 387–396. [Google Scholar] [CrossRef]
- Kivovich, V.; Gilbert, L.; Vuento, M.; Naides, S.J. Parvovirus B19 genotype specific amino acid substitution in NS1 reduces the protein’s cytotoxicity in culture. Int. J. Med. Sci. 2010, 7, 110–119. [Google Scholar] [CrossRef] [PubMed]
- Ozawa, K.; Ayub, J.; Kajigaya, S.; Shimada, T.; Young, N. The gene encoding the nonstructural protein of B19 (human) parvovirus may be lethal in transfected cells. J. Virol. 1988, 62, 2884–2889. [Google Scholar] [CrossRef] [PubMed]
- Kerr, J.R.; Curran, M.D.; Moore, J.E.; Erdman, D.D.; Coyle, P.V.; Nunoue, T.; Middleton, D.; Ferguson, W.P. Genetic diversity in the non-structural gene of parvovirus B19 detected by single-stranded conformational polymorphism assay (SSCP) and partial nucleotide sequencing. J. Virol. Methods 1995, 53, 213–222. [Google Scholar] [CrossRef]
- Blümel, J.; Eis-Hübinger, A.M.; Stühler, A.; Bönsch, C.; Gessner, M.; Löwer, J. Characterization of Parvovirus B19 genotype 2 in KU812Ep6 cells. J. Virol. 2005, 79, 14197–14206. [Google Scholar] [CrossRef]
- Parsyan, A.; Kerr, S.; Owusu-Ofori, S.; Elliott, G.; Allain, J.P. Reactivity of genotype-specific recombinant proteins of human erythrovirus B19 with plasmas from areas where genotype 1 or 3 is endemic. J. Clin. Microbiol. 2006, 44, 1367–1375. [Google Scholar] [CrossRef] [PubMed]
- Candotti, D.; Danso, K.; Parsyan, A.; Dompreh, A.; Allain, J.P. Maternal-fetal transmission of human parvovirus B19 genotype 3. J. Infect. Dis. 2006, 194, 608–611. [Google Scholar] [CrossRef] [PubMed]
- Parsyan, A.; Addo-Yobo, E.; Owusu-Ofori, S.; Akpene, H.; Sarkodie, F.; Allain, J.P. Effects of transfusion on human erythrovirus B19-susceptible or -infected pediatric recipients in a genotype 3-endemic area. Transfusion 2006, 46, 1593–1600. [Google Scholar] [CrossRef]
- Stamenković, G.G.; Ćirković, V.S.; Šiljić, M.M.; Blagojević, J.V.; Knežević, A.M.; Joksić, I.D.; Stanojević, M.P. Substitution rate and natural selection in parvovirus B19. Sci. Rep. 2016, 6, 1–9. [Google Scholar] [CrossRef]
- Chirambo-Kalolekesha, M.; Kaile, T.; Mwaba, F.; Daka, V.; Simakando, M.; Kowa, S. Seroprevalence of parvovirus B19 in blood donors: The risks and challenges of blood transfusion in Zambia in the era of HIV/AIDS at the Kitwe Central Hospital, blood bank. Afr. Health Sci. 2018, 18, 496–502. [Google Scholar] [CrossRef]
- Nabae, K.; Satoh, H.; Nishiura, H.; Tanaka-Taya, K.; Okabe, N.; Oishi, K.; Matsumoto, K.; Hasegawa, T. Estimating the Risk of Parvovirus B19 Infection in Blood Donors and Pregnant Women in Japan. PLoS ONE 2014, 9, e92519. [Google Scholar] [CrossRef]
- Baylis, S.A. Standardization of nucleic acid amplification technique (NAT)-based assays for different genotypes of parvovirus B19: A meeting summary. Vox Sang. 2008, 94, 74–80. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Interpretation | B19V Marker | ||
|---|---|---|---|
| DNA | IgM | IgG | |
| Acute infection | + | +/− | − |
| Persistent | + | − | + |
| Reactivated | + | + | + |
| Characteristic | No (%) | |
|---|---|---|
| Sex | ||
| Male | 4862 (96.73) | |
| Female | 149 (2.96) | |
| Unknown | 15 (0.29) | |
| Total | 5026 | |
| Age (years) | ||
| 19–30 | 1124 (22.36) | |
| 30–40 | 2061 (41.00) | |
| 40–50 | 1275 (25.36) | |
| >50 | 543 (10.84) | |
| Unknown | 23 | |
| Total | 5026 | |
| Nationality | ||
| Qatari | 1045 (21) | |
| Non-Qatari | 3981 (79) | |
| Egyptian | 827 (16.4) | |
| Sudanese | 112 (2.2) | |
| Philippine | 115 (2.3) | |
| Indian | 555 (11) | |
| Sri Lankan | 77 (1.5) | |
| Lebanese | 108 (2.1) | |
| Jordanian | 311 (6.2) | |
| Palestinian | 271 (5.4) | |
| Syrian | 644 (12.8) | |
| Pakistan | 197 (3.9) | |
| Iranian | 99 (2) | |
| Yemen | 124 (2.5) | |
| Other * | 541 (11) | |
| Total | 5026 |
| Category | Total No. | B19V DNA qPCR Positive No. (%) | p-Value * |
|---|---|---|---|
| Gender | |||
| Male | 4862 | 68 (1.4) | 0.453 |
| Female | 149 | 1 (0.7) | |
| Unknown | 15 | 1 (6.7) | |
| Nationality | |||
| Qatari | 1045 | 16 (1.5) | |
| Male | 1007 | 15 (1.5) | |
| Female | 37 | 0 (0.0) | |
| Non-Qatari | 3937 | 49 (1.2) | 0.468 |
| Male | 3825 | 48 (1.3) | |
| Female | 111 | 1 (0.9) | |
| Unknown | 44 | 1 (2.3) | |
| Age Group | |||
| 19–30 | 1124 | 25 (2.2) | |
| 31–40 | 2061 | 26 (1.3) | 0.034 |
| 41–50 | 1275 | 11 (0.9) | |
| > 51 | 543 | 7 (1.3) | |
| Unknown | 23 | 1 (4.4) |
| Category by ELISA | No. (%) | Viral Load Copies/mL of Blood |
|---|---|---|
| Acute infection | 4 (5.7) | - |
| Reactivated | 20 (28.6) | 1.2 × 101–5.95 × 106 |
| Persistent | 46 (65.7) | 7.41 × 101–8.27 × 106 |
| Total | 70 |
| Category | Total No. | B19V Serology | |||
|---|---|---|---|---|---|
| IgG-Positive No. (%) | p-Value * | IgM-Positive No. (%) | p-Value * | ||
| Gender | |||||
| Male | 808 | 495 (61.2) | 0.0449 | 17 (2.1) | 0.649 |
| Female | 122 | 66 (54.1) | 3 (2.4) | ||
| Total | 930 | 561 (60.3) | 20 (2.1) | ||
| Nationality | |||||
| Qatari | 298 | 178 (59.7) | 0.6335 | 10 (3.3) | 0.648 |
| Male | 261 | 158 (60.5) | 9 (3.4) | ||
| Female | 37 | 20 (54.1) | 1 (2.7) | ||
| Non-Qatari | 632 | 383 (60.6) | 10 (1.6) | ||
| Male | 547 | 337 (61.6) | 8 (1.4) | ||
| Female | 85 | 46 (54.1) | 2 (2.3) | ||
| Age Group | |||||
| 19–30 | 307 | 182 (59.3) | 0.9194 | 4 (1.3) | 0.2132 |
| 31–40 | 335 | 203 (60.6) | 7 (2.1) | ||
| 41–50 | 209 | 127 (60.7) | 9 (4.3) | ||
| > 51 | 76 | 48 (63.1) | 0 (0) | ||
| Unknown | 3 | 1 (33.3) | 0 (0) | ||
| nt Position | nt Change | Amino Acid Codon | Amino Acid Change | No of Samples with Mutation |
|---|---|---|---|---|
| 1926 | C/G | 1926 | R > T | 1 |
| 1929 | C/T | 1929 | F > S | 2 |
| 1930 | A/C/T | 1932 | K > Q | 3 |
| 1939 | A/C | 1941 | Q > p, Q > S | 2 |
| 1967 | G/C | 1968 | A > P | 1 |
| 1997 | G/C | 1998 | V > L | 1 |
| 2036 | G/A | 2037 | G > N, G > R | 1 |
| 2235 | T/C | 2235 | V > A | 2 |
| 2244 | A/G | 2244 | N > H, N > C | 3 |
| 2268 | G/C | 2268 | G > A | 5 |
| 2309 | A/G | 2310 | I > V | 1 |
| 2352 | A/G | 2352 | D > G | 5 |
| 2453 | A/G | 2454 | K > E | 8 |
| 2531 | G/C | 2532 | V > L | 5 |
| 2548 | G/A | 2550 | E > K | 1 |
| 2555 | A/G | 2556 | I > K, I > V | 1 |
| 2603 | G/A | 2604 | E > P, E > K | 1 |
| 2616 | C/T | 2616 | S > L | 1 |
| 2620 | G/A | 2622 | F < L | 1 |
| 2629 | G /A | 2631 | V < I | 1 |
| 2646 | A/T | 2646 | N > I | 1 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Abdelrahman, D.; Al-Sadeq, D.W.; Smatti, M.K.; Taleb, S.A.; AbuOdeh, R.O.; Al-Absi, E.S.; Al-Thani, A.A.; Coyle, P.V.; Al-Dewik, N.; Qahtani, A.A.A.; et al. Prevalence and Phylogenetic Analysis of Parvovirus (B19V) among Blood Donors with Different Nationalities Residing in Qatar. Viruses 2021, 13, 540. https://doi.org/10.3390/v13040540
Abdelrahman D, Al-Sadeq DW, Smatti MK, Taleb SA, AbuOdeh RO, Al-Absi ES, Al-Thani AA, Coyle PV, Al-Dewik N, Qahtani AAA, et al. Prevalence and Phylogenetic Analysis of Parvovirus (B19V) among Blood Donors with Different Nationalities Residing in Qatar. Viruses. 2021; 13(4):540. https://doi.org/10.3390/v13040540
Chicago/Turabian StyleAbdelrahman, Doua, Duaa W. Al-Sadeq, Maria K. Smatti, Sara A. Taleb, Raed O AbuOdeh, Enas S. Al-Absi, Asmaa A. Al-Thani, Peter. V. Coyle, Nader Al-Dewik, Ahmed A. Al Qahtani, and et al. 2021. "Prevalence and Phylogenetic Analysis of Parvovirus (B19V) among Blood Donors with Different Nationalities Residing in Qatar" Viruses 13, no. 4: 540. https://doi.org/10.3390/v13040540