
Roles of Virion-Incorporated CD162 (PSGL-1), CD43, and CD44 in HIV-1 Infection of T Cells


{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Mechanism(s) That Promote the Virion Incorporation of PSGL-1, CD43, and CD44
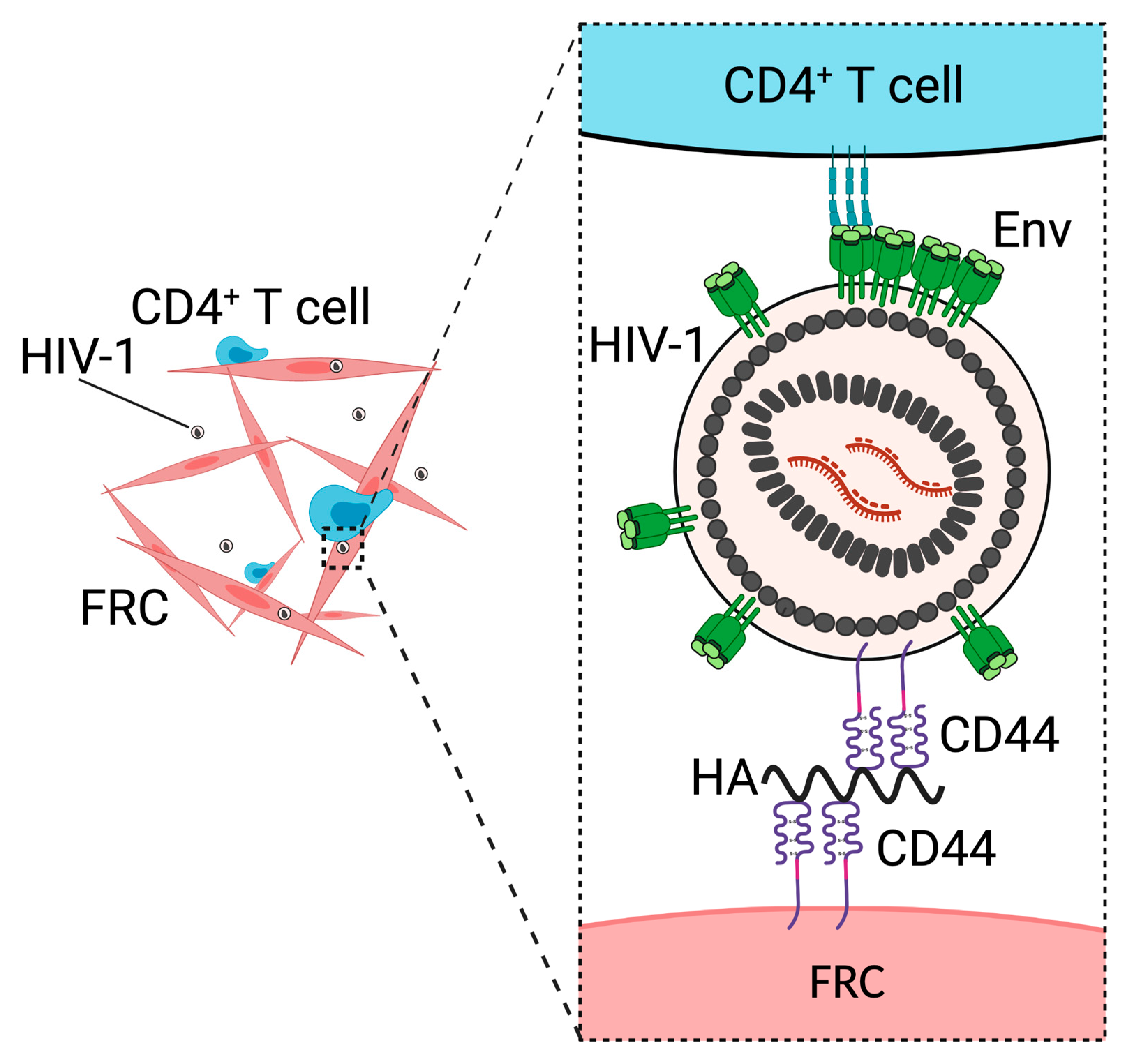
3. CD44 Promotes Fibroblastic Reticular Cell-Mediated Trans-Infection
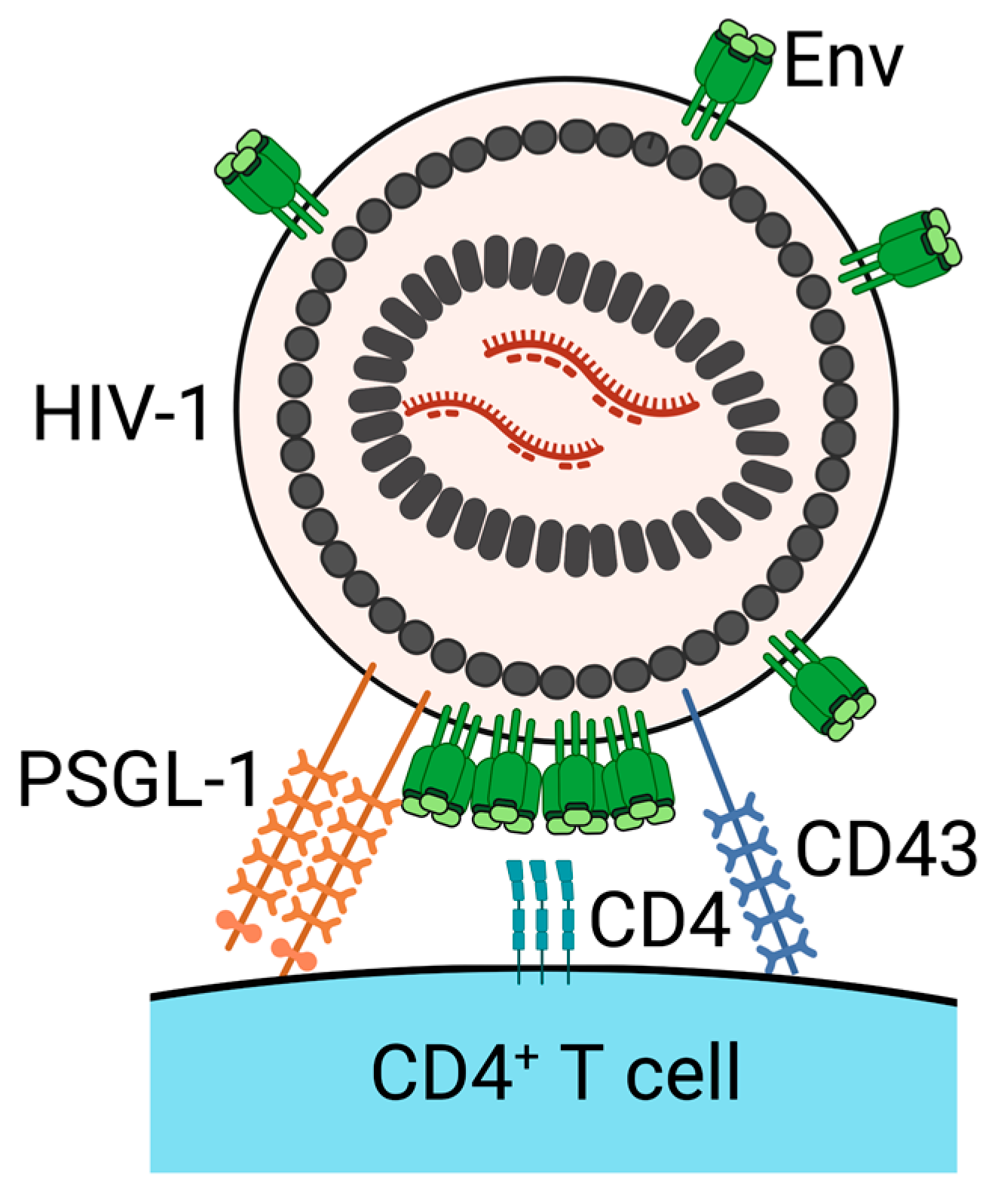
4. PSGL-1 and CD43 Inhibit Virus Attachment to Target Cells
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Burnie, J.; Guzzo, C. The incorporation of host proteins into the external HIV-1 envelope. Viruses 2019, 11, 85. [Google Scholar] [CrossRef] [Green Version]
- Tardif, M.R.; Tremblay, M.J. Presence of Host ICAM-1 in human immunodeficiency virus type 1 virions increases productive infection of CD4 + T lymphocytes by favoring cytosolic delivery of viral material. J. Virol. 2003, 77, 12299–12309. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kondo, N.; Melikyan, G.B. Intercellular adhesion molecule 1 promotes HIV-1 attachment but not fusion to target cells. PLoS ONE 2012, 7, e44827. [Google Scholar] [CrossRef] [PubMed]
- Yu, X.; Shang, H.; Jiang, Y. ICAM-1 in HIV infection and underlying mechanisms. Cytokine 2019, 125, 154830. [Google Scholar] [CrossRef]
- Guzzo, C.; Ichikawa, D.; Park, C.; Phillips, D.; Liu, Q.; Zhang, P.; Kwon, A.; Miao, H.; Lu, J.; Rehm, C.; et al. Virion incorporation of integrin alpha4beta7 facilitates HIV-1 infection and intestinal homing. Sci. Immunol. 2017, 2, eaam7341. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Q.; Lusso, P. Integrin alpha4beta7 in HIV-1 infection: A critical review. J. Leukoc. Biol. 2020, 108, 627–632. [Google Scholar] [CrossRef] [PubMed]
- Rosa, A.; Chande, A.; Ziglio, S.; De Sanctis, V.; Bertorelli, R.; Goh, S.L.; McCauley, S.M.; Nowosielska, A.; Antonarakis, S.; Luban, J.; et al. HIV-1 Nef promotes infection by excluding SERINC5 from virion incorporation. Nature 2015, 526, 212–217. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Usami, Y.; Wu, Y.; Göttlinger, H.G. SERINC3 and SERINC5 restrict HIV-1 infectivity and are counteracted by Nef. Nature 2015, 526, 218–223. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Firrito, C.; Bertelli, C.; Vanzo, T.; Chande, A.; Pizzato, M. SERINC5 as a New Restriction factor for human immunodeficiency virus and murine leukemia virus. Annu. Rev. Virol. 2018, 5, 323–340. [Google Scholar] [CrossRef] [Green Version]
- Murakami, T.; Ono, A. HIV-1 entry: Duels between Env and host antiviral transmembrane proteins on the surface of virus particles. Curr. Opin. Virol. 2021, 50, 59–68. [Google Scholar] [CrossRef]
- Compton, A.A.; Bruel, T.; Porrot, F.; Mallet, A.; Sachse, M.; Euvrard, M.; Liang, C.; Casartelli, N.; Schwartz, O. IFITM Proteins Incorporated into HIV-1 Virions Impair Viral Fusion and Spread. Cell Host Microbe 2014, 16, 736–747. [Google Scholar] [CrossRef] [Green Version]
- Tartour, K.; Appourchaux, R.; Gaillard, J.; Nguyen, X.-N.; Durand, S.; Turpin, J.; Beaumont, E.; Roch, E.; Berger, G.; Mahieux, R.; et al. IFITM proteins are incorporated onto HIV-1 virion particles and negatively imprint their infectivity. Retrovirology 2014, 11, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Shi, G.; Schwartz, O.; Compton, A.A. More than meets the I: The diverse antiviral and cellular functions of interferon-induced transmembrane proteins. Retrovirology 2017, 14, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Marziali, F.; Cimarelli, A. Membrane interference against HIV-1 by Intrinsic antiviral factors: The case of IFITMs. Cells 2021, 10, 1171. [Google Scholar] [CrossRef]
- Chen, P.; Hübner, W.; Spinelli, M.; Chen, B.K. Predominant mode of human immunodeficiency virus transfer between T cells is mediated by sustained env-dependent neutralization-resistant virological synapses. J. Virol. 2007, 81, 12582–12595. [Google Scholar] [CrossRef] [Green Version]
- Llewellyn, G.N.; Hogue, I.B.; Grover, J.R.; Ono, A. Nucleocapsid Promotes Localization of HIV-1 gag to uropods that participate in virological synapses between T Cells. PLoS Pathog. 2010, 6, e1001167. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Llewellyn, G.N.; Grover, J.R.; Olety, B.; Ono, A. HIV-1 Gag Associates with specific uropod-directed microdomains in a manner dependent on its MA highly basic region. J. Virol. 2013, 87, 6441–6454. [Google Scholar] [CrossRef] [Green Version]
- Martin-Jaular, L.; Nevo, N.; Schessner, J.P.; Tkach, M.; Jouve, M.; Dingli, F.; Loew, D.; Witwer, K.W.; Ostrowski, M.; Borner, G.H.H.; et al. Unbiased proteomic profiling of host cell extracellular vesicle composition and dynamics upon HIV-1 infection. EMBO J. 2021, 40, e105492. [Google Scholar] [CrossRef] [PubMed]
- Orentas, R.J.; Hildreth, J.E.K. Association of Host Cell Surface Adhesion Receptors and Other Membrane Proteins with HIV and SIV. AIDS Res. Hum. Retrovir. 1993, 9, 1157–1165. [Google Scholar] [CrossRef] [PubMed]
- Bastiani, L.; Laal, S.; Kim, M.; Zolla-Pazner, S. Host cell-dependent alterations in envelope components of human immunodeficiency virus type 1 virions. J. Virol. 1997, 71, 3444–3450. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Frank, I.; Kacani, L.; Stoiber, H.; Stossel, H.; Spruth, M.; Steindl, F.; Romani, N.; Dierich, M.P. Human immunodeficiency virus type 1 derived from cocultures of immature dendritic cells with autologous T cells carries T-cell-specific molecules on its surface and is highly infectious. J. Virol. 1999, 73, 3449–3454. [Google Scholar] [CrossRef] [Green Version]
- Lawn, S.D.; Roberts, B.D.; Griffin, G.E.; Folks, T.M.; Butera, S.T. Cellular compartments of human immunodeficiency virus type 1 replication in vivo: Determination by presence of virion-associated host proteins and impact of opportunistic infection. J. Virol. 2000, 74, 139–145. [Google Scholar] [CrossRef] [Green Version]
- Chertova, E.; Chertov, O.; Coren, L.V.; Roser, J.D.; Trubey, C.M.; Bess, J.W.; Sowder, R.C.; Barsov, E.; Hood, B.L.; Fisher, R.J.; et al. Proteomic and biochemical analysis of purified human immunodeficiency virus type 1 produced from infected monocyte-derived macrophages. J. Virol. 2006, 80, 9039–9052. [Google Scholar] [CrossRef] [Green Version]
- Murakami, T.; Kim, J.; Li, Y.; Green, G.E.; Shikanov, A.; Ono, A. Secondary lymphoid organ fibroblastic reticular cells mediate trans-infection of HIV-1 via CD44-hyaluronan interactions. Nat. Commun. 2018, 9, 2436. [Google Scholar] [CrossRef]
- Liu, Y.; Fu, Y.; Wang, Q.; Li, M.; Zhou, Z.; Dabbagh, D.; Fu, C.; Zhang, H.; Li, S.; Zhang, T.; et al. Proteomic profiling of HIV-1 infection of human CD4+ T cells identifies PSGL-1 as an HIV restriction factor. Nat. Microbiol. 2019, 4, 813–825. [Google Scholar] [CrossRef]
- Fu, Y.; He, S.; Waheed, A.A.; Dabbagh, D.; Zhou, Z.; Trinité, B.; Wang, Z.; Yu, J.; Wang, D.; Li, F.; et al. PSGL-1 restricts HIV-1 infectivity by blocking virus particle attachment to target cells. Proc. Natl. Acad. Sci. USA 2020, 117, 9537–9545. [Google Scholar] [CrossRef] [Green Version]
- Murakami, T.; Carmona, N.; Ono, A. Virion-incorporated PSGL-1 and CD43 inhibit both cell-free infection and transinfection of HIV-1 by preventing virus–cell binding. Proc. Natl. Acad. Sci. USA 2020, 117, 8055–8063. [Google Scholar] [CrossRef]
- Shao, B.; Yago, T.; Setiadi, H.; Wang, Y.; Mehta-D’Souza, P.; Fu, J.; Crocker, P.R.; Rodgers, W.; Xia, L.; McEver, R.P. O-glycans direct selectin ligands to lipid rafts on leukocytes. Proc. Natl. Acad. Sci. USA 2015, 112, 8661–8666. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zarbock, A.; Ley, K.; McEver, R.P.; Hidalgo, A. Leukocyte ligands for endothelial selectins: Specialized glycoconjugates that mediate rolling and signaling under flow. Blood 2011, 118, 6743–6751. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grover, J.R.; Veatch, S.L.; Ono, A. Basic Motifs Target PSGL-1, CD43, and CD44 to Plasma Membrane Sites Where HIV-1 Assembles. J. Virol. 2015, 89, 454–467. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chan, R.; Uchil, P.; Jin, J.; Shui, G.; Ott, D.E.; Mothes, W.; Wenk, M.R. Retroviruses Human Immunodeficiency Virus and Murine Leukemia Virus Are Enriched in Phosphoinositides. J. Virol. 2008, 82, 11228–11238. [Google Scholar] [CrossRef] [Green Version]
- Favard, C.; Chojnacki, J.; Merida, P.; Yandrapalli, N.; Mak, J.; Eggeling, C.; Muriaux, D. HIV-1 Gag specifically restricts PI (4,5) P2 and cholesterol mobility in living cells creating a nanodomain platform for virus assembly. Sci. Adv. 2019, 5, eaaw8651. [Google Scholar] [CrossRef] [Green Version]
- Mücksch, F.; Citir, M.; Lüchtenborg, C.; Glass, B.; Traynor-Kaplan, A.; Schultz, C.; Brügger, B.; Kräusslich, H.-G. Quantification of phosphoinositides reveals strong enrichment of PIP2 in HIV-1 compared to producer cell membranes. Sci. Rep. 2019, 9, 1–13. [Google Scholar] [CrossRef]
- Ono, A.; Ablan, S.D.; Lockett, S.J.; Nagashima, K.; Freed, E.O. Phosphatidylinositol (4,5) bisphosphate regulates HIV-1 Gag targeting to the plasma membrane. Proc. Natl. Acad. Sci. USA 2004, 101, 14889–14894. [Google Scholar] [CrossRef] [Green Version]
- Saad, J.; Miller, J.; Tai, J.; Kim, A.; Ghanam, R.H.; Summers, M.F. Structural basis for targeting HIV-1 Gag proteins to the plasma membrane for virus assembly. Proc. Natl. Acad. Sci. USA 2006, 103, 11364–11369. [Google Scholar] [CrossRef] [Green Version]
- Shkriabai, N.; Datta, S.; Zhao, Z.; Hess, S.; Rein, A.; Kvaratskhelia, M. Interactions of HIV-1 Gag with Assembly Cofactors. Biochemistry 2006, 45, 4077–4083. [Google Scholar] [CrossRef]
- Chukkapalli, V.; Hogue, I.; Boyko, V.; Hu, W.-S.; Ono, A. Interaction between the Human Immunodeficiency Virus Type 1 Gag Matrix Domain and Phosphatidylinositol-(4,5)-Bisphosphate Is Essential for Efficient Gag Membrane Binding. J. Virol. 2008, 82, 2405–2417. [Google Scholar] [CrossRef] [Green Version]
- Alfadhli, A.; Barklis, R.L.; Barklis, E. HIV-1 matrix organizes as a hexamer of trimers on membranes containing phosphatidylinositol-(4,5)-bisphosphate. Virology 2009, 387, 466–472. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alfadhli, A.; Still, A.; Barklis, E. Analysis of Human Immunodeficiency Virus Type 1 Matrix Binding to Membranes and Nucleic Acids. J. Virol. 2009, 83, 12196–12203. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Anraku, K.; Fukuda, R.; Takamune, N.; Misumi, S.; Okamoto, Y.; Otsuka, M.; Fujita, M. Highly Sensitive Analysis of the Interaction between HIV-1 Gag and Phosphoinositide Derivatives Based on Surface Plasmon Resonance. Biochemistry 2010, 49, 5109–5116. [Google Scholar] [CrossRef] [PubMed]
- Chukkapalli, V.; Oh, S.J.; Ono, A. Opposing mechanisms involving RNA and lipids regulate HIV-1 Gag membrane binding through the highly basic region of the matrix domain. Proc. Natl. Acad. Sci. USA 2010, 107, 1600–1605. [Google Scholar] [CrossRef] [Green Version]
- Chukkapalli, V.; Ono, A. Molecular Determinants that Regulate Plasma Membrane Association of HIV-1 Gag. J. Mol. Biol. 2011, 410, 512–524. [Google Scholar] [CrossRef] [Green Version]
- Inlora, J.; Chukkapalli, V.; Derse, D.; Ono, A. Gag localization and virus-like particle release mediated by the matrix domain of human t-lymphotropic virus type 1 gag are less dependent on phosphatidylinositol-(4,5)-bisphosphate than those mediated by the matrix domain of HIV-1 Gag. J. Virol. 2011, 85, 3802–3810. [Google Scholar] [CrossRef] [Green Version]
- Inlora, J.; Collins, D.R.; Trubin, M.E.; Chung, J.Y.J.; Ono, A. Membrane binding and subcellular localization of retroviral gag proteins are differentially regulated by ma interactions with phosphatidylinositol-(4,5)-bisphosphate and RNA. mBio 2014, 5, e02202. [Google Scholar] [CrossRef] [Green Version]
- Olety, B.; Veatch, S.L.; Ono, A. Phosphatidylinositol-(4,5)-Bisphosphate Acyl Chains Differentiate Membrane Binding of HIV-1 Gag from That of the Phospholipase Cdelta1 Pleckstrin Homology Domain. J. Virol. 2015, 89, 7861–7873. [Google Scholar] [CrossRef] [Green Version]
- Inlora, J.; Chukkapalli, V.; Bedi, S.; Ono, A. Molecular Determinants Directing HIV-1 Gag Assembly to Virus-Containing Compartments in Primary Macrophages. J. Virol. 2016, 90, 8509–8519. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thornhill, D.; Olety, B.; Ono, A. Relationships between MA-RNA Binding in Cells and Suppression of HIV-1 Gag Mislocalization to Intracellular Membranes. J. Virol. 2019, 93. [Google Scholar] [CrossRef]
- Thornhill, D.; Murakami, T.; Ono, A. Rendezvous at Plasma Membrane: Cellular Lipids and tRNA Set up Sites of HIV-1 Particle Assembly and Incorporation of Host Transmembrane Proteins. Viruses 2020, 12, 842. [Google Scholar] [CrossRef] [PubMed]
- Zhou, W.; Parent, L.J.; Wills, J.W.; Resh, M.D. Identification of a membrane-binding domain within the amino-terminal region of human immunodeficiency virus type 1 Gag protein which interacts with acidic phospholipids. J. Virol. 1994, 68, 2556–2569. [Google Scholar] [CrossRef] [Green Version]
- Ehrlich, L.S.; Fong, S.; Scarlata, S.; Zybarth, A.G.; Carter, C. Partitioning of HIV-1 Gag and Gag-Related Proteins to Membranes. Biochemistry 1996, 35, 3933–3943. [Google Scholar] [CrossRef] [PubMed]
- Dalton, A.K.; Ako-Adjei, D.; Murray, P.S.; Murray, D.; Vogt, V.M. Electrostatic Interactions Drive Membrane Association of the Human Immunodeficiency Virus Type 1 Gag MA Domain. J. Virol. 2007, 81, 6434–6445. [Google Scholar] [CrossRef] [Green Version]
- Wen, Y.; Feigenson, G.W.; Vogt, V.M.; Dick, R.A. Mechanisms of PI (4,5) P2 Enrichment in HIV-1 Viral Membranes. J. Mol. Biol. 2020, 432, 5343–5364. [Google Scholar] [CrossRef]
- Chen, X.; Khajeh, J.A.; Ju, J.H.; Gupta, Y.K.; Stanley, C.; Do, C.; Heller, W.; Aggarwal, A.K.; Callaway, D.J.E.; Bu, Z. Phosphatidylinositol 4,5-Bisphosphate Clusters the Cell Adhesion Molecule CD44 and Assembles a Specific CD44-Ezrin Heterocomplex, as Revealed by Small Angle Neutron Scattering. J. Biol. Chem. 2015, 290, 6639–6652. [Google Scholar] [CrossRef] [Green Version]
- Purohit, P.; Dupont, S.; Stevenson, M.; Green, M.R. Sequence-specific interaction between HIV-1 matrix protein and viral genomic RNA revealed by in vitro genetic selection. RNA 2001, 7, 576–584. [Google Scholar] [CrossRef] [Green Version]
- Hearps, A.C.; Wagstaff, K.; Piller, S.C.; Jans, D. The N-terminal basic domain of the HIV-1 matrix protein does not contain a conventional nuclear localization sequence but is required for DNA binding and protein self-association. Biochemistry 2008, 47, 2199–2210. [Google Scholar] [CrossRef] [PubMed]
- Jones, C.P.; Datta, S.A.; Rein, A.; Rouzina, I.; Musier-Forsyth, K. Matrix domain modulates HIV-1 Gag′s nucleic acid chaperone activity via inositol phosphate binding. J. Virol. 2011, 85, 1594–1603. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chukkapalli, V.; Inlora, J.; Todd, G.C.; Ono, A. Evidence in Support of RNA-Mediated Inhibition of Phosphatidylserine-Dependent HIV-1 Gag Membrane Binding in Cells. J. Virol. 2013, 87, 7155–7159. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kutluay, S.B.; Zang, T.; Blanco-Melo, D.; Powell, C.; Jannain, D.; Errando, M.; Bieniasz, P.D. Global Changes in the RNA Binding Specificity of HIV-1 Gag Regulate Virion Genesis. Cell 2014, 159, 1096–1109. [Google Scholar] [CrossRef] [Green Version]
- Todd, G.C.; Duchon, A.; Inlora, J.; Olson, E.D.; Musier-Forsyth, K.; Ono, A. Inhibition of HIV-1 Gag–membrane interactions by specific RNAs. RNA 2016, 23, 395–405. [Google Scholar] [CrossRef] [Green Version]
- Gaines, C.R.; Tkacik, E.; Rivera-Oven, A.; Somani, P.; Achimovich, A.; Alabi, T.; Zhu, A.; Getachew, N.; Yang, A.L.; McDonough, M.; et al. HIV-1 Matrix Protein Interactions with tRNA: Implications for Membrane Targeting. J. Mol. Biol. 2018, 430, 2113–2127. [Google Scholar] [CrossRef] [PubMed]
- Kroupa, T.; Datta, S.A.K.; Rein, A. Distinct Contributions of Different Domains within the HIV-1 Gag Polyprotein to Specific and Nonspecific Interactions with RNA. Viruses 2020, 12, 394. [Google Scholar] [CrossRef] [Green Version]
- Hart, S.P.; Dougherty, G.J.; Haslett, C.; Dransfield, I. CD44 regulates phagocytosis of apoptotic neutrophil granulocytes, but not apoptotic lymphocytes, by human macrophages. J. Immunol. 1997, 159. [Google Scholar]
- Toyos, J.D.L.; Jalkanen, S.; Butcher, E.C. Flow cytometric analysis of the Hermes homing-associated antigen on human lymphocyte subsets. Blood 1989, 74, 751–760. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aruffo, A.; Stamenkovic, I.; Melnick, M.; Underhill, C.B.; Seed, B. CD44 is the principal cell surface receptor for hyaluronate. Cell 1990, 61, 1303–1313. [Google Scholar] [CrossRef]
- Borland, G.A.; Ross, J.; Guy, K. Forms and functions of CD44. Immunology 1998, 93, 139–148. [Google Scholar] [CrossRef]
- Zoller, M. CD44, Hyaluronan, the hematopoietic stem cell, and leukemia-initiating cells. Front. Immunol. 2015, 6, 235. [Google Scholar]
- Bennett, K.L.; Modrell, B.; Greenfield, B.; Bartolazzi, A.; Stamenkovic, I.; Peach, R.; Jackson, D.G.; Spring, F.; Aruffo, A. Regulation of CD44 binding to hyaluronan by glycosylation of variably spliced exons. J. Cell Biol. 1995, 131, 1623–1633. [Google Scholar] [CrossRef]
- Katoh, S.; Zheng, Z.; Oritani, K.; Shimozato, T.; Kincade, P.W. Glycosylation of CD44 negatively regulates its recognition of hyaluronan. J. Exp. Med. 1995, 182, 419–429. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, L.; Fu, C.; Zhang, Q.; He, C.; Zhang, F.; Wei, Q. The role of CD44 in pathological angiogenesis. FASEB J. 2020, 34, 13125–13139. [Google Scholar] [CrossRef]
- Degrendele, H.C.; Estess, P.; Picker, L.J.; Siegelman, M.H. CD44 and its ligand hyaluronate mediate rolling under physiologic flow: A novel lymphocyte-endothelial cell primary adhesion pathway. J. Exp. Med. 1996, 183, 1119–1130. [Google Scholar] [CrossRef] [Green Version]
- DeGrendele, H.C.; Estess, P.; Siegelman, M.H. Requirement for CD44 in Activated T Cell Extravasation into an Inflammatory Site. Science 1997, 278, 672–675. [Google Scholar] [CrossRef]
- Degrendele, H.C.; Kosfiszer, M.; Estess, P.; Siegelman, M.H. CD44 activation and associated primary adhesion is inducible via T cell receptor stimulation. J. Immunol. 1997, 159, 2549–2553. [Google Scholar]
- Bonder, C.S.; Clark, S.R.; Norman, M.U.; Johnson, P.; Kubes, P. Use of CD44 by CD4+ Th1 and Th2 lymphocytes to roll and adhere. Blood 2006, 107, 4798–4806. [Google Scholar] [CrossRef] [Green Version]
- Nácher, M.; Blázquez, A.B.; Shao, B.; Matesanz, A.; Prophete, C.; Berin, M.C.; Frenette, P.S.; Hidalgo, A. Physiological Contribution of CD44 as a Ligand for E-Selectin during Inflammatory T-Cell Recruitment. Am. J. Pathol. 2011, 178, 2437–2446. [Google Scholar] [CrossRef] [Green Version]
- Terry, V.H.; Johnston, I.C.; Spina, C.A. CD44 MicroBeads accelerate HIV-1 infection in T cells. Virology 2009, 388, 294–304. [Google Scholar] [CrossRef] [Green Version]
- Burnie, J.; Persaud, A.T.; Thaya, L.; Liu, Q.; Miao, H.; Grabinsky, S.; Norouzi, V.; Lusso, P.; Tang, V.A.; Guzzo, C. The P-selectin ligand PSGL-1 (CD162) is efficiently incorporated by primary HIV-1 isolates and can facilitate trans-infection. bioRxiv 2021. [Google Scholar] [CrossRef]
- Bajénoff, M.; Egen, J.; Qi, H.; Huang, A.; Castellino, F.; Germain, R.N. Highways, byways and breadcrumbs: Directing lymphocyte traffic in the lymph node. Trends Immunol. 2007, 28, 346–352. [Google Scholar] [CrossRef]
- Cremasco, V.; Woodruff, M.C.; Onder, L.; Cupovic, J.; Nieves-Bonilla, J.; Schildberg, F.A.; Chang, J.; Cremasco, F.; Harvey, C.J.; Wucherpfennig, K.; et al. B cell homeostasis and follicle confines are governed by fibroblastic reticular cells. Nat. Immunol. 2014, 15, 973–981. [Google Scholar] [CrossRef] [PubMed]
- Geijtenbeek, T.B.; Kwon, D.S.; Torensma, R.; van Vliet, S.J.; van Duijnhoven, G.C.; Middel, J.; Cornelissen, I.L.; Nottet, H.S.; KewalRamani, V.N.; Littman, D.R.; et al. DC-SIGN, a Dendritic Cell–Specific HIV-1-Binding Protein that Enhances trans-Infection of T Cells. Cell 2000, 100, 587–597. [Google Scholar] [CrossRef] [Green Version]
- Rappocciolo, G.; Piazza, P.; Fuller, C.L.; Reinhart, T.A.; Watkins, S.C.; Rowe, D.T.; Jais, M.; Supta, P.; Rinaldo, C.R., Jr. DC-SIGN on B lymphocytes is required for transmission of HIV-1 to T lymphocytes. PLoS Pathog. 2006, 2, e70. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Izquierdo-Useros, N.; Blanco, J.; Erkizia, I.; Fernández-Figueras, M.T.; Borras, F.E.; Naranjo-Gomez, M.; Bofill, M.; Ruiz, L.; Clotet, B.; Martinez-Picado, J. Maturation of Blood-Derived Dendritic Cells Enhances Human Immunodeficiency Virus Type 1 Capture and Transmission. J. Virol. 2007, 81, 7559–7570. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sewald, X.; Ladinsky, M.S.; Uchil, P.; Beloor, J.; Pi, R.; Herrmann, C.; Motamedi, N.; Murooka, T.; Brehm, M.; Greiner, D.L.; et al. Retroviruses use CD169-mediated trans-infection of permissive lymphocytes to establish infection. Science 2015, 350, 563–567. [Google Scholar] [CrossRef] [Green Version]
- Izquierdo-Useros, N.; Lorizate, M.; Puertas, M.C.; Rodriguez-Plata, M.T.; Zangger, N.; Erikson, E.; Pino, M.; Erkizia, I.; Glass, B.; Clotet, B.; et al. Siglec-1 Is a Novel Dendritic Cell Receptor That Mediates HIV-1 Trans-Infection Through Recognition of Viral Membrane Gangliosides. PLoS Biol. 2012, 10, e1001448. [Google Scholar] [CrossRef]
- Puryear, W.B.; Akiyama, H.; Geer, S.D.; Ramirez, N.-G.; Yu, X.; Reinhard, B.M.; Gummuluru, S. Interferon-Inducible Mechanism of Dendritic Cell-Mediated HIV-1 Dissemination Is Dependent on Siglec-1/CD169. PLoS Pathog. 2013, 9, e1003291. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hammonds, J.E.; Beeman, N.; Ding, L.; Takushi, S.; Francis, A.C.; Wang, J.-J.; Melikyan, G.B.; Spearman, P. Siglec-1 initiates formation of the virus-containing compartment and enhances macrophage-to-T cell transmission of HIV-1. PLoS Pathog. 2017, 13, e1006181. [Google Scholar] [CrossRef] [PubMed]
- Kwon, D.S.; Gregorio, G.G.; Bitton, N.; Hendrickson, W.A.; Littman, D.R. DC-SIGN-Mediated Internalization of HIV Is Required for Trans-Enhancement of T Cell Infection. Immunity 2002, 16, 135–144. [Google Scholar] [CrossRef] [Green Version]
- Boggiano, C.; Manel, N.; Littman, D.R. Dendritic Cell-Mediated trans -Enhancement of Human Immunodeficiency Virus Type 1 Infectivity Is Independent of DC-SIGN. J. Virol. 2007, 81, 2519–2523. [Google Scholar] [CrossRef] [Green Version]
- Culty, M.; Nguyen, H.A.; Underhill, C.B. The hyaluronan receptor (CD44) participates in the uptake and degradation of hyaluronan. J. Cell Biol. 1992, 116, 1055–1062. [Google Scholar] [CrossRef] [Green Version]
- Cavrois, M.; Neidleman, J.; Kreisberg, J.; Greene, W.C. In Vitro Derived Dendritic Cells Trans-Infect CD4 T Cells Primarily with Surface-Bound HIV-1 Virions. PLoS Pathog. 2007, 3, e4. [Google Scholar] [CrossRef] [Green Version]
- Yu, H.J.; Reuter, M.A.; McDonald, D. HIV Traffics through a Specialized, Surface-Accessible Intracellular Compartment during trans-Infection of T Cells by Mature Dendritic Cells. PLoS Pathog. 2008, 4, e1000134. [Google Scholar] [CrossRef] [Green Version]
- Akiyama, H.; Ramirez, N.-G.; Gudheti, M.V.; Gummuluru, S. CD169-Mediated Trafficking of HIV to Plasma Membrane Invaginations in Dendritic Cells Attenuates Efficacy of Anti-gp120 Broadly Neutralizing Antibodies. PLoS Pathog. 2015, 11, e1004751. [Google Scholar] [CrossRef]
- Ménager, M.; Littman, D.R. Actin Dynamics Regulates Dendritic Cell-Mediated Transfer of HIV-1 to T Cells. Cell 2016, 164, 695–709. [Google Scholar] [CrossRef] [Green Version]
- Deneka, M.; Pelchen-Matthews, A.; Byland, R.; Ruiz-Mateos, E.; Marsh, M. In macrophages, HIV-1 assembles into an intracellular plasma membrane domain containing the tetraspanins CD81, CD9, and CD53. J. Cell Biol. 2007, 177, 329–341. [Google Scholar] [CrossRef]
- Bennett, A.E.; Narayan, K.; Shi, D.; Hartnell, L.M.; Gousset, K.; He, H.; Lowekamp, B.C.; Yoo, T.S.; Bliss, D.; Freed, E.O.; et al. Ion-Abrasion Scanning Electron Microscopy Reveals Surface-Connected Tubular Conduits in HIV-Infected Macrophages. PLoS Pathog. 2009, 5, e1000591. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chu, H.; Wang, J.-J.; Qi, M.; Yoon, J.-J.; Wen, X.; Chen, X.; Ding, L.; Spearman, P. The Intracellular Virus-Containing Compartments in Primary Human Macrophages Are Largely Inaccessible to Antibodies and Small Molecules. PLoS ONE 2012, 7, e35297. [Google Scholar] [CrossRef] [PubMed]
- Rosenblum, M.D.; Way, S.S.; Abbas, A.K. Regulatory T cell memory. Nat. Rev. Immunol. 2015, 16, 90–101. [Google Scholar] [CrossRef] [PubMed]
- Cornelissen, M.; Heeregrave, E.J.; Zorgdrager, F.; Pollakis, G.; Paxton, W.A.; Van Der Kuyl, A.C. Generation of representative primary virus isolates from blood plasma after isolation of HIV-1 with CD44 MicroBeads. Arch. Virol. 2010, 155, 2017–2022. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, P.; Fujimoto, K.; Bourguingnon, L.; Yukl, S.; Deeks, S.; Wong, J.K. Exogenous and endogenous hyaluronic acid reduces HIV infection of CD4 + T cells. Immunol. Cell Biol. 2014, 92, 770–780. [Google Scholar] [CrossRef] [PubMed]
- Neidleman, J.A.; Chen, J.C.; Kohgadai, N.; Müller, J.A.; Laustsen, A.; Thavachelvam, K.; Jan, M.; Stürzel, C.M.; Jones, J.J.; Ochsenbauer, C.; et al. Mucosal stromal fibroblasts markedly enhance HIV infection of CD4+ T cells. PLoS Pathog. 2017, 13, e1006163. [Google Scholar] [CrossRef]
- Egedal, J.H.; Xie, G.; Packard, T.A.; Laustsen, A.; Neidleman, J.; Georgiou, K.; Pillai, S.K.; Greene, W.C.; Jakobsen, M.R.; Roan, N.R. Hyaluronic acid is a negative regulator of mucosal fibroblast-mediated enhancement of HIV infection. Mucosal Immunol. 2021, 14, 1203–1213. [Google Scholar] [CrossRef]
- Colomer-Lluch, M.; Ruiz, A.; Moris, A.; Prado, J.G. Restriction Factors: From Intrinsic Viral Restriction to Shaping Cellular Immunity Against HIV-1. Front. Immunol. 2018, 9, 2876. [Google Scholar] [CrossRef] [Green Version]
- Ramirez, P.; Sharma, S.; Singh, R.; Stoneham, C.; Vollbrecht, T.; Guatelli, J. Plasma Membrane-Associated Restriction Factors and Their Counteraction by HIV-1 Accessory Proteins. Cells 2019, 8, 1020. [Google Scholar] [CrossRef] [Green Version]
- Haller, C.; Müller, B.; Fritz, J.V.; Lamas-Murua, M.; Stolp, B.; Pujol, F.M.; Keppler, O.T.; Fackler, O.T. HIV-1 Nef and Vpu Are Functionally Redundant Broad-Spectrum Modulators of Cell Surface Receptors, Including Tetraspanins. J. Virol. 2014, 88, 14241–14257. [Google Scholar] [CrossRef] [Green Version]
- Matheson, N.J.; Sumner, J.; Wals, K.; Rapiteanu, R.; Weekes, M.P.; Vigan, R.; Weinelt, J.; Schindler, M.; Antrobus, R.; Costa, A.S.; et al. Cell Surface Proteomic Map of HIV Infection Reveals Antagonism of Amino Acid Metabolism by Vpu and Nef. Cell Host Microbe 2015, 18, 409–423. [Google Scholar] [CrossRef] [Green Version]
- McLaren, P.J.; Gawanbacht, A.; Pyndiah, N.; Krapp, C.; Hotter, D.; Kluge, S.F.; Götz, N.; Heilmann, J.; Mack, K.; Sauter, D.; et al. Identification of potential HIV restriction factors by combining evolutionary genomic signatures with functional analyses. Retrovirology 2015, 12, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Hirata, T.; Furukawa, Y.; Yang, B.-G.; Hieshima, K.; Fukuda, M.; Kannagi, R.; Yoshie, O.; Miyasaka, M. Human P-selectin Glycoprotein Ligand-1 (PSGL-1) Interacts with the Skin-associated Chemokine CCL27 via Sulfated Tyrosines at the PSGL-1 Amino Terminus. J. Biol. Chem. 2004, 279, 51775–51782. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Veerman, K.M.; Williams, M.J.; Uchimura, K.; Singer, M.S.; Merzaban, J.S.; Naus, S.; A Carlow, D.A.; Owen, P.; Rivera-Nieves, J.; Rosen, S.D.; et al. Interaction of the selectin ligand PSGL-1 with chemokines CCL21 and CCL19 facilitates efficient homing of T cells to secondary lymphoid organs. Nat. Immunol. 2007, 8, 532–539. [Google Scholar] [CrossRef]
- Ladinsky, M.S.; Gnanapragasam, P.N.; Yang, Z.; West, A.P.; Kay, M.S.; Bjorkman, P.J. Electron tomography visualization of HIV-1 fusion with target cells using fusion inhibitors to trap the pre-hairpin intermediate. eLife 2020, 9, e58411. [Google Scholar] [CrossRef] [PubMed]
- Dabbagh, D.; He, S.; Hetrick, B.; Chilin, L.; Andalibi, A.; Wu, Y. Identification of the SHREK Family of Proteins as Broad-Spectrum Host Antiviral Factors. Viruses 2021, 13, 832. [Google Scholar] [CrossRef] [PubMed]
- He, S.; Waheed, A.A.; Hetrick, B.; Dabbagh, D.; Akhrymuk, I.V.; Kehn-Hall, K.; Freed, E.O.; Wu, Y. PSGL-1 Inhibits the Incorporation of SARS-CoV and SARS-CoV-2 Spike Glycoproteins into Pseudovirions and Impairs Pseudovirus Attachment and Infectivity. Viruses 2020, 13, 46. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Song, Y.; Zhang, S.; Diao, M.; Huang, S.; Li, S.; Tan, X. PSGL-1 inhibits HIV-1 infection by restricting actin dynamics and sequestering HIV envelope proteins. Cell Discov. 2020, 6, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Pantaleo, G.; Graziosi, C.; Butini, L.; Pizzo, P.A.; Schnittman, S.M.; Kotler, D.P.; Fauci, A.S. Lymphoid organs function as major reservoirs for human immunodeficiency virus. Proc. Natl. Acad. Sci. USA 1991, 88, 9838–9842. [Google Scholar] [CrossRef] [Green Version]
- Pantaleo, G.; Graziosi, C.; Demarest, J.F.; Butini, L.; Montroni, M.; Fox, C.H.; Orenstein, J.M.; Kotler, D.P.; Fauci, A.S. HIV infection is active and progressive in lymphoid tissue during the clinically latent stage of disease. Nature 1993, 362, 355–358. [Google Scholar] [CrossRef]
- Fox, C.H.; Tenner-Racz, K.; Racz, P.; Firpo, A.; Pizzo, P.A.; Fauci, A.S. Lymphoid Germinal Centers Are Reservoirs of Human Immunodeficiency Virus Type 1 RNA. J. Infect. Dis. 1991, 164, 1051–1057. [Google Scholar] [CrossRef]
- Embretson, J.; Zupancic, M.; Ribas, J.L.; Burke, A.; Racz, P.; Tenner-Racz, K.; Haase, A.T. Massive covert infection of helper T lymphocytes and macrophages by HIV during the incubation period of AIDS. Nature 1993, 362, 359–362. [Google Scholar] [CrossRef] [PubMed]
- Heath, S.L.; Tew, J.G.; Szakal, A.K.; Burton, G.F. Follicular dendritic cells and human immunodeficiency virus infectivity. Nature 1995, 377, 740–744. [Google Scholar] [CrossRef] [PubMed]
- Murooka, T.; Deruaz, M.; Marangoni, F.; Vrbanac, V.D.; Seung, E.; Von Andrian, U.H.; Tager, A.M.; Luster, A.D.; Mempel, T.R. HIV-infected T cells are migratory vehicles for viral dissemination. Nature 2012, 490, 283–287. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sewald, X.; Gonzalez, D.G.; Haberman, A.M.; Mothes, W. In vivo imaging of virological synapses. Nat. Commun. 2012, 3, 1–9. [Google Scholar] [CrossRef]
- Law, K.M.; Komarova, N.L.; Yewdall, A.W.; Lee, R.K.; Herrera, O.L.; Wodarz, D.; Chen, B.K. In Vivo HIV-1 Cell-to-Cell Transmission Promotes Multicopy Micro-compartmentalized Infection. Cell Rep. 2016, 15, 2771–2783. [Google Scholar] [CrossRef] [Green Version]


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Murakami, T.; Ono, A. Roles of Virion-Incorporated CD162 (PSGL-1), CD43, and CD44 in HIV-1 Infection of T Cells. Viruses 2021, 13, 1935. https://doi.org/10.3390/v13101935
Murakami T, Ono A. Roles of Virion-Incorporated CD162 (PSGL-1), CD43, and CD44 in HIV-1 Infection of T Cells. Viruses. 2021; 13(10):1935. https://doi.org/10.3390/v13101935
Chicago/Turabian StyleMurakami, Tomoyuki, and Akira Ono. 2021. "Roles of Virion-Incorporated CD162 (PSGL-1), CD43, and CD44 in HIV-1 Infection of T Cells" Viruses 13, no. 10: 1935. https://doi.org/10.3390/v13101935