
Phages from Genus Bruynoghevirus and Phage Therapy: Pseudomonas Phage Delta Case

 , ,
, , 
Abstract
:1. Introduction
2. Materials and Methods
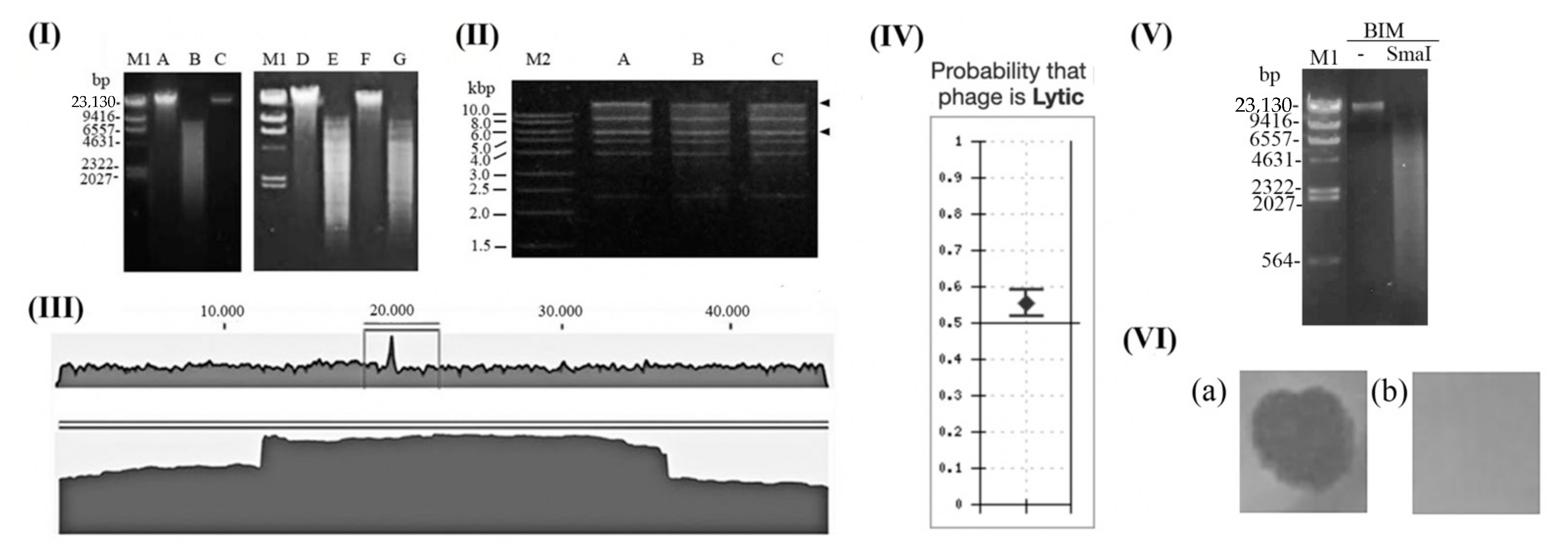
2.1. Phage Preparation
2.2. DNA Sequencing
2.3. Phage Delta Genome Analysis
2.4. Lysogeny Formation
2.5. Bacteriophage Insensitive Mutants (BIM)
2.6. Determination of Phage Genome Ends
2.7. Potential Virulence Factors
2.8. Antibiotic Resistance Genes
2.9. Potential Allergens
3. Results and Discussion
3.1. Is Bacteriophage Delta Temperate or Obligatorily Lytic Phage?
3.2. Is Pseudomonas Phage PaP3 Prone to Generalised Transduction?
3.3. Does the Phage Delta Genome Encode Undesirable Proteins?
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Knezevic, P.; Hoyle, N.S.; Matsuzaki, S.; Gorski, A. Editorial: Advances in Phage Therapy: Present Challenges and Future Perspectives. Front. Microbiol. 2021, 12, 701898. [Google Scholar] [CrossRef] [PubMed]
- Summers, W.C. The strange history of phage therapy. Bacteriophage 2012, 2, 130–133. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Z.; Kottadiel, V.I.; Vafabakhsh, R.; Dai, L.; Chemla, Y.R.; Ha, T.; Rao, V.B. A Promiscuous DNA Packaging Machine from Bacteriophage T4. PLoS Biol. 2011, 9, e1000592. [Google Scholar] [CrossRef] [PubMed]
- Gibson, E.J. Chapter 23. In An Odyssey in Learning and Perception; The MIT Press: Cambridge, MA, USA, 2020; p. 501. [Google Scholar] [CrossRef]
- Chen, J.; Novick, R.P. Phage-Mediated Intergeneric Transfer of Toxin Genes. Science 2009, 323, 139–141. [Google Scholar] [CrossRef] [Green Version]
- Balcazar, J.L. Bacteriophages as Vehicles for Antibiotic Resistance Genes in the Environment. PLOS Pathog. 2014, 10, e1004219. [Google Scholar] [CrossRef] [Green Version]
- James, C.; Davies, E.V.; Fothergill, J.; Walshaw, M.J.; Beale, C.M.; Brockhurst, M.A.; Winstanley, C. Lytic activity by temperate phages of Pseudomonas aeruginosa in long-term cystic fibrosis chronic lung infections. ISME J. 2015, 9, 1391–1398. [Google Scholar] [CrossRef] [Green Version]
- Shousha, A.; Awaiwanont, N.; Sofka, D.; Smulders, F.J.M.; Paulsen, P.; Szostak, M.P.; Humphrey, T.; Hilbert, F. Bacteriophages Isolated from Chicken Meat and the Horizontal Transfer of Antimicrobial Resistance Genes. Appl. Environ. Microbiol. 2015, 81, 4600–4606. [Google Scholar] [CrossRef] [Green Version]
- Waldor, M.K.; Mekalanos, J.J. Lysogenic Conversion by a Filamentous Phage Encoding Cholera Toxin. Science 1996, 272, 1910–1914. [Google Scholar] [CrossRef] [Green Version]
- Tan, Y.; Zhang, K.; Rao, X.; Jin, X.; Huang, J.; Zhu, J.; Chen, Z.; Hu, X.; Shen, X.; Wang, L.; et al. Whole genome sequencing of a novel temperate bacteriophage ofP. aeruginosa: Evidence of tRNA gene mediating integration of the phage genome into the host bacterial chromosome. Cell. Microbiol. 2006, 9, 479–491. [Google Scholar] [CrossRef]
- Zschach, H.; Joensen, K.G.; Lindhard, B.; Lund, O.; Goderdzishvili, M.; Chkonia, I.; Jgenti, G.; Kvatadze, N.; Alavidze, Z.; Kutter, E.M.; et al. What Can We Learn from a Metagenomic Analysis of a Georgian Bacteriophage Cocktail? Viruses 2015, 7, 6570–6589. [Google Scholar] [CrossRef] [Green Version]
- Ceyssens, P.-J.; Hertveldt, K.; Ackermann, H.-W.; Noben, J.-P.; Demeke, M.; Volckaert, G.; Lavigne, R. The intron-containing genome of the lytic Pseudomonas phage LUZ24 resembles the temperate phage PaP3. Virology 2008, 377, 233–238. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Knezevic, P.; Kostanjsek, R.; Obreht, D.; Petrovic, O. Isolation of Pseudomonas aeruginosa Specific Phages with Broad Activity Spectra. Curr. Microbiol. 2009, 59, 173–180. [Google Scholar] [CrossRef] [PubMed]
- Knezevic, P.; Obreht, D.; Curcin, S.; Petrusic, M.; Aleksic, V.; Kostanjšek, R.; Petrović, O. Phages of Pseudomonas aeruginosa: Response to environmental factors and in vitro ability to inhibit bacterial growth and biofilm formation. J. Appl. Microbiol. 2011, 111, 245–254. [Google Scholar] [CrossRef]
- Sambrook, J.; Russell, D. Molecular Cloning: A Laboratory Manual, 3rd ed.; Cold Spring Harbor Laboratory Press: Cold Spring Harbor, NY, USA, 2001. [Google Scholar]
- Besemer, J.; Lomsadze, A.; Borodovsky, M. GeneMarkS: A self-training method for prediction of gene starts in microbial genomes. Implications for finding sequence motifs in regulatory regions. Nucleic Acids Res. 2001, 29, 2607–2618. [Google Scholar] [CrossRef] [Green Version]
- Aziz, R.K.; Bartels, D.; Best, A.A.; DeJongh, M.; Disz, T.; Edwards, R.A.; Formsma, K.; Gerdes, S.; Glass, E.M.; Kubal, M.; et al. The RAST Server: Rapid Annotations using Subsystems Technology. BMC Genom. 2008, 9, 75. [Google Scholar] [CrossRef] [Green Version]
- Adriaenssens, E.; Brister, J.R. How to Name and Classify Your Phage: An Informal Guide. Viruses 2017, 9, 70. [Google Scholar] [CrossRef] [Green Version]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular Evolutionary Genetics Analysis across Computing Platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef]
- Felsenstein, J. Confidence limits on phylogenies: An approach using the bootstrap. Evolution 1985, 39, 783–791. [Google Scholar] [CrossRef]
- Laslett, D.; Canback, B. ARAGORN, a program to detect tRNA genes and tmRNA genes in nucleotide sequences. Nucleic Acids Res. 2004, 32, 11–16. [Google Scholar] [CrossRef]
- Lowe, T.M.; Chan, P.P. tRNAscan-SE On-line: Integrating search and context for analysis of transfer RNA genes. Nucleic Acids Res. 2016, 44, W54–W57. [Google Scholar] [CrossRef]
- McNair, K.; Bailey, B.A.; Edwards, R.A. PHACTS, a computational approach to classifying the lifestyle of phages. Bioinformatics 2012, 28, 614–618. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- O’Flynn, G.; Coffey, A.; Fitzgerald, G.; Ross, R. The newly isolated lytic bacteriophages st104a and st104b are highly virulent against Salmonella enterica. J. Appl. Microbiol. 2006, 101, 251–259. [Google Scholar] [CrossRef]
- Casjens, S.R.; Gilcrease, E.B. Determining DNA Packaging Strategy by Analysis of the Termini of the Chromosomes in Tailed-Bacteriophage Virions. Stem Cells and Aging 2009, 502, 91–111. [Google Scholar] [CrossRef] [Green Version]
- Chen, L.; Zheng, D.; Liu, B.; Yang, J.; Jin, Q. VFDB 2016: Hierarchical and refined dataset for big data analysis—10 years on. Nucleic Acids Res. 2016, 44, D694–D697. [Google Scholar] [CrossRef] [PubMed]
- Alcock, B.P.; Raphenya, A.R.; Lau, T.T.Y.; Tsang, K.K.; Bouchard, M.; Edalatmand, A.; Huynh, W.; Nguyen, A.-L.V.; Cheng, A.A.; Liu, S.; et al. CARD 2020: Antibiotic resistome surveillance with the comprehensive antibiotic resistance database. Nucleic Acids Res. 2020, 48, D517–D525. [Google Scholar] [CrossRef] [PubMed]
- Aalberse, R.C. Structural biology of allergens. J. Allergy Clin. Immunol. 2000, 106, 228–238. [Google Scholar] [CrossRef] [PubMed]
- Codex Alimentarius Commission. Alinorm 03/34: Joint FAO/WHO Food Standard Programme, Codex Alimentarius Commission, Twenty-Fifth Session, Rome, Italy, 30 June–5 July 2003. Appendix III, Guideline for the Conduct of Food Safety Assessment of Foods Derived from-DNA Plants and Appendix IV, Annex on the Assessment of Possible Allergenicity. 2003, pp. 47–60. Available online: http://www.fao.org/3/y4800e/y4800e00.htm (accessed on 16 July 2016).
- Colloms, S.D.; Sykora, P.; Szatmari, G.; Sherratt, D.J. Recombination at ColE1 cer requires the Escherichia coli xerC gene product, a member of the lambda integrase family of site-specific recombinases. J. Bacteriol. 1990, 172, 6973–6980. [Google Scholar] [CrossRef] [Green Version]
- Alemayehu, D.; Casey, P.G.; Mcauliffe, O. Bacteriophages ϕMR299-2 and ϕNH-4 Can Eliminate Pseudomonas aeruginosa in the Murine Lung and on Cystic Fibrosis Lung Airway Cells. MBio 2012, 3, e00029-12. [Google Scholar] [CrossRef] [Green Version]
- Olsen, N.S.; Hendriksen, N.B.; Hansen, L.H.; Kot, W. A New High-Throughput Screening Method for Phages: Enabling Crude Isolation and Fast Identification of Diverse Phages with Therapeutic Potential. Phage 2020, 1, 137–148. [Google Scholar] [CrossRef]
- Patpatia, S.; Yilmaz, O.; Ylänne, M.; Kiljunen, S. Isolation and Genomic Analysis of the Phage vB_PaeP_fHoPae04 Infecting Pseudomonas aeruginosa. Microbiol. Resour. Announc. 2021, 10, e0007621. [Google Scholar] [CrossRef]
- Magill, D.J.; Shaburova, O.V.; Chesnokova, E.N.; Pleteneva, E.A.; Krylov, V.N.; Kulakov, L.A. Complete nucleotide sequence of phiCHU: A Luz24likevirus infecting Pseudomonas aeruginosa and displaying a unique host range. FEMS Microbiol. Lett. 2015, 362, 2014–2016. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fong, S.A.; Drilling, A.; Morales, S.; Cornet, M.E.; Woodworth, B.A.; Fokkens, W.J.; Psaltis, A.; Vreugde, S.; Wormald, P.-J. Activity of Bacteriophages in Removing Biofilms of Pseudomonas aeruginosa Isolates from Chronic Rhinosinusitis Patients. Front. Cell. Infect. Microbiol. 2017, 7, 418. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alves, D.R.; Esteban, P.P.; Kot, W.; Bean, J.; Arnot, T.; Hansen, L.; Enright, M.; Jenkins, T. A novel bacteriophage cocktail reduces and dispersesPseudomonas aeruginosabiofilms under static and flow conditions. Microb. Biotechnol. 2015, 9, 61–74. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pires, D.P.; Kropinski, A.M.; Azeredo, J.; Sillankorva, S. Complete genome sequence of the Pseudomonas aeruginosa bacteriophage phiIBB-PAA2. Genome Announc. 2014, 2, 7–8. [Google Scholar] [CrossRef] [Green Version]
- Zhang, L.; Le, S.; Lu, S.-G.; Yao, X.-Y.; Zhao, Y.; Wang, J.; Tan, Y.-L.; Hu, F.-Q.; Li, M. Isolation and identification a novel Pseudomonas aeruginosa phage PaP4. Microbiol. China 2013, 40, 609–616. [Google Scholar]
- Alvi, I.A.; Asif, M.; Rehman, S.U. A single dose of a virulent bacteriophage vB PaeP-SaPL, rescues bacteremic mice infected with multi drug resistant Pseudomonas aeruginosa. Virus Res. 2021, 292, 198250. [Google Scholar] [CrossRef] [PubMed]
- Campbell, R.A.; Farlow, J.; Freyberger, H.R.; He, Y.; Ward, A.M.; Ellison, D.W.; Getnet, D.; Swierczewski, B.E.; Nikolich, M.P.; Filippov, A.A. Genome Sequences of 17 Diverse Pseudomonas aeruginosa Phages. Microbiol. Resour. Announc. 2021, 10, e00031-21. [Google Scholar] [CrossRef]
- Farlow, J.; Freyberger, H.R.; He, Y.; Ward, A.M.; Rutvisuttinunt, W.; Li, T.; Campbell, R.; Jacobs, A.C.; Nikolich, M.P.; Filippov, A.A. Complete Genome Sequences of 10 Phages Lytic against Multidrug-Resistant Pseudomonas aeruginosa. Microbiol. Resour. Announc. 2020, 9, e00503-20. [Google Scholar] [CrossRef]
- Pleteneva, E.A.; Bourkaltseva, M.V.; Shaburova, O.V.; Krylov, S.V.; Pechnikova, E.V.; Sokolova, O.; Krylov, V.N. TL, the new bacteriophage of Pseudomonas aeruginosa and its application for the search of halo-producing bacteriophages. Russ. J. Genet. 2011, 47, 5–9. [Google Scholar] [CrossRef]
- Latz, S.; Krüttgen, A.; Häfner, H.; Buhl, E.M.; Ritter, K.; Horz, H.-P. Differential Effect of Newly Isolated Phages Belonging to PB1-Like, phiKZ-Like and LUZ24-Like Viruses against Multi-Drug Resistant Pseudomonas aeruginosa under Varying Growth Conditions. Viruses 2017, 9, 315. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Glukhov, A.S.; Krutilina, A.I.; Shlyapnikov, M.G.; Severinov, K.; Lavysh, D.; Kochetkov, V.V.; McGrath, J.W.; De Leeuwe, C.; Shaburova, O.V.; Krylov, V.N.; et al. Genomic Analysis of Pseudomonas putida Phage tf with Localized Single-Strand DNA Interruptions. PLoS ONE 2012, 7, e51163. [Google Scholar] [CrossRef] [Green Version]
- Storey, N.; Rabiey, M.; Neuman, B.W.; Jackson, R.W.; Mulley, G. Genomic Characterisation of Mushroom Pathogenic Pseudomonads and Their Interaction with Bacteriophages. Viruses 2020, 12, 1286. [Google Scholar] [CrossRef]
- Eller, M.R.; Salgado, R.L.; Vidigal, P.M.; Alves, M.P.; Dias, R.S.; de Oliveira, L.L.; da Silva, C.C.; de Carvalho, A.F.; De Paula, S.O. Complete Genome Sequence of the Pseudomonas fluorescens Bacteriophage UFV-P2. Genome Announc. 2013, 1, e00006-12. [Google Scholar] [CrossRef] [Green Version]
- Reiter, W.D.; Palm, P.; Yeats, S. Transfer RNA genes frequently serve as integration sites for prokaryotic genetic elements. Nucleic Acids Res. 1989, 17, 5. [Google Scholar] [CrossRef] [Green Version]
- Lei, X.; Wang, L.; Zhao, G.; Ding, X. Site-specificity of serine integrase demonstrated by the attB sequence preference of ɸ BT 1 integrase. FEBS Lett. 2018, 592, 1389–1399. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mohaisen, M.R.; McCarthy, A.J.; Adriaenssens, E.M.; Allison, H.E. The Site-Specific Recombination System of the Escherichia coli Bacteriophage Φ24B. Front. Microbiol. 2020, 11, 578056. [Google Scholar] [CrossRef] [PubMed]
- Campbell, A.M. Chromosomal insertion sites for phages and plasmids. J. Bacteriol. 1992, 174, 7495–7499. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bailly-Bechet, M.; Vergassola, M.; Rocha, E. Causes for the intriguing presence of tRNAs in phages. Genome Res. 2007, 17, 1486–1495. [Google Scholar] [CrossRef] [Green Version]
- Weiss, S.B.; Hsu, W.T.; Foft, J.W.; Scherberg, N.H. Transfer RNA coded by the T4 bacteriophage genome. Proc. Natl. Acad. Sci. USA 1968, 61, 114–121. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kihara, A.; Akiyama, Y.; Ito, K. Host regulation of lysogenic decision in bacteriophage: Transmembrane modulation of FtsH (HflB), the cII degrading protease, by HflKC (HflA). Proc. Natl. Acad. Sci. USA 1997, 94, 5544–5549. [Google Scholar] [CrossRef] [Green Version]
- Rokney, A.; Kobiler, O.; Amir, A.; Court, D.L.; Stavans, J.; Adhya, S.; Oppenheim, A.B. Host responses influence on the induction of lambda prophage. Mol. Microbiol. 2008, 68, 29–36. [Google Scholar] [CrossRef] [Green Version]
- Morello, E.; Saussereau, E.; Maura, D.; Huerre, M.; Touqui, L.; Debarbieux, L. Pulmonary Bacteriophage Therapy on Pseudomonas aeruginosa Cystic Fibrosis Strains: First Steps Towards Treatment and Prevention. PLoS ONE 2011, 6, e16963. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Uchiyama, J.; Rashel, M.; Takemura, I.; Kato, S.-I.; Ujihara, T.; Muraoka, A.; Matsuzaki, S.; Daibata, M. Genetic characterization of Pseudomonas aeruginosa bacteriophage KPP10. Arch. Virol. 2012, 157, 733–738. [Google Scholar] [CrossRef] [PubMed]
- Henry, M.; Lavigne, R.; Debarbieux, L. Predicting In Vivo Efficacy of Therapeutic Bacteriophages Used To Treat Pulmonary Infections. Antimicrob. Agents Chemother. 2013, 57, 5961–5968. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Essoh, C.; Latino, L.; Midoux, C.; Blouin, Y.; Loukou, G.; Nguetta, S.-P.A.; Lathro, S.; Cablanmian, A.; Kouassi, A.K.; Vergnaud, G.; et al. Investigation of a Large Collection of Pseudomonas aeruginosa Bacteriophages Collected from a Single Environmental Source in Abidjan, Côte d’Ivoire. PLoS ONE 2015, 10, e0130548. [Google Scholar] [CrossRef] [Green Version]
- Coleman, S.; Yao, T.; Nguyen, T.V.P.; Golding, I.; Igoshin, O. Bacteriophage self-counting in the presence of viral replication. bioRxiv 2021. [Google Scholar] [CrossRef]
- Zhao, X.; Chen, C.; Shen, W.; Huang, G.; Le, S.; Lu, S.; Li, M.; Zhao, Y.; Wang, J.; Rao, X.; et al. Global Transcriptomic Analysis of Interactions between Pseudomonas aeruginosa and Bacteriophage PaP3. Sci. Rep. 2016, 6, 19237. [Google Scholar] [CrossRef] [Green Version]
- Chen, J.; Quiles-Puchalt, N.; Chiang, Y.N.; Bacigalupe, R.; Fillol-Salom, A.; Chee, M.S.J.; Fitzgerald, J.R.; Penadés, J.R. Genome hypermobility by lateral transduction. Science 2018, 362, 207–212. [Google Scholar] [CrossRef] [Green Version]
- Garneau, J.R.; Depardieu, F.; Fortier, L.-C.; Bikard, D.; Monot, M. PhageTerm: A tool for fast and accurate determination of phage termini and packaging mechanism using next-generation sequencing data. Sci. Rep. 2017, 7, 8292. [Google Scholar] [CrossRef]
- Casjens, S. Prophages and bacterial genomics: What have we learned so far? Mol. Microbiol. 2003, 49, 277–300. [Google Scholar] [CrossRef]
- Eller, M.R.; Vidigal, P.M.P.; Salgado, R.L.; Alves, M.P.; Dias, R.S.; da Silva, C.C.; de Carvalho, A.F.; Kropinski, A.; O De Paula, S. UFV-P2 as a member of the Luz24likevirus genus: A new overview on comparative functional genome analyses of the LUZ24-like phages. BMC Genom. 2014, 15, 7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Savalia, D.; Westblade, L.F.; Goel, M.; Florens, L.; Kemp, P.; Akulenko, N.; Pavlova, O.; Padovan, J.C.; Chait, B.T.; Washburn, M.; et al. Genomic and Proteomic Analysis of phiEco32, a Novel Escherichia coli Bacteriophage. J. Mol. Biol. 2008, 377, 774–789. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schneider, C.L. Bacteriophage-Mediated Horizontal Gene Transfer: Transduction. In Bacteriophages; Springer: Cham, Switzerland, 2017; pp. 1–42. [Google Scholar] [CrossRef]
- Guttman, B.; Raya, R.; Kutter, E. Basic phage biology. In Bacteriophages; CRC Press: Boca Raton, FL, USA, 2005. [Google Scholar]
- FAO; WHO. Evaluation of Allergenicity of Genetically Modified Foods Derived from Biotechnology; FAO: Rome, Italy, 2001. [Google Scholar]
- Curtright, A.J.; Abedon, S.T. Phage Therapy: Emergent Property Pharmacology. J. Bioanal. Biomed. 2012, S6, 2. [Google Scholar] [CrossRef] [Green Version]
- Petrovic Fabijan, A.; Lin, R.C.Y.; Ho, J.; Maddocks, S.; Ben Zakour, N.L.; Iredell, J.R.; Westmead Bacteriophage Therapy Team. Safety of bacteriophage therapy in severe Staphylococcus aureus infection. Nat. Microbiol. 2020, 5, 465–472. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Genus in Family Podoviridae | Species | Strain 1 Access. No. | Genome (bp)/No. CDS | Origin (Country) | Plaques (mm) | Head Diameter (nm) | tRNA | Similarity to Delta (%) | HflK/C/cos 1 | Integrat. Tolaasii/PaP3 | Phage Lifestyle 2 | Ref. |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Bruynoghevirus | Pseudomonas virus PaP3 | PaP3 AY078382 | 45,503/72 | Hospital sewage/China | Turbid (1.5) | 55 | Asn, Asp, Pro, Tyr | 96.2 | N/Y | N/Y | 0.51 ± 0.04 L | [10] |
| C1-14_Or HE983844 | 45,469/64 | Sewage water/France | N. A. 3 | 60 | Asn, Asp, Pro | 96.3 | N/Y | N/Y | 0.51 ± 0.03 L | [30] | ||
| P2-10_Or1 HF543949 | 44,030/71 | Eliava “Pyophage”/Georgia | N. A. | 58–60 | Asn, Asp, Pro | 96.8 | N/Y | N/Y | 0.52 ± 0.05 L | [30] | ||
| MR299-2 JN254801 | 44,789/68 | Sewage from water treatment plant | N. A. | 40–60 | Asn, Asp, Pro | 96.6 | N/Y | N/Y | 0.53 ± 0.03 L * | [31] | ||
| otherone MT119373.1 | 44,930/67 | Wastewater | N.A. | N.A. | Asn, Asp, Pro | 96.9 | N/Y | N/Y | 0.53 ± 0.04 L | [32] | ||
| Clash MT119362.1 | 44,912/67 | Wastewater | N.A. | N.A. | Asn, Asp, Pro | 96.8 | N/Y | N/Y | 0.53 ± 0.06 L | [32] | ||
| Delta MG432151.1 | 45,970/69 | Municipal wastewater/Serbia | Clear/turbid (2.5–5.0) | 63 | Asn, Asp, Pro | 100.0 | N/Y | N/Y | 0.54 ± 0.03 L * | This ref. | ||
| vB_PaeP_fHoPae04 MW329986.1 | 45,491/70 | Hospital wastewater, Finland | N.A. | N.A. | Asn, Asp, Pro | 96.4 | N/Y | N/Y | 0.53 ± 0.03 L | [33] | ||
| Epa 1 MT108723.1 | 45,230/67 | N.A. | N.A. | N.A. | Asn, Asp, Pro | 94.3 | N/Y | N/Y | 0.52 ± 0.04 L | N.A. | ||
| Pseudomonas virus CHU | CHU KP233880.1 | 45,626/76 | Pond/Russia | Variable | N.A. | Pro, Asp, Asn | 92.1 | N/Y | N/Y | 0.54 ± 0.06 L | [34] | |
| Pseudomonas virus Pa223 | Pa223 MK837012.1 | 45,703/71 | N.A. | N.A. | N.A. | Asn *, Tyr *, Pro * | 78.4 | Y/Y | N/Y (1) * | 0.58 ± 0.05 L * | [35] | |
| Pseudomonas virus Luz24 | Luz24 AM910650.1 | 45,503/68 | Hospital sewage/Belgium | Clear/turbid (1.0–5.0) | 63 | Asn *, Tyr *, Pro * | 74.3 | N/N | N/Y (1) | 0.66 ± 0.12 L * | [12] | |
| Pseudomonas virus Dl54 | DL54 KR054029.1 | 45,673/71 | crude sewage or flood water/UK | Clear/turbid | 45 | Ile *, Asp *, Pro * | 77.1 | Y/Y | N/Y (1) | 0.53 ± 0.07 L | [36] | |
| Pseudomonas virus C2-10_Ab22 | C2-10_Ab22 LN610578.1 | 45,808/71 | Carrefour de l’Indénié/Ivory Coast | N.A. | N.A. | Pro, Tyr, Asn | 74.8 | Y/N | N/Y (1) | 0.55 ± 0.08 L | [30] | |
| Pseudomonas virus phiBB-PAA2 | phiBB-PAA2 KF856712.1 | 45,344/66 | Hospital sewage/Portugal | N.A. | N.A. | Pro, Asp, Ile | 81.8 | N/Y | N/Y (1) | 0.58 ± 0.06 L * | [37] | |
| Pseudomonas virus Pap4 | Pap4 KC294142.1 | 43,895 */70 | N.A. | Transparent (3.0–5.0) | 50 | No | 75.3 | Y/Y | N/N | 0.58 ± 0.05 L * | [38] | |
| phiPAO1-EW MG589386.1 | 46,403/71 | N.A. | N.A. | N.A. | Ile, asp, Pro | 75.5 | Y/Y | N/Y (3) | 0.56 ± 0.05 L * | N.A. | ||
| phiPAO1_302 MG589385.1 | 46,093/70 | N.A. | N.A. | N.A. | Pro, Tyr, Asp, Ile | 65.3 | Y/Y | N/Y (1) | 0.54 ± 0.09 L | N.A. | ||
| SaPL MH973725.1 | 45,796/63 | Samanabad Canal, Lahore/Pakistan | Transparent (4.0–5.0) | N.A. | Asn *, Asp *, Tyr *, Pro * | 64.5 | Y/Y | N/Y (1) | 0.56 ± 0.07 L | [39] | ||
| EPa4 MT118288.1 | 45,439/53 | N.A: | N.A. | N.A. | Asn *, Tyr *, Pro * | 64.9 | Y/Y | N/Y (1) | 0.56 ± 0.05 L * | [40] | ||
| Pa222 MK837011.1 | 45,770/58 | N.A. | N.A. | N.A. | Asn *, Asp *, Tyr *, Pro * | 78.4 | Y/Y | N/Y (1) | 0.56 ± 0.06 L * | [35] | ||
| Oldone MT119371.1 | 45,313/70 | Wastewater | N.A. | N.A. | Asn *, Asp *, Tyr *, Pro * | 78.8 | Y/Y | N/Y (1) | 0.52 ± 0.07 L | [32] | ||
| U47 MN562749.1 | 43,444/68 | N.A. | N.A. | N.A. | Asn *, Asp *, Tyr *, Pro* | 78.2 | Y/Y | N/Y (1) | 0.59 ± 0.07 L * | N.A. | ||
| Epa 2 MT108724.1 | 43,229/51 | N.A. | N.A. | N.A. | Asn *, Tyr *, Pro * | 78.0 | N/Y | N/Y (1) | 0.53 ± 0.06 L | [41] | ||
| TL HG518155.1 | 45,696/65 | N.A. | Transparent turbid, large | Asn *, Tyr *, Pro * | 73.3 | Y/N | N/Y (1) | 0.55 ± 0.08 L | [42] | |||
| SL4 MF768469.1 | 44,194/65 | Hospital sewage, Germany | N.A. | 55 | Asn *, Tyr *, Pro * | 76.4 | Y/N | N/Y (1) | 0.54 ± 0.05 L | [43] | ||
| Krylovvirus | Pseudomonas virus tf | tf HE611333 | 46,271/72 | N.A. | N.A. | N.A. | Tyr * | 23.1 | N/N | N/N | 0.52 ± 0.04 L | [44] |
| Pseudomonas virus SCZY1 | SCYZ1 MH518298.3 | 47,475/62 | Soil | N.A. | N.A. | Pro * | 2.3 | Y/N | N/N | 0.51 ± 0.04 L | N.A. | |
| Vicosavirus | Pseudomonas virus NV1 | NV1 NC_042107.1 | 45,058/64 | Untreated sewage, River Thames, UK | Hazy(<2) | No | 35.0 | Y/N | Y (1)/N | 0.52 ± 0.05 T | [45] | |
| Pseudomonas virus UVF-P2 | UVF-P2 JX863101 | 45,517/75 | Dairy industry wastewater, Brazil | N.A. | N.A. | No | 34.7 | Y/N | Y (2)/N | 0.53 ± 0.03 T * | [46] | |
| Prophage CP020369.1 (6815074.. 6769117) | 52.973/85 | - | - | - | No | Y/N | Y/N | 0.50 ± 0.04 T | This ref. | |||
| Bjornvirus | Pseudomonas virus Bjorn | Bjorn NC_042103.1 | 45,936/69 | Plant compost | N.A. | N.A. | No | 20.0 | N/N | N/N | 0.53 ± 0.05 L | N.A. |
| BIM Frequency | ||
|---|---|---|
| Bacterial Host | after 24 h | after 48 h |
| PA-4U | 3.30 × 10−7 ± 3.12 × 10−7 | 5.38 × 10−7 ± 5.56 × 10−8 |
| UB-2596 | 2.36 × 10−6 ± 2.74 × 10−7 | 5.67 × 10−6 ± 1.67 × 10−6 |
| OB-7025 | 7.95 × 10−4 ± 3.24 × 10−5 | 9.62 × 10−4 ± 4.12 × 10−5 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Knezevic, P.; Petrovic Fabijan, A.; Gavric, D.; Pejic, J.; Doffkay, Z.; Rakhely, G. Phages from Genus Bruynoghevirus and Phage Therapy: Pseudomonas Phage Delta Case. Viruses 2021, 13, 1965. https://doi.org/10.3390/v13101965
Knezevic P, Petrovic Fabijan A, Gavric D, Pejic J, Doffkay Z, Rakhely G. Phages from Genus Bruynoghevirus and Phage Therapy: Pseudomonas Phage Delta Case. Viruses. 2021; 13(10):1965. https://doi.org/10.3390/v13101965
Chicago/Turabian StyleKnezevic, Petar, Aleksandra Petrovic Fabijan, Damir Gavric, Jovana Pejic, Zsolt Doffkay, and Gábor Rakhely. 2021. "Phages from Genus Bruynoghevirus and Phage Therapy: Pseudomonas Phage Delta Case" Viruses 13, no. 10: 1965. https://doi.org/10.3390/v13101965