
In Vitro and In Vivo Models for Studying SARS-CoV-2, the Etiological Agent Responsible for COVID-19 Pandemic

 , , , , and
, , , , and
Abstract
:1. Introduction
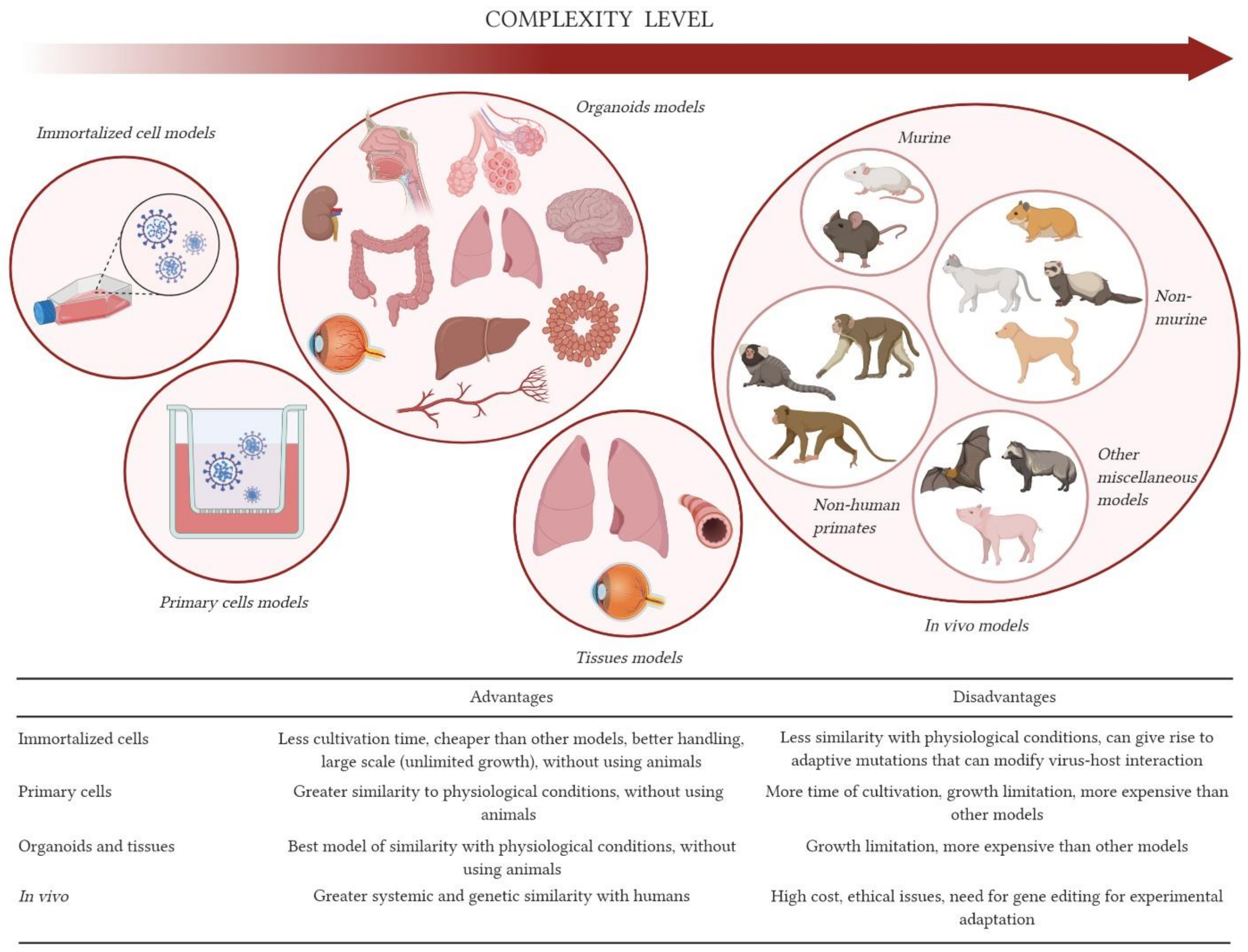
2. In Vitro Models
2.1. 2D-Cell Models: Immortalized Cells
2.2. 3D-Cell Models: Explants and Organoids
3. Animal Models
3.1. Mice
3.2. Golden Syrian Hamsters
3.3. Ferrets
3.4. Dogs
3.5. Cats
3.6. Non-Human Primates
4. Other Miscellaneous Models
5. Concluding Remarks
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Gao, Z.C. Efficient management of novel coronavirus pneumonia by efficient prevention and control in scientific manner. Zhonghua Jie He He Hu Xi Za Zhi 2020, 43, 163–166. [Google Scholar] [CrossRef] [PubMed]
- Wu, F.; Zhao, S.; Yu, B.; Chen, Y.M.; Wang, W.; Song, Z.G.; Hu, Y.; Tao, Z.W.; Tian, J.H.; Pei, Y.Y.; et al. A new coronavirus associated with human respiratory disease in China. Nature 2020, 579, 265–269. [Google Scholar] [CrossRef] [Green Version]
- Gorbalenya, A.E.; Baker, S.C.; Baric, R.S.; de Groot, R.J.; Drosten, C.; Gulyaeva, A.A.; Haagmans, B.L.; Lauber, C.; Leontovich, A.M.; Neuman, B.W.; et al. The species Severe acute respiratory syndrome-related coronavirus: Classifying 2019-nCoV and naming it SARS-CoV-2. Nat. Microbiol. 2020, 5, 536–544. [Google Scholar] [CrossRef] [Green Version]
- Bin, D.; Haibo, Q.; Xi, Z.; Yishan, W.; Hanyujie, K.; Xuyan, L.; Feng, W.; Bing, S.; Tong, Z.H. Pharmacotherapeutics for the new coronavirus pneumonia. Chin. J. Tuberc. Respir. Dis. 2020, 43, 185–188. [Google Scholar] [CrossRef]
- Zheng, J. SARS-coV-2: An emerging coronavirus that causes a global threat. Int. J. Biol. Sci. 2020, 16, 1678–1685. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hong, N.; Yu, W.; Xia, J.; Shen, Y.; Yap, M.; Han, W. Evaluation of ocular symptoms and tropism of SARS-CoV-2 in patients confirmed with COVID-19. Acta Ophthalmol. 2020, 1–7. [Google Scholar] [CrossRef]
- Jin, X.; Lian, J.S.; Hu, J.H.; Gao, J.; Zheng, L.; Zhang, Y.M.; Hao, S.R.; Jia, H.Y.; Cai, H.; Zhang, X.L.; et al. Epidemiological, clinical and virological characteristics of 74 cases of coronavirus-infected disease 2019 (COVID-19) with gastrointestinal symptoms. Gut 2020, 1002–1009. [Google Scholar] [CrossRef] [Green Version]
- Prasad, A.; Prasad, M. Single Virus Targeting Multiple Organs: What We Know and Where We Are Heading? Front. Med. 2020, 7, 5–10. [Google Scholar] [CrossRef]
- Xia, S.; Wu, M.; Chen, S.; Zhang, T.; Ye, L.; Liu, J.; Li, H. Long Term Culture of Human Kidney Proximal Tubule Epithelial Cells Maintains Lineage Functions and Serves as an Ex vivo Model for Coronavirus Associated Kidney Injury. Virol. Sin. 2020, 35, 1–10. [Google Scholar] [CrossRef]
- Hui, K.P.Y.; Cheung, M.C.; Perera, R.A.P.M.; Ng, K.C.; Bui, C.H.T.; Ho, J.C.W.; Ng, M.M.T.; Kuok, D.I.T.; Shih, K.C.; Tsao, S.W.; et al. Tropism, replication competence, and innate immune responses of the coronavirus SARS-CoV-2 in human respiratory tract and conjunctiva: An analysis in ex-vivo and in-vitro cultures. Lancet Respir. Med. 2020, 2600, 1–9. [Google Scholar] [CrossRef]
- Alexander, S.P.H.; Armstrong, J.F.; Davenport, A.P.; Davies, J.A.; Faccenda, E.; Harding, S.D.; Levi-Schaffer, F.; Maguire, J.J.; Pawson, A.J.; Southan, C.; et al. A rational roadmap for SARS-CoV-2/COVID-19 pharmacotherapeutic research and development: IUPHAR Review 29. Br. J. Pharmacol. 2020, 177, 4942–4966. [Google Scholar] [CrossRef]
- Pena, L.J.; Guarines, K.M.; Duarte Silva, A.J.; Sales Leal, L.R.; Félix, D.M.; Silva, A.; De Oliveira, S.A.; Junqueira Ayres, C.F.; Silva JúNior, A.; De Freitas, A.C. In vitro and in vivo models for studying Zika virus biology. J. Gen. Virol. 2018, 99, 1529–1550. [Google Scholar] [CrossRef]
- Ren, X.; Glende, J.; Al-Falah, M.; de Vries, V.; Schwegmann-Wessels, C.; Qu, X.; Tan, L.; Tschernig, T.; Deng, H.; Naim, H.Y.; et al. Analysis of ACE2 in polarized epithelial cells: Surface expression and function as receptor for severe acute respiratory syndrome-associated coronavirus. J. Gen. Virol. 2006, 87, 1691–1695. [Google Scholar] [CrossRef]
- Hoffmann, M.; Kleine-Weber, H.; Schroeder, S.; Krüger, N.; Herrler, T.; Erichsen, S.; Schiergens, T.S.; Herrler, G.; Wu, N.H.; Nitsche, A.; et al. SARS-CoV-2 Cell Entry Depends on ACE2 and TMPRSS2 and Is Blocked by a Clinically Proven Protease Inhibitor. Cell 2020, 181, 271–280.e8. [Google Scholar] [CrossRef]
- Zhou, P.; Yang, X.L.; Wang, X.G.; Hu, B.; Zhang, L.; Zhang, W.; Si, H.R.; Zhu, Y.; Li, B.; Huang, C.L.; et al. A pneumonia outbreak associated with a new coronavirus of probable bat origin. Nature 2020, 579, 270–273. [Google Scholar] [CrossRef] [Green Version]
- Park, W.B.; Kwon, N.J.; Choi, S.J.; Kang, C.K.; Choe, P.G.; Kim, J.Y.; Yun, J.; Lee, G.W.; Seong, M.W.; Kim, N.J.; et al. Virus isolation from the first patient with SARS-CoV-2 in Korea. J. Korean Med. Sci. 2020, 35, 10–14. [Google Scholar] [CrossRef] [PubMed]
- Harcourt, J.; Tamin, A.; Lu, X.; Kamili, S.; Sakthivel, S.K.; Murray, J.; Queen, K.; Tao, Y.; Paden, C.R.; Zhang, J.; et al. Severe acute respiratory syndrome coronavirus 2 from patient with coronavirus disease, United States. Emerg. Infect. Dis. 2020, 26, 1266–1273. [Google Scholar] [CrossRef] [PubMed]
- Araujo, D.B.; Machado, R.R.G.; Amgarten, D.E.; Malta, F.d.M.; de Araujo, G.G.; Monteiro, C.O.; Candido, E.D.; Soares, C.P.; de Menezes, F.G.; Pires, A.C.C.; et al. SARS-CoV-2 isolation from the first reported patients in Brazil and establishment of a coordinated task network. Mem. Inst. Oswaldo Cruz 2020, 115, e200342. [Google Scholar] [CrossRef]
- Milewska, A.; Kula-Pacurar, A.; Wadas, J.; Suder, A.; Szczepanski, A.; Dabrowska, A.; Owczarek, K.; Marcello, A.; Ochman, M.; Stacel, T.; et al. Replication of Severe Acute Respiratory Syndrome Coronavirus 2 in Human Respiratory Epithelium. J. Virol. 2020, 94, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Klimstra, W.B.; Tilston-Lunel, N.L.; Nambulli, S.; Boslett, J.; McMillen, C.M.; Gilliland, T.; Dunn, M.D.; Sun, C.; Wheeler, S.E.; Wells, A.; et al. SARS-CoV-2 growth, furin-cleavage-site adaptation and neutralization using serum from acutely infected hospitalized COVID-19 patients. J. Gen. Virol. 2020, 101, 1156–1169. [Google Scholar] [CrossRef]
- Liu, Z.; Zheng, H.; Lin, H.; Li, M.; Yuan, R.; Peng, J.; Xiong, Q.; Sun, J.; Li, B.; Wu, J.; et al. Identification of common deletions in the spike protein of SARS-CoV-2. J. Virol. 2020, 1–9. [Google Scholar] [CrossRef]
- Hoehl, S.; Rabenau, H.; Berger, A.; Kortenbusch, M.; Cinatl, J.; Bojkova, D.; Behrens, P.; Böddinghaus, B.; Götsch, U.; Naujoks, F.; et al. Evidence of SARS-CoV-2 infection in returning travelers from Wuhan, China. N. Engl. J. Med. 2020, 382, 1278–1280. [Google Scholar] [CrossRef] [PubMed]
- Bojkova, D.; Klann, K.; Koch, B.; Widera, M.; Krause, D.; Ciesek, S.; Cinatl, J.; Münch, C. Proteomics of SARS-CoV-2-infected host cells reveals therapy targets. Nature 2020. [Google Scholar] [CrossRef]
- Daniloski, Z.; Jordan, T.X.; Wessels, H.H.; Hoagland, D.A.; Kasela, S.; Legut, M.; Maniatis, S.; Mimitou, E.P.; Lu, L.; Geller, E.; et al. Identification of Required Host Factors for SARS-CoV-2 Infection in Human Cells. Cell 2020, 92–105. [Google Scholar] [CrossRef] [PubMed]
- Fan, H.H.; Wang, L.Q.; Liu, W.L.; An, X.P.; Liu, Z.D.; He, X.Q.; Song, L.H.; Tong, Y.G. Repurposing of clinically approved drugs for treatment of coronavirus disease 2019 in a 2019-novel coronavirus-related coronavirus model. Chin. Med. J. (Engl). 2020, 133, 1051–1056. [Google Scholar] [CrossRef]
- Ma, C.; Sacco, M.D.; Hurst, B.; Townsend, J.A.; Hu, Y.; Szeto, T.; Zhang, X.; Tarbet, B.; Marty, M.T.; Chen, Y.; et al. Boceprevir, GC-376, and calpain inhibitors II, XII inhibit SARS-CoV-2 viral replication by targeting the viral main protease. Cell Res. 2020. [Google Scholar] [CrossRef] [PubMed]
- Swaim, C.D.; Perng, Y.-C.; Zhao, X.; Canadeo, L.A.; Harastani, H.H.; Darling, T.L.; Boon, A.C.M.; Lenschow, D.J.; Huibregtse, J.M. 6-Thioguanine blocks SARS-CoV-2 replication by inhibition of PLpro protease activities. bioRxiv Prepr. Serv. Biol. 2020, 1–20. [Google Scholar] [CrossRef]
- Pruijssers, A.J.; George, A.S.; Schäfer, A.; Leist, S.R.; Gralinksi, L.E.; Dinnon, K.H.; Yount, B.L.; Agostini, M.L.; Stevens, L.J.; Chappell, J.D.; et al. Remdesivir Inhibits SARS-CoV-2 in Human Lung Cells and Chimeric SARS-CoV Expressing the SARS-CoV-2 RNA Polymerase in Mice. Cell Rep. 2020, 32. [Google Scholar] [CrossRef]
- Caly, L.; Druce, J.D.; Catton, M.G.; Jans, D.A.; Wagstaff, K.M. The FDA-approved drug ivermectin inhibits the replication of SARS-CoV-2 in vitro. Antiviral Res. 2020, 178, 3–6. [Google Scholar] [CrossRef]
- Yao, X.; Ye, F.; Zhang, M.; Cui, C.; Huang, B.; Niu, P.; Liu, X.; Zhao, L.; Dong, E.; Song, C.; et al. In Vitro Antiviral Activity and Projection of Optimized Dosing Design of Hydroxychloroquine for the Treatment of Severe Acute Respiratory Syndrome Coronavirus 2 (SARS-CoV-2). Clin. Infect. Dis. 2020, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Kaur, H.; Shekhar, N.; Sharma, S.; Sarma, P.; Prakash, A.; Medhi, B. Ivermectin as a potential drug for treatment of COVID-19: An in-sync review with clinical and computational attributes. Pharmacol. Reports 2021. [Google Scholar] [CrossRef]
- Sands, K.; Wenzel, R.; McLean, L.; Korwek, K.; Roach, J.; Miller, K.; Poland, R.E.; Burgess, L.H.; Jackson, E.; Perlin, J.B. No clinical benefit in mortality associated with hydroxychloroquine treatment in patients with COVID-19. Int. J. Infect. Dis. 2021, 104, 34–40. [Google Scholar] [CrossRef]
- Banach, B.S.; Orenstein, J.M.; Fox, L.M.; Randell, S.H.; Rowley, A.H.; Baker, S.C. Human airway epithelial cell culture to identify new respiratory viruses: Coronavirus NL63 as a model. J. Virol. Methods 2009, 156, 19–26. [Google Scholar] [CrossRef]
- Jonsdottir, H.R.; Dijkman, R. Coronaviruses and the human airway: A universal system for virus-host interaction studies Coronaviruses: Emerging and re-emerging pathogens in humans and animals Susanna Lau Emerging viruses. Virol. J. 2016, 13, 1–9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, X.; Wu, Y.; Rong, L. Conditionally Reprogrammed Human Normal Airway Epithelial Cells at ALI: A Physiological Model for Emerging Viruses. Virol. Sin. 2020, 12250, 1–10. [Google Scholar] [CrossRef]
- Hou, Y.J.; Okuda, K.; Edwards, C.E.; Martinez, D.R.; Asakura, T.; Dinnon, K.H.; Kato, T.; Lee, R.E.; Yount, B.L.; Mascenik, T.M.; et al. SARS-CoV-2 Reverse Genetics Reveals a Variable Infection Gradient in the Respiratory Tract. Cell 2020, 1–18. [Google Scholar] [CrossRef]
- Shang, J.; Wan, Y.; Luo, C.; Ye, G.; Geng, Q.; Auerbach, A.; Li, F. Cell entry mechanisms of SARS-CoV-2. Proc. Natl. Acad. Sci. USA 2020, 117. [Google Scholar] [CrossRef]
- Abo, K.M.; Ma, L.; Matte, T.; Huang, J.; Alysandratos, K.D.; Werder, R.B.; Mithal, A.; Beermann, M.L.; Lindstrom-Vautrin, J.; Mostoslavsky, G.; et al. Human iPSC-derived alveolar and airway epithelial cells can be cultured at air-liquid interface and express SARS-CoV-2 host factors. bioRxiv 2020. [Google Scholar] [CrossRef]
- Sungnak, W.; Huang, N.; Bécavin, C.; Berg, M.; Queen, R.; Litvinukova, M.; Talavera-López, C.; Maatz, H.; Reichart, D.; Sampaziotis, F.; et al. SARS-CoV-2 entry factors are highly expressed in nasal epithelial cells together with innate immune genes. Nat. Med. 2020, 26, 681–687. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stelzig, K.E.; Canepa-Escaro, F.; Schiliro, M.; Berdnikovs, S.; Prakash, Y.S.; Chiarella, S.E. Estrogen regulates the expression of SARS-CoV-2 receptor ACE2 in differentiated airway epithelial cells. Am. J. Physiol. Lung Cell. Mol. Physiol. 2020, 318, L1280–L1281. [Google Scholar] [CrossRef] [PubMed]
- Blanco-Melo, D.; Nilsson-Payant, B.E.; Liu, W.C.; Uhl, S.; Hoagland, D.; Møller, R.; Jordan, T.X.; Oishi, K.; Panis, M.; Sachs, D.; et al. Imbalanced Host Response to SARS-CoV-2 Drives Development of COVID-19. Cell 2020, 181, 1036–1045.e9. [Google Scholar] [CrossRef]
- Sheahan, T.P.; Sims, A.C.; Zhou, S.; Graham, R.L.; Pruijssers, A.J.; Agostini, M.L.; Leist, S.R.; Schäfer, A.; Dinnon, K.H.; Stevens, L.J.; et al. An orally bioavailable broad-spectrum antiviral inhibits SARS-CoV-2 in human airway epithelial cell cultures and multiple coronaviruses in mice. Sci. Transl. Med. 2020, 12. [Google Scholar] [CrossRef] [Green Version]
- Clevers, H. COVID-19: Organoids go viral. Nat. Rev. Mol. Cell Biol. 2020, 21, 355–356. [Google Scholar] [CrossRef]
- Qi, F.; Qian, S.; Zhang, S.; Zhang, Z. Single cell RNA sequencing of 13 human tissues identify cell types and receptors of human coronaviruses. Biochem. Biophys. Res. Commun. 2020, 526, 135–140. [Google Scholar] [CrossRef]
- Singh, M.; Bansal, V.; Feschotte, C. A Single-Cell RNA Expression Map of Human Coronavirus Entry Factors. Cell Rep. 2020, 32. [Google Scholar] [CrossRef]
- Chu, H.; Chan, J.F.W.; Wang, Y.; Yuen, T.T.T.; Chai, Y.; Hou, Y.; Shuai, H.; Yang, D.; Hu, B.; Huang, X.; et al. Comparative replication and immune activation profiles of SARS-CoV-2 and SARS-CoV in human lungs: An ex vivo study with implications for the pathogenesis of COVID-19. Clin. Infect. Dis. 2020, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Yang, L.; Han, Y.; Nilsson-Payant, B.E.; Gupta, V.; Wang, P.; Duan, X.; Tang, X.; Zhu, J.; Zhao, Z.; Jaffré, F.; et al. A Human Pluripotent Stem Cell-based Platform to Study SARS-CoV-2 Tropism and Model Virus Infection in Human Cells and Organoids. Cell Stem Cell 2020, 27, 125–136.e7. [Google Scholar] [CrossRef] [PubMed]
- Han, Y.; Duan, X.; Yang, L.; Nilsson-Payant, B.E.; Wang, P.; Duan, F.; Tang, X.; Yaron, T.M.; Zhang, T.; Uhl, S.; et al. Identification of SARS-CoV-2 Inhibitors using Lung and Colonic Organoids. Nature 2020, 589, 270–275. [Google Scholar] [CrossRef] [PubMed]
- Matschke, J.; Lütgehetmann, M.; Hagel, C.; Sperhake, J.P.; Schröder, A.S.; Edler, C.; Mushumba, H.; Fitzek, A.; Allweiss, L.; Dandri, M.; et al. Neuropathology of patients with COVID-19 in Germany: A post-mortem case series. Lancet Neurol. 2020, 19, 919–929. [Google Scholar] [CrossRef]
- Ramani, A.; Müller, L.; Ostermann, P.N.; Gabriel, E.; Abida--Islam, P.; Müller--Schiffmann, A.; Mariappan, A.; Goureau, O.; Gruell, H.; Walker, A.; et al. SARS --CoV--2 targets neurons of 3D human brain organoids. EMBO J. 2020, 39, 1–14. [Google Scholar] [CrossRef]
- Zhang, B.Z.; Chu, H.; Han, S.; Shuai, H.; Deng, J.; Hu, Y.F.; Gong, H.R.; Lee, A.C.Y.; Zou, Z.; Yau, T.; et al. SARS-CoV-2 infects human neural progenitor cells and brain organoids. Cell Res. 2020, 30, 928–931. [Google Scholar] [CrossRef]
- Leonardi, A.; Rosani, U.; Brun, P. Ocular Surface Expression of SARS-CoV-2 Receptors. Ocul. Immunol. Inflamm. 2020, 28, 735–738. [Google Scholar] [CrossRef]
- Wu, P.; Duan, F.; Luo, C.; Liu, Q.; Qu, X.; Liang, L.; Wu, K. Characteristics of Ocular Findings of Patients with Coronavirus Disease 2019 (COVID-19) in Hubei Province, China. JAMA Ophthalmol. 2020, 138, 575–578. [Google Scholar] [CrossRef]
- Seah, I.Y.J.; Anderson, D.E.; Kang, A.E.Z.; Wang, L.; Rao, P.; Young, B.E.; Lye, D.C.; Agrawal, R. Assessing Viral Shedding and Infectivity of Tears in Coronavirus Disease 2019 (COVID-19) Patients. Ophthalmology 2020, 127, 977–979. [Google Scholar] [CrossRef] [PubMed]
- Makovoz, B.; Moeller, R.; Zebitz Eriksen, A.; tenOever, B.R.; Blenkinsop, T.A. SARS-CoV-2 Infection of Ocular Cells from Human Adult Donor Eyes and hESC-Derived Eye Organoids. Ssrn 2020, 3650574. [Google Scholar] [CrossRef]
- Monteil, V.; Kwon, H.; Prado, P.; Hagelkrüys, A.; Wimmer, R.A.; Stahl, M.; Leopoldi, A.; Garreta, E.; Hurtado del Pozo, C.; Prosper, F.; et al. Inhibition of SARS-CoV-2 Infections in Engineered Human Tissues Using Clinical-Grade Soluble Human ACE2. Cell 2020, 181, 905–913.e7. [Google Scholar] [CrossRef] [PubMed]
- Yuan, L.; Tang, Q.; Cheng, T.; Xia, N. Animal models for emerging coronavirus: Progress and new insights. Emerg. Microbes Infect. 2020, 9, 949–961. [Google Scholar] [CrossRef]
- Muñoz-Fontela, C.; Dowling, W.E.; Funnell, S.G.P.; Gsell, P.S.; Riveros-Balta, A.X.; Albrecht, R.A.; Andersen, H.; Baric, R.S.; Carroll, M.W.; Cavaleri, M.; et al. Animal models for COVID-19. Nature 2020, 586, 509–515. [Google Scholar] [CrossRef] [PubMed]
- Wan, Y.; Shang, J.; Graham, R.; Baric, R.S.; Li, F. Receptor Recognition by the Novel Coronavirus from Wuhan: An Analysis Based on Decade-Long Structural Studies of SARS Coronavirus. J. Virol. 2020, 94, 1–9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Frieman, M.; Yount, B.; Agnihothram, S.; Page, C.; Donaldson, E.; Roberts, A.; Vogel, L.; Woodruff, B.; Scorpio, D.; Subbarao, K.; et al. Molecular Determinants of Severe Acute Respiratory Syndrome Coronavirus Pathogenesis and Virulence in Young and Aged Mouse Models of Human Disease. J. Virol. 2012, 86, 884–897. [Google Scholar] [CrossRef] [Green Version]
- Menachery, V.D.; Yount, B.L.; Sims, A.C.; Debbink, K.; Agnihothram, S.S.; Gralinski, L.E.; Graham, R.L.; Scobey, T.; Plante, J.A.; Royal, S.R.; et al. SARS-like WIV1-CoV poised for human emergence. Proc. Natl. Acad. Sci. USA 2016, 113, 3048–3053. [Google Scholar] [CrossRef] [Green Version]
- Roberts, A.; Deming, D.; Paddock, C.D.; Cheng, A.; Yount, B.; Vogel, L.; Herman, B.D.; Sheahan, T.; Heise, M.; Genrich, G.L.; et al. A mouse-adapted SARS-coronavirus causes disease and mortality in BALB/c mice. PLoS Pathog. 2007, 3, 0023–0037. [Google Scholar] [CrossRef]
- Tseng, C.-T.K.; Huang, C.; Newman, P.; Wang, N.; Narayanan, K.; Watts, D.M.; Makino, S.; Packard, M.M.; Zaki, S.R.; Chan, T.; et al. Severe Acute Respiratory Syndrome Coronavirus Infection of Mice Transgenic for the Human Angiotensin-Converting Enzyme 2 Virus Receptor. J. Virol. 2007, 81, 1162–1173. [Google Scholar] [CrossRef] [Green Version]
- Yang, X.H.; Deng, W.; Tong, Z.; Liu, Y.X.; Zhang, L.F.; Zhu, H.; Gao, H.; Huang, L.; Liu, Y.I.; Ma, C.M.; et al. Mice transgenic for human angiotensin-converting enzyme 2 provide a model for SARS coronavirus infection. Comp. Med. 2007, 57, 450–459. [Google Scholar] [PubMed]
- Sun, S.H.; Chen, Q.; Gu, H.J.; Yang, G.; Wang, Y.X.; Huang, X.Y.; Liu, S.S.; Zhang, N.N.; Li, X.F.; Xiong, R.; et al. A Mouse Model of SARS-CoV-2 Infection and Pathogenesis. Cell Host Microbe 2020, 1–10. [Google Scholar] [CrossRef]
- Ostrowski, L.E.; Hutchins, J.R.; Zakel, K.; O’Neal, W.K. Targeting expression of a transgene to the airway surface epithelium using a ciliated cell-specific promoter. Mol. Ther. 2003, 8, 637–645. [Google Scholar] [CrossRef]
- Jiang, R.D.; Liu, M.Q.; Chen, Y.; Shan, C.; Zhou, Y.W.; Shen, X.R.; Li, Q.; Zhang, L.; Zhu, Y.; Si, H.R.; et al. Pathogenesis of SARS-CoV-2 in Transgenic Mice Expressing Human Angiotensin-Converting Enzyme 2. Cell 2020, 1–9. [Google Scholar] [CrossRef]
- Bao, L.; Deng, W.; Huang, B.; Gao, H.; Liu, J.; Ren, L.; Wei, Q.; Yu, P.; Xu, Y.; Qi, F.; et al. The pathogenicity of SARS-CoV-2 in hACE2 transgenic mice. Nature 2020. [Google Scholar] [CrossRef]
- Shulla, A.; Heald-Sargent, T.; Subramanya, G.; Zhao, J.; Perlman, S.; Gallagher, T. A Transmembrane Serine Protease Is Linked to the Severe Acute Respiratory Syndrome Coronavirus Receptor and Activates Virus Entry. J. Virol. 2011, 85, 873–882. [Google Scholar] [CrossRef] [Green Version]
- Soldatov, V.O.; Kubekina, M.V.; Silaeva, Y.Y.; Bruter, A.V.; Deykin, A.V. On the way from SARS-CoV-sensitive mice to murine COVID-19 model. Res. Results Pharmacol. 2020, 6, 1–7. [Google Scholar] [CrossRef]
- Dinnon, K.H.; Leist, S.R.; Schäfer, A.; Edwards, C.E.; Martinez, D.R.; Montgomery, S.A.; West, A.; Yount, B.L.; Hou, Y.J.; Adams, L.E.; et al. A mouse-adapted SARS-CoV-2 model for the evaluation of COVID-19 medical countermeasures. bioRxiv 2020. [Google Scholar] [CrossRef]
- Dinnon, K.H.; Leist, S.R.; Schäfer, A.; Edwards, C.E.; Martinez, D.R.; Montgomery, S.A.; West, A.; Yount, B.L.; Hou, Y.J.; Adams, L.E.; et al. A mouse-adapted model of SARS-CoV-2 to test COVID-19 countermeasures. Nature 2020, 586, 560–566. [Google Scholar] [CrossRef] [PubMed]
- Hassan, A.O.; Case, J.B.; Winkler, E.S.; Thackray, L.B.; Kafai, N.M.; Bailey, A.L.; McCune, B.T.; Fox, J.M.; Chen, R.E.; Alsoussi, W.B.; et al. A SARS-CoV-2 Infection Model in Mice Demonstrates Protection by Neutralizing Antibodies. Cell 2020, 182, 744–753.e4. [Google Scholar] [CrossRef]
- Israelow, B.; Song, E.; Mao, T.; Lu, P.; Meir, A.; Liu, F.; Alfajaro, M.M.; Wei, J.; Dong, H.; Homer, R.J.; et al. Mouse model of SARS-CoV-2 reveals inflammatory role of type i interferon signaling. J. Exp. Med. 2020, 217. [Google Scholar] [CrossRef]
- Zhang, Y.N.; Li, X.D.; Zhang, Z.R.; Zhang, H.Q.; Li, N.; Liu, J.; Li, J.Q.; Zhang, H.J.; Wang, Z.J.; Shen, S.; et al. A mouse model for SARS-CoV-2 infection by exogenous delivery of hACE2 using alphavirus replicon particles. Cell Res. 2020, 30, 1046–1048. [Google Scholar] [CrossRef]
- Rathnasinghe, R.; Strohmeier, S.; Amanat, F.; Gillespie, V.L.; Krammer, F.; García-Sastre, A.; Coughlan, L.; Schotsaert, M.; Uccellini, M. Comparison of Transgenic and Adenovirus hACE2 Mouse Models for SARS-CoV-2 Infection. Emerg. Microbes Infect. 2020, 1–19. [Google Scholar] [CrossRef]
- Gu, H.; Chen, Q.; Yang, G.; He, L.; Fan, H.; Deng, Y.Q.; Wang, Y.; Teng, Y.; Zhao, Z.; Cui, Y.; et al. Adaptation of SARS-CoV-2 in BALB/c mice for testing vaccine efficacy. Science 2020, 369, 1603–1607. [Google Scholar] [CrossRef]
- Damas, J.; Hughes, G.M.; Keough, K.C.; Painter, C.A.; Persky, N.S.; Corbo, M.; Hiller, M.; Koepfli, K.P.; Pfenning, A.R.; Zhao, H.; et al. Broad host range of SARS-CoV-2 predicted by comparative and structural analysis of ACE2 in vertebrates. Proc. Natl. Acad. Sci. USA 2020, 117, 22311–22322. [Google Scholar] [CrossRef] [PubMed]
- Chan, J.F.; Zhang, A.J.; Yuan, S.; Kwok-, V. Simulation of the clinical and pathological manifestations of Coronavirus Disease 2019 (COVID-19) in golden Syrian hamster model: Implications for disease pathogenesis and transmissibility. Clin Infect Dis. 2019, 1–50. [Google Scholar] [CrossRef] [Green Version]
- Sia, S.F.; Yan, L.M.; Chin, A.W.H.; Fung, K.; Choy, K.T.; Wong, A.Y.L.; Kaewpreedee, P.; Perera, R.A.P.M.; Poon, L.L.M.; Nicholls, J.M.; et al. Pathogenesis and transmission of SARS-CoV-2 in golden hamsters. Nature 2020. [Google Scholar] [CrossRef] [PubMed]
- Imai, M.; Iwatsuki-Horimoto, K.; Hatta, M.; Loeber, S.; Halfmann, P.J.; Nakajima, N.; Watanabe, T.; Ujie, M.; Takahashi, K.; Ito, M.; et al. Syrian hamsters as a small animal model for SARS-CoV-2 infection and countermeasure development. Proc. Natl. Acad. Sci. USA 2020, 117, 16587–16595. [Google Scholar] [CrossRef] [PubMed]
- Boudewijns, R.; Thibaut, H.J.; Kaptein, S.J.F.; Li, R.; Vergote, V.; Seldeslachts, L.; Van Weyenbergh, J.; De Keyzer, C.; Bervoets, L.; Sharma, S.; et al. STAT2 signaling restricts viral dissemination but drives severe pneumonia in SARS-CoV-2 infected hamsters. Nat. Commun. 2020, 11, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Osterrieder, N.; Bertzbach, L.D.; Dietert, K.; Abdelgawad, A.; Vladimirova, D.; Kunec, D.; Hoffmann, D.; Beer, M.; Gruber, A.D.; Trimpert, J. Age-dependent progression of SARS-CoV-2 infection in Syrian hamsters. Viruses 2020, 12, 779. [Google Scholar] [CrossRef]
- Chan, J.F.-W.; Yuan, S.; Zhang, A.J.; Poon, V.K.-M.; Chan, C.C.-S.; Lee, A.C.-Y.; Fan, Z.; Li, C.; Liang, R.; Cao, J.; et al. Surgical Mask Partition Reduces the Risk of Noncontact Transmission in a Golden Syrian Hamster Model for Coronavirus Disease 2019 (COVID-19). Clin. Infect. Dis. 2020, 2019, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Mohandas, S.; Shete, A.; Abraham, P.; Mohan, K. Immunogenicity and protective e cacy of BBV152: A whole virion inactivated SARS CoV-2 vaccine in the Syrian hamster model. Res. Sq. 2020, 1–25. [Google Scholar]
- Vitner, E.B.; Israeli, O.; Milrot, E.; Stein, D.; Cohen-gihon, I.; Lazar, S.; Gutman, H.; Lupu, E.; David, E.B.; Sittner, A.; et al. A single dose of recombinant VSV-∆G-spike vaccine provides protection against SARS-CoV-2 challenge Yfat. Nat. Commun. 2020, 11, 6402. [Google Scholar]
- Tostanoski, L.H.; Wegmann, F.; Martinot, A.J.; Loos, C.; McMahan, K.; Mercado, N.B.; Yu, J.; Chan, C.N.; Bondoc, S.; Starke, C.E.; et al. Ad26 vaccine protects against SARS-CoV-2 severe clinical disease in hamsters. Nat. Med. 2020. [Google Scholar] [CrossRef] [PubMed]
- Kaptein, S.J.F.; Jacobs, S.; Langendries, L.; Seldeslachts, L.; ter Horst, S.; Liesenborghs, L.; Hens, B.; Vergote, V.; Heylen, E.; Barthelemy, K.; et al. Favipiravir at high doses has potent antiviral activity in SARS-CoV-2−infected hamsters, whereas hydroxychloroquine lacks activity. Proc. Natl. Acad. Sci. USA 2020, 117, 26955–26965. [Google Scholar] [CrossRef]
- Yuan, S.; Wang, R.; Chan, J.F.W.; Zhang, A.J.; Cheng, T.; Chik, K.K.H.; Ye, Z.W.; Wang, S.; Lee, A.C.Y.; Jin, L.; et al. Metallodrug ranitidine bismuth citrate suppresses SARS-CoV-2 replication and relieves virus-associated pneumonia in Syrian hamsters. Nat. Microbiol. 2020, 5, 1439–1448. [Google Scholar] [CrossRef]
- Yuan, S.; Yin, X.; Meng, X.; Chan, J.; Ye, Z.-W.; Riva, L.; Pache, L.; Chan, C.C.-Y.; Lai, P.-M.; Chan, C.; et al. Clofazimine is a broad-spectrum coronavirus inhibitor that antagonizes SARS-CoV-2 replication in primary human cell culture and hamsters. Res. Sq. 2020, 1–26. [Google Scholar] [CrossRef]
- Sun, X.; Meyerholz, D.K.; Engelhardt, J.F.; Sun, X.; Sui, H.; Fisher, J.T.; Yan, Z.; Liu, X.; Cho, H.; Joo, N.S.; et al. Disease phenotype of a ferret CFTR-knockout model of cystic fibrosis. J. Clin. Investig. 2010, 120, 3149–3160. [Google Scholar] [CrossRef] [Green Version]
- Guan, W.; Ni, Z.; Hu, Y.; Liang, W.; Ou, C.; He, J.; Liu, L.; Shan, H.; Lei, C.; Hui, D.S.C.; et al. Clinical characteristics of coronavirus disease 2019 in China. N. Engl. J. Med. 2020, 382, 1708–1720. [Google Scholar] [CrossRef] [PubMed]
- Park, S.J.; Yu, K.M.; Kim, Y.I.; Kim, S.M.; Kim, E.H.; Kim, S.G.; Kim, E.J.; Casel, M.A.B.; Rollon, R.; Jang, S.G.; et al. Antiviral efficacies of FDA-approved drugs against SARS-COV-2 infection in ferrets. MBio 2020, 11, 1–10. [Google Scholar] [CrossRef]
- Wu, S.; Zhong, G.; Zhang, J.; Shuai, L.; Zhang, Z.; Wen, Z.; Wang, B.; Zhao, Z.; Song, X.; Chen, Y.; et al. A single dose of an adenovirus-vectored vaccine provides protection against SARS-CoV-2 challenge. Nat. Commun. 2020, 11. [Google Scholar] [CrossRef]
- Shi, J.; Wen, Z.; Zhong, G.; Yang, H.; Wang, C.; Huang, B.; Liu, R.; He, X.; Shuai, L.; Sun, Z.; et al. Susceptibility of ferrets, cats, dogs, and other domesticated animals to SARS-coronavirus 2. Science 2020, 368, 1016–1020. [Google Scholar] [CrossRef] [Green Version]
- Ryan, K.A.; Bewley, K.R.; Fotheringham, S.A.; Brown, P.; Hall, Y.; Marriott, A.C.; Tree, J.A.; Allen, L.; Aram, M.J.; Brunt, E.; et al. Dose-dependent response to infection with SARS-CoV-2 in the ferret model: Evidence of protection to re-challenge. bioRxiv 2020, 2. [Google Scholar] [CrossRef]
- Richard, M.; Kok, A.; de Meulder, D.; Bestebroer, T.M.; Lamers, M.M.; Okba, N.M.A.; van Vlissingen, M.F.; Rockx, B.; Haagmans, B.L.; Marion, P.G.; et al. SARS-CoV-2 is transmitted via contact and via the air between ferrets. Nat. Commun. 2020, 11, 3496. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.I.; Kim, S.G.; Kim, S.M.; Kim, E.H.; Park, S.J.; Yu, K.M.; Chang, J.H.; Kim, E.J.; Lee, S.; Casel, M.A.B.; et al. Infection and Rapid Transmission of SARS-CoV-2 in Ferrets. Cell Host Microbe 2020, 27, 704–709.e2. [Google Scholar] [CrossRef] [PubMed]
- Mathavarajah, S.; Dellaire, G. Lions, tigers and kittens too: ACE2 and susceptibility to COVID-19. Evol. Med. Public Heal. 2020, 2020, 109–113. [Google Scholar] [CrossRef] [PubMed]
- Sit, T.H.C.; Brackman, C.J.; Ip, S.M.; Tam, K.W.S.; Law, P.Y.T.; To, E.M.W.; Yu, V.Y.T.; Sims, L.D.; Tsang, D.N.C.; Chu, D.K.W.; et al. Infection of dogs with SARS-CoV-2. Nature 2020. [Google Scholar] [CrossRef]
- Bosco-Lauth, A.M.; Hartwig, A.E.; Porter, S.M.; Gordy, P.W.; Nehring, M.; Byas, A.D.; Vandewoude, S.; Ragan, I.K.; Maison, R.M.; Bowen, R.A. Pathogenesis, transmission and response to re-exposure of SARS-CoV-2 in domestic cats. BioRxiv 2020, 21, 1–9. [Google Scholar] [CrossRef]
- Bosco-Lauth, A.M.; Hartwig, A.E.; Porter, S.M.; Gordy, P.W.; Nehring, M.; Byas, A.D.; VandeWoude, S.; Ragan, I.K.; Maison, R.M.; Bowen, R.A. Experimental infection of domestic dogs and cats with SARS-CoV-2: Pathogenesis, transmission, and response to reexposure in cats. Proc. Natl. Acad. Sci. USA 2020, 117, 26382–26388. [Google Scholar] [CrossRef] [PubMed]
- Fritz, M.; Rosolen, B.; Krafft, E.; Becquart, P.; Elguero, E.; Vratskikh, O.; Denolly, S.; Boson, B.; Vanhomwegen, J. High prevalence of SARS-CoV-2 antibodies in pets from COVID-19+ households. One Health 2020, 11, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Zhang, H.; Huang, K.; Yang, Y.; Hui, X.; Gao, J.; He, X.; Li, C.; Gong, W.; Zhang, Y.; et al. SARS-CoV-2 neutralizing serum antibodies in cats: A serological investigation. bioRxiv 2020. [Google Scholar] [CrossRef] [Green Version]
- Halfmann, P.J.; Hatta, M.; Chiba, S.; Maemura, T.; Fan, S.; Takeda, M.; Kinoshita, N.; Hattori, S.; Sakai-Tagawa, Y.; Iwatsuki-Horimoto, K.; et al. Transmission of SARS-CoV-2 in Domestic Cats. N. Engl. J. Med. 2020. [Google Scholar] [CrossRef]
- Rockx, B.; Kuiken, T.; Herfst, S.; Bestebroer, T.; Lamers, M.M.; Oude Munnink, B.B.; de Meulder, D.; van Amerongen, G.; van den Brand, J.; Okba, N.M.A.; et al. Comparative pathogenesis of COVID-19, MERS, and SARS in a nonhuman primate model. Science 2020, 368, 1012–1015. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Munster, V.J.; Feldmann, F.; Williamson, B.N.; van Doremalen, N.; Pérez-Pérez, L.; Schulz, J.; Meade-White, K.; Okumura, A.; Callison, J.; Brumbaugh, B.; et al. Respiratory disease in rhesus macaques inoculated with SARS-CoV-2. Nature 2020. [Google Scholar] [CrossRef]
- Lu, S.; Zhao, Y.; Yu, W.; Yang, Y.; Gao, J.; Wang, J.; Kuang, D.; Yang, M.; Yang, J.; Ma, C.; et al. Comparison of nonhuman primates identified the suitable model for COVID-19. Signal Transduct. Target. Ther. 2020, 5. [Google Scholar] [CrossRef]
- Hartman, A.L.; Nambulli, S.; McMillen, C.M.; White, A.G.; Tilston-Lunel, N.L.; Albe, J.R.; Cottle, E.; Dunn, M.D.; James Frye, L.; Gilliland, T.H.; et al. SARS-CoV-2 infection of African green monkeys results in mild respiratory disease discernible by PET/CT imaging and shedding of infectious virus from both respiratory and gastrointestinal tracts. PLoS Pathog. 2020, 16, 1–24. [Google Scholar] [CrossRef]
- Curtis, M.J.; Alexander, S.; Cirino, G.; Docherty, J.R.; George, C.H.; Giembycz, M.A.; Hoyer, D.; Insel, P.A.; Izzo, A.A.; Ji, Y.; et al. Experimental design and analysis and their reporting II: Updated and simplified guidance for authors and peer reviewers. Br. J. Pharmacol. 2018, 175, 987–993. [Google Scholar] [CrossRef] [Green Version]
- Deng, W.; Bao, L.; Gao, H.; Xiang, Z.; Qu, Y.; Song, Z.; Gong, S.; Liu, J.; Liu, J.; Yu, P.; et al. Ocular conjunctival inoculation of SARS-CoV-2 can cause mild COVID-19 in rhesus macaques. Nat. Commun. 2020, 11, 1–7. [Google Scholar] [CrossRef]
- Yu, P.; Qi, F.; Xu, Y.; Li, F.; Liu, P.; Liu, J.; Bao, L.; Deng, W.; Gao, H.; Xiang, Z.; et al. Age--related rhesus macaque models of COVID--19. Anim. Model. Exp. Med. 2020, 3, 93–97. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Deng, W.; Bao, L.; Liu, J.; Xiao, C.; Liu, J.; Xue, J.; Lv, Q.; Qi, F.; Gao, H.; Yu, P.; et al. Primary exposure to SARS-CoV-2 protects against reinfection in rhesus macaques. Science 2020, 369, 818–823. [Google Scholar] [CrossRef]
- Chandrashekar, A.; Liu, J.; Martino, A.J.; McMahan, K.; Mercad, N.B.; Peter, L.; Tostanosk, L.H.; Yu, J.; Maliga, Z.; Nekorchuk, M.; et al. SARS-CoV-2 infection protects against rechallenge in rhesus macaques. Science 2020, 369, 812–817. [Google Scholar] [CrossRef]
- Yu, J.; Tostanosk, L.H.; Peter, L.; Mercad, N.B.; McMahan, K.; Mahrokhia, S.H.; Nkolol, J.P.; Liu, J.; Li, Z.; Chandrashekar, A.; et al. DNA vaccine protection against SARS-CoV-2 in rhesus macaques. Science 2020, 369, 806–811. [Google Scholar] [CrossRef]
- Wang, H.; Zhang, Y.; Huang, B.; Deng, W.; Quan, Y.; Wang, W.; Xu, W.; Zhao, Y.; Li, N.; Zhang, J.; et al. Development of an Inactivated Vaccine Candidate, BBIBP-CorV, with Potent Protection against SARS-CoV-2. Cell 2020, 182, 713–721.e9. [Google Scholar] [CrossRef] [PubMed]
- Mercado, N.B.; Zahn, R.; Wegmann, F.; Loos, C.; Chandrashekar, A.; Yu, J.; Liu, J.; Peter, L.; McMahan, K.; Tostanoski, L.H.; et al. Single-shot Ad26 vaccine protects against SARS-CoV-2 in rhesus macaques. Nature 2020, 586, 583–588. [Google Scholar] [CrossRef] [PubMed]
- Doremalen, N.; Lambe, T.; Spencer, A.; Belij-Rammerstorfer, S.; Purushotham, J.N.; Port, J.R.; Avanzato, V.A.; Bushmaker, T.; Flaxman, A.; Ulaszewska, M.; et al. ChAdOx1 nCoV-19 vaccine prevents SARS-CoV-2 pneumonia in rhesus macaques. Nature 2020, 586, 578–582. [Google Scholar] [CrossRef]
- Corbett, K.S.; Flynn, B.; Foulds, K.E.; Francica, J.R.; Boyoglu-Barnum, S.; Werner, A.P.; Flach, B.; O’Connell, S.; Bock, K.W.; Minai, M.; et al. Evaluation of the mRNA-1273 Vaccine against SARS-CoV-2 in Nonhuman Primates. N. Engl. J. Med. 2020, 383, 1544–1555. [Google Scholar] [CrossRef] [PubMed]
- Maisonnasse, P.; Guedj, J.; Contreras, V.; Behillil, S.; Solas, C.; Marlin, R.; Naninck, T.; Pizzorno, A.; Lemaitre, J.; Gonçalves, A.; et al. Hydroxychloroquine use against SARS-CoV-2 infection in non-human primates. Nature 2020, 585, 584–587. [Google Scholar] [CrossRef]
- Williamson, B.N.; Feldmann, F.; Schwarz, B.; Meade-White, K.; Porter, D.P.; Schulz, J.; van Doremalen, N.; Leighton, I.; Yinda, C.K.; Pérez-Pérez, L.; et al. Clinical benefit of remdesivir in rhesus macaques infected with SARS-CoV-2. Nature 2020. [Google Scholar] [CrossRef]
- Schlottau, K.; Rissmann, M.; Graaf, A.; Schön, J.; Sehl, J.; Wylezich, C.; Höper, D.; Mettenleiter, T.C.; Balkema-Buschmann, A.; Harder, T.; et al. SARS-CoV-2 in fruit bats, ferrets, pigs, and chickens: An experimental transmission study. Lancet Microbe 2020, 1, e218–e225. [Google Scholar] [CrossRef]
- Meekins, D.A.; Morozov, I.; Trujillo, J.D.; Gaudreault, N.N.; Bold, D.; Carossino, M.; Artiaga, B.L.; Indran, S.V.; Kwon, T.; Balaraman, V.; et al. Susceptibility of swine cells and domestic pigs to SARS-CoV-2. Emerg. Microbes Infect. 2020, 9, 2278–2288. [Google Scholar] [CrossRef] [PubMed]
- Oreshkova, N.; Molenaar, R.J.; Vreman, S.; Harders, F.; Oude Munnink, B.B.; Van Der Honing, R.W.H.; Gerhards, N.; Tolsma, P.; Bouwstra, R.; Sikkema, R.S.; et al. SARS-CoV-2 infection in farmed minks, the Netherlands, April and May 2020. Eurosurveillance 2020, 25, 1–7. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
| Cell/Tissue Type | SARS-CoV-2 Strain | Viral Cultivation | Main Applications | References |
|---|---|---|---|---|
| Two-dimensional (2D) Models: Immortalized Cells | ||||
| Vero CCL-81 and Vero E6 | 2019-nCoVBetaCoV/Wuhan/WIV04/2019; BetaCoV/Korea/SNU01/2020; SARS-CoV-2/USA-WA1/2020; Australia/VIC01/2020; C-Tan-nCoV Wuhan strain 01 | Vero cells used in viral isolation were cultured as monolayers in flasks at 37 °C and 5% atm of carbon dioxide (CO2) in Dulbecco’s Modified Eagle Medium (DMEM) or Earle’s Minimum Essential Medium (EMEM) supplemented with 2 to 10% fetal bovine serum (FBS). Some authors cite extra supplementation with antibiotics and buffers. Cytopathic effect could be observed in 2 to 6 days of infection (d.p.i.) or after another viral passage. Protocols for antiviral assays include cultivation in microplates with different values for multiplicity of infection (MOI). | Virus isolation of different SARS-CoV-2 strains; infection characterization; evaluation of host responses; evaluation of antiviral activity; vaccine production. | [15,16,17,18,19,25,26,27,28,29,30] |
| Huh-7 | 2019-nCoV BetaCoV/Wuhan/WIV04/2019; SARS-CoV-2/USA-WA1/2020 | Huh-7 cells were cultured in DMEM supplemented with 10% FBS at 37 °C and 5% atm CO2. Some authors have reported supplementation with antibiotics and antimycotics. For antiviral assays, MOI 0.01 was used, and the supernatant was collected after 48 h. | SARS-CoV-2 isolation; infection characterization; evaluation of host responses; evaluation of antiviral activity. | [15,28] |
| Caco-2 | BetaCoV/Hong Kong/VM20001061/2020 | Caco-2 cells were maintained at 37 °C and 5% atm of CO2 and cultured with MEM or DMEM supplemented with 10% FBS. Some authors have reported supplementation with antibiotics and antimycotics. SARS-CoV-2 was infected at MOI 0.1, 1 or 2. Detection of viral RNA after 48 h post-infection. | SARS-CoV-2 isolation; infection characterization; evaluation of host responses. | [10,23] |
| Calu-3 | SARS-CoV-2/USA-WA1/2020 | Calu-3 cells were cultured in DMEM supplemented with 10 or 20% FBS at 37 °C and 5% atm CO2. Some authors have reported supplementation with antibiotics and antimycotics. SARS-CoV-2 was infected at MOI 0.01 or 0.001 and viral presence was evaluated at 24, 48, or 72 h post-infection. | Evaluation of antiviral activity. | [28] |
| A549-ACE2 | SARS-CoV-2/USA-WA1/2020 | A549 cells were maintained at 37 °C and 5% atm of CO2 and cultured in DMEM supplemented with 10% Serum Plus II. SARS-CoV-2 was infected at MOI 0.1 for 36 h. | Mapping of genes that are associated with SARS-CoV-2 infection. | [24] |
| Three-dimensional (3D) Models: Explants and Organoids | ||||
| Human nasal epithelial cells (HNE), bronchial epithelial cells (large airway respiratory cells—LAE), bronquiolar epithelial cells (small airway respiratory—SAE) | SARS-CoV-2/USA-WA1/2020 | HNE cells were grown in an air-liquid system (ALI) system. LAE and SAE cells were initially co-cultured with mitomycin-treated 3T3 J2 cells in DMEM then passed to an ALI system. After cultivation, the viruses were inoculated at MOI 0.5 and 3. Cytopathic effect not described. Viral titers were described 24 h post-infection. | SARS-CoV-2 isolation; infection characterization; evaluation of host responses. | [19,36] |
| Human alveolar epithelial cells (AECs) | BetaCoV/Hong Kong/VM20001061/2020 | Culture of primary cells in flasks. AECs were infected with SARS-CoV-2 at MOI 0.1 or 2. Detection of viral RNA after 24 h. | Infection characterization; evaluation of host responses. | [10] |
| Normal human bronchial epithelial cells (NHBE) | SARS-CoV-2/USA-WA1/2020 | ALI system was not mentioned as a cultivation mechanism. NHBE cells were cultured in bronchial epithelial growth medium supplemented with BEGM SingleQuots. SARS-CoV-2 was infected at MOI 2 for 24 h. | Infection characterization; evaluation of host responses. | [41] |
| Human tracheobronchial epithelial cells | SARS-CoV-2/USA-WA1/2020 | Cultivation of human tranqueobronchial epithelial cells was performed in an ALI system for 6 to 8 weeks. The virus was inoculated at MOI 0.5 and incubated for 48 h. | Infection characterization; evaluation of antiviral activity. | [28,42] |
| Adult human ocular cells | SARS-CoV-2/USA-WA1/2020 | Ocular tissue was maintained in DMEM/F-12 supplemented with FBS and ROCK inhibitor. SARS-CoV-2 was infected in MOI 1 for 24 h. The titrations were performed with Vero E6 cells. Real-time quantitative polymerase chain reaction (qRT-PCR) was also used for analysis. | Infection characterization; evaluation of host responses. | [55] |
| Human neural progenitor cells (hNPCs) | SARS-CoV-2 HKU-001a; SARSCoV GZ50 | hNPCs are derived from induced pluripotent stem cells (iPSCs) grown in mTeSRTM1 medium induced by a cocktail of supplements. After this induction, the cells were cultured in 1:1 DMEM/F-12 and neurobasal medium with extra supplementation. SARS-CoV-2 was infected in a MOI 10 and the infection analyzed by qRT-PCR. | Infection characterization. | [51] |
| Neurospheres | SARS-CoV-2 HKU-001a; SARSCoV GZ50 | Neurospheres are derived from iPSCs and were isolated using Accutase and then maintained in Iscove’s Modified Dulbecco’s Medium (IMDM) supplemented with 15% FBS and additional components. After the formation of the embryoid body (EB), the rosettes were formed in 96-well plates containing DMEM/F12 medium supplemented with fibroblast growth factor 2 (FGF2) and Gem21 NeuroPlex. The formation of neurospheres is given by the rotation of cells without FGF2. A virus title of 2.6 × 106 p.f.u./mL were used to infect the neurospheres. Plates and qRT-PCR assays were performed to detect and quantify SARS-CoV-2. | Infection characterization. | [51] |
| Brain organoids | SARS-CoV-2 HKU-001a; SARSCoV GZ50 | Brain organoids are derived from iPSCs (from donors or not) and were isolated using Accutase and then kept in supplemented mTeSR1 medium for three days, where a series of changes in neurobasal media takes place to form organoids. A 2.6 × 106 p.f.u./mL virus titer or supernatant of SARS-CoV-2 cultivar was used to infect the organoids. qRT-PCR plates and assays were performed to detect and quantify SARS-CoV-2. | Infection characterization. | [50,51] |
| Liver organoids | SARS-CoV-2/USA-WA1/2020 | Liver organoids were derived from human pluripotent stem cells (hPSCs), which were cultured in Matrigel medium and differentiated with Activin A, BMP-4, bFGF and hepatocyte growth factor. SARS-CoV-2 was infected with MOI 0.01, 0.05, and 0.1. The titrations were performed with Vero E6 and HEK293 cells. qRT-PCR was also used for analysis. | Infection characterization. | [47] |
| Kidney organoids | SARS-CoV-2/human/SWE/01/2020 | Kidney organoids were derived from human pluripotent stem cells (hPSCs), which were cultured in RPMI medium with multiple supplements. SARS-CoV-2 was infected at 103 or 105 viral particles and quantified by qRT-PCR. | Infection characterization. | [56] |
| Lung organoids | SARS-CoV-2/USA-WA1/2020 | Lung organoids were derived from human pluripotent stem cells (hPSCs), which were cultured in supplemented DMEM/F12 medium. SARS-CoV-2 was infected at MOI 0.01 for 24 h and quantified by qRT-PCR. | Infection characterization; evaluation of antiviral activity. | [48] |
| Colonic organoids | SARS-CoV-2/USA-WA1/2020 | Colonic organoids were derived from human pluripotent stem cells (hPSCs), which were cultured in supplemented DMEM/F12 or RPMI1640 medium. SARS-CoV-2 was infected at MOI 0.01 for 24 h and quantified by qRT-PCR. | Infection characterization; evaluation of antiviral activity. | [48] |
| Blood vessels organoids | SARS-CoV-2/human/SWE/01/2020 | Blood vessels organoids were derived from human pluripotent stem cells (hPSCs). SARS-CoV-2 was infected at 102, 104 or 106 viral particles and quantified by qRT-PCR. | Infection characterization. | [56] |
| Eye organoids | SARS-CoV-2/USA-WA1/2020 | Ocular organoids were derived from human embryonic stem cells (hESCs), which were cultured in mTeSR1 medium for 10 days and exchanged for SEAM differentiation media to form ocular organoids. SARS-CoV-2 was infected in MOI 1 for 24 h. The titrations were performed with Vero E6 cells. qRT-PCR was also used for analysis. | Infection characterization; evaluation of host responses. | [55] |
| Bronquial tissue | BetaCoV/Hong Kong/VM20001061/2020 | Bronquial tissue were maintained on F12K medium in ALI system. The authors cite extra supplementation with antibiotics and buffers. SARS-CoV-2 was infected for 1 h at 37 °C and washed with PBS buffer after that time. Supernatant samples were collected after 1, 24, 48, 72, and 96 h post-infection. The titrations were performed with Vero E6 or MDCK cells. Histological and immunohistochemical analyzes were also performed. | Infection characterization. | [10] |
| Lung tissue | BetaCoV/Hong Kong/VM20001061/2020; SARS-CoV-2/human/HKG/HKU-001a/2020 | Lung tissue fragments were maintained on supplemented DMEM/F12 medium in plaques or supplemented F12K medium in an ALI system. SARS-CoV-2 was infected for 1 or 2 h at 37 °C and then washed with PBS buffer. Supernatant samples were collected after 1, 2, 24, 48, 72, and 96 h post-infection. Titrations were performed with Vero E6 or MDCK cells. Histological and immunohistochemical analyzes were also performed. | Infection characterization. | [10,46] |
| Conjunctiva tissue | BetaCoV/Hong Kong/VM20001061/2020 | Conjunctiva tissue were maintained on supplemented F12K medium in an ALI system. SARS-CoV-2 was infected for 1 h at 33 °C and then washed with PBS buffer. Supernatant samples were collected after 1, 24, 48, 72, and 96 h post-infection. Titrations were performed with Vero E6 or MDCK cells. Histological and immunohistochemical analyzes were also performed. | Infection characterization. | [10] |
| Mouse Strain | Background | Age | Viral Strain | Route of Infection /Dose | Major Findings | Reference |
|---|---|---|---|---|---|---|
| ACE2 (angiotensin-converting enzyme) | C57BL/6 | 4–5-weeks and 30-weeks-old | BetaCoV/Wuhan/AMMS01/2020 | Intranasal/4 × 105 p.f.u./mL or Intragastric/4 × 106 p.f.u./mL | Young and elderly hACE2 (human angiotensin-converting enzyme) 2 mice showed high viral loads in the trachea and brain when inoculated intranasally. Intragastric inoculation led to pathological pulmonary changes. | [65] |
| HFH4 | C3B6 | 8–10-weeks-old | IVCAS 6.7512 | Intranasal × 104 TCID50/mL or 7 × 105 TCID50/mL | Infected mice had typical interstitial pneumonia. Viral loads were found in the lungs at higher titers, but viral RNA was also found in the eyes, heart and brain. Pre-exposure to SARS-CoV-2 has been shown to protect mice from developing severe pneumonia. | [67] |
| hACE2 | ICR | 6–11-months-old | BetaCoV/Wuhan/IVDC-HB-01/2020 | Intranasal/1 × 105 TCID50/mL | Transgenic hACE2 mice inoculated with SARS-CoV-2 showed interstitial pneumonia. Viral antigens were found in bronchial epithelial cells, alveolar macrophages and alveolar epithelium. | [68] |
| hACE2 | * | * | SARS-CoV-2 MA (clone 14569023) | Intranasal/1 × 105 p.f.u./mL | Through reverse genetics, a recombinant virus was created capable of replicating in upper and lower airways of young and elderly BALB/c mice. The disease was more evident in older animals. | [71] |
| WT (wild-type) | BALB/c | 12-months and 10-weeks-old | ||||
| K18-hACE2 | C57BL/6 | 6-weeks-old | USA-WA1/2020 | Intranasal/1 × 104 p.f.u./mL | K18-hACE2 mice allow replication of the virus at high titers in the nasal conchae, lung and brain, showing high lethality and production of cytokines and chemokines. Adenovirus-mediated delivery, on the other hand, results in viral replication with lower titers limited to the nasal conchae and lung, with no clinical signs of infection. | [74] |
| WT | BALB/c | 9-months and 6-weeks-old | BetaCov/human/CHN/Beijing_IMEBJ05/2020 | Intranasal/7.2 × 105 p.f.u./mL | The MASCp6, an adapted strain of SARS-CoV-2, infected elderly and young mice efficiently, resulting in moderate pneumonia and inflammatory response. | [77] |
| Animal Species | Age | Viral Strain | Route of Infection/Dose | Major Findings | Reference |
|---|---|---|---|---|---|
| Syrian hamster | 6–10-weeks-old | * | Intranasal/105 p.f.u./mL | Animals challenged with SARS-CoV-2 showed viral replication, severe edema, inflammation and cell death in the lungs. The animals also showed weight loss and increased respiratory rate. | [79] |
| 4–5-weeks-old | BetaCoV/Hong Kong/VM20001061/2020 | Intranasal/8 × 104 TCID50/mL | Viral antigens were observed in the nasal mucosa, bronchial epithelial cells, duodenal epithelial cells and lung of infected hamsters. Rapid viral clearance and pneumocyte hyperplasia were also found. | [80] | |
| 1-month-old and 6–7-months-old | SARS-CoV-2/UT-NCGM02/Tóquio and SARS-CoV-2/UW-001/Human/2020/Wisconsin | Intranasal and ocular /103, 105, 106 p.f.u./mL | SARS-CoV-2 isolates replicated efficiently in the animals’ lungs, causing severe lung disease. Serious lung injuries were observed. Infected hamsters developed neutralizing antibody responses that prevented infection after viral re-exposure. | [81] | |
| 6–8-weeks and 7–12-weeks-old | BetaCov/Belgium/GHB-03021/2020 | Intranasal/2 × 105 TCID50/mL or 2 × 106 TCID50/mL | Infected wild-type hamsters showed bronchopneumonia and pulmonary inflammatory response with neutrophil infiltration and edema. | [82] | |
| 6-weeks and 32–34-weeks-old | BetaCoV/Germany/BavPat1/2020 | Intranasal/1 × 105 p.f.u./mL | The replication of the virus in the upper and lower respiratory tract occurred regardless animals’ age. However, old hamsters had greater weight loss compared to young animals, in addition to developing conspicuous alveolar and perivascular edema. Viral RNA was found in the bronchial epithelium, type I and II alveolar epithelial cells and macrophages. | [83] | |
| 6–10-weeks | * | Intranasal/105 p.f.u./mL | Surgical mask partition for challenged index or naive hamsters significantly reduced the transmission to 25%. Surgical mask partition for challenged index hamsters significantly reduced transmission to only 16.7% of exposed naive hamsters. | [84] | |
| Ferrets | 3–4-months-old | F13/environnment/2020/Wuhan and CTan/human/2020Wuhan | Intranasal/1 × 105 p.f.u./mL | Viral RNA was found in nasal washes and rectal swabs from infected ferrets. Fever and appetite loss were observed in some animals. However, RNA was not detected in animals’ organs. | [95] |
| 7-months-old | Victoria/01/2020 | Intranasal/5 × 102/104/106 p.f.u./mL | High and medium viral doses induced a consistent viral infection in the animals’ upper respiratory tract, causing bronchopneumonia (high dose) and bronchointerstitial pneumonia (medium dose). | [96] | |
| 6-months-old | BetaCoV/Munich/BavPat1/2020 | Intranasal/6 × 105 TCID50/mL | SARS-CoV-2 could be transmitted via direct contact and via air (drops and/or aerosols) between ferrets. Viral RNA was detected in infected animals directly between 1 to 3 days after inoculation and after 7 days in animals infected by indirect contact. | [97] | |
| 12–20-months-old | NMC2019-nCoV02 | Intranasal/105 TCID50/mL | Infected ferrets exhibited elevated body temperatures and viral replication. The virus was retrieved from nasal samples, saliva, urine, and feces. Viral RNA was detected in the nasal concha, trachea, lungs, and intestine. The study demonstrated the possibility of transmission by direct or indirect contact. | [98] | |
| 4-months-old | USA-WA1/2020 | Intranasal/5 × 104 p.f.u./mL | Infected ferrets showed low airway immune responses when compared to Influenza A infection. | [41] | |
| Dogs | 3-months-old | CTam-H | Intranasal/1 × 105 p.f.u./mL | Infected dogs had viral RNA detected in rectal samples, but viral RNA was not detected in any other organ or tissue. The study demonstrated a low susceptibility of dogs to SARS-CoV-2. | [95] |
| 5–6-years-old | WAI/2020WY96 | Intranasal/1.4 × 105 p.f.u./mL | Dogs inoculated with SARS-CoV-2 did not develop an evident disease, but the production of neutralizing antibodies after infection was found. | [102] | |
| Cats | 70-days-old to 3-months-old | CTam-H | Intranasal/1 × 105 p.f.u./mL | The study showed that cats are susceptible to experimental infection and that virus can be transmitted to uninfected cats when housed together. The virus replicated only in the upper respiratory tract of infected cats, especially in younger animals. | [95] |
| 6–8-years-old | WAI/2020WY96 | Intranasal/3 × 105 p.f.u./mL | The findings showed that cats are highly susceptible to infection by SARS-CoV-2 by maintaining a prolonged period of oral and nasal viral release. It has been reported that infected cats develop neutralizing antibodies that prevent possible reinfection, but there are no clinical signs of the disease. The study also demonstrated the possibility of transition by direct contact between animals. | [102] |
| Animal Specie | Age | Viral Strain | Route of Infection/Dose | Major Findings | Reference |
|---|---|---|---|---|---|
| Cynomolgus macaques (Macaca fascicularis) | 4–5-years-old and 15–20-years-old | * | Intratracheal and Intranasal/* | SARS-CoV caused more severe lung injuries than SARS-CoV-2 and milder infection than MERS-CoV in these animals. | [106] |
| Rhesus macaques (Macaca mulatta) | 4–6-years-old | nCoV-WA1-2020 | Intranasal, Intratracheal and Ocular/4 × 105 TCID50/mL | Rhesus monkeys manifest the disease caused by SARS-CoV-2. The animals developed respiratory disease, and high viral loads were found in the nose, throat, and bronchoalveolar lavages. | [107] |
| Rhesus macaques (Macaca mulatta) | 3–5-years-old | WH-09/humam/2020/CHN | Ocular Conjuctiva and Intratracheal/1 × 106 TCID50/mL | The conjunctival ocular route proved to be efficient for infection of these animals, leading them to develop mild pneumonia. However, the disease did not manifest severely. | [111] |
| Rhesus macaques (Macaca mulatta), Cynomolgus macaques (Macaca fascicularis) and Marmoset (Callithrix jacchus) | * | * | Intratracheal, Intranasal and Conjuctiva/4.75 × 106 p.f.u./mL and 1 × 106 p.f.u./mL | Two families of monkeys from the Old World and one from the New World were inoculated experimentally with SARS-CoV-2. Among the studied species, M. mulatta was the most susceptible to infection followed by M. fascicularis and C. jacchus. | [108] |
| Rhesus macaques (Macaca mulatta) | 3–5-years and 15-years-old | BetaCoV/Wuhan/IVDC-HD-01/2020 | Intratracheal/1 × 106 TCID50/mL | Viral replication occurred more effectively in elderly monkeys, causing severe interstitial pneumonia. Authors suggest that elderly monkeys are useful to model the severe form of the disease. | [112] |
| Rhesus macaques (Macaca mulatta) | 3–5 years-old | * | Intratracheal/1 × 106 TCID50/mL | SARS-CoV-2 reinfection was described and its signs were presented. Authors suggest that an initial infection prepares the immune system for a possible new infection. | [113] |
| Rhesus macaques (Macaca mulatta) | 6–12-years-old | * | Intratracheal or intranasal/1.1 × 104 p.f.u./mL or 1.1 × 105 p.f.u./mL or 1.1 × 106 p.f.u./mL | SARS-CoV-2 reinfection was described and its signs were presented. Authors suggest that an initial infection prepares the immune and humoral systems for a possible new infection. | [114] |
| African green monkeys (Chlorocebus aethiops) | 3–5 years-old | SARS-CoV-2/München-1.1/2020/929 (Munich) | Intranasal, oral, ocular and intratracheal/5 × 105 p.f.u/mL | Infected young monkeys had low fever and the respiratory symptoms were limited to a transient decrease in tidal volume. Viral RNA was found in all airways and gastrointestinal system. All animals seroconverted simultaneously for IgM and IgG. | [109] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rosa, R.B.; Dantas, W.M.; do Nascimento, J.C.F.; da Silva, M.V.; de Oliveira, R.N.; Pena, L.J. In Vitro and In Vivo Models for Studying SARS-CoV-2, the Etiological Agent Responsible for COVID-19 Pandemic. Viruses 2021, 13, 379. https://doi.org/10.3390/v13030379
Rosa RB, Dantas WM, do Nascimento JCF, da Silva MV, de Oliveira RN, Pena LJ. In Vitro and In Vivo Models for Studying SARS-CoV-2, the Etiological Agent Responsible for COVID-19 Pandemic. Viruses. 2021; 13(3):379. https://doi.org/10.3390/v13030379
Chicago/Turabian StyleRosa, Rafael B., Willyenne M. Dantas, Jessica C. F. do Nascimento, Murilo V. da Silva, Ronaldo N. de Oliveira, and Lindomar J. Pena. 2021. "In Vitro and In Vivo Models for Studying SARS-CoV-2, the Etiological Agent Responsible for COVID-19 Pandemic" Viruses 13, no. 3: 379. https://doi.org/10.3390/v13030379