
Chlorovirus PBCV-1 Multidomain Protein A111/114R Has Three Glycosyltransferase Functions Involved in the Synthesis of Atypical N-Glycans

 ,
,  , , and
, , and 
Abstract
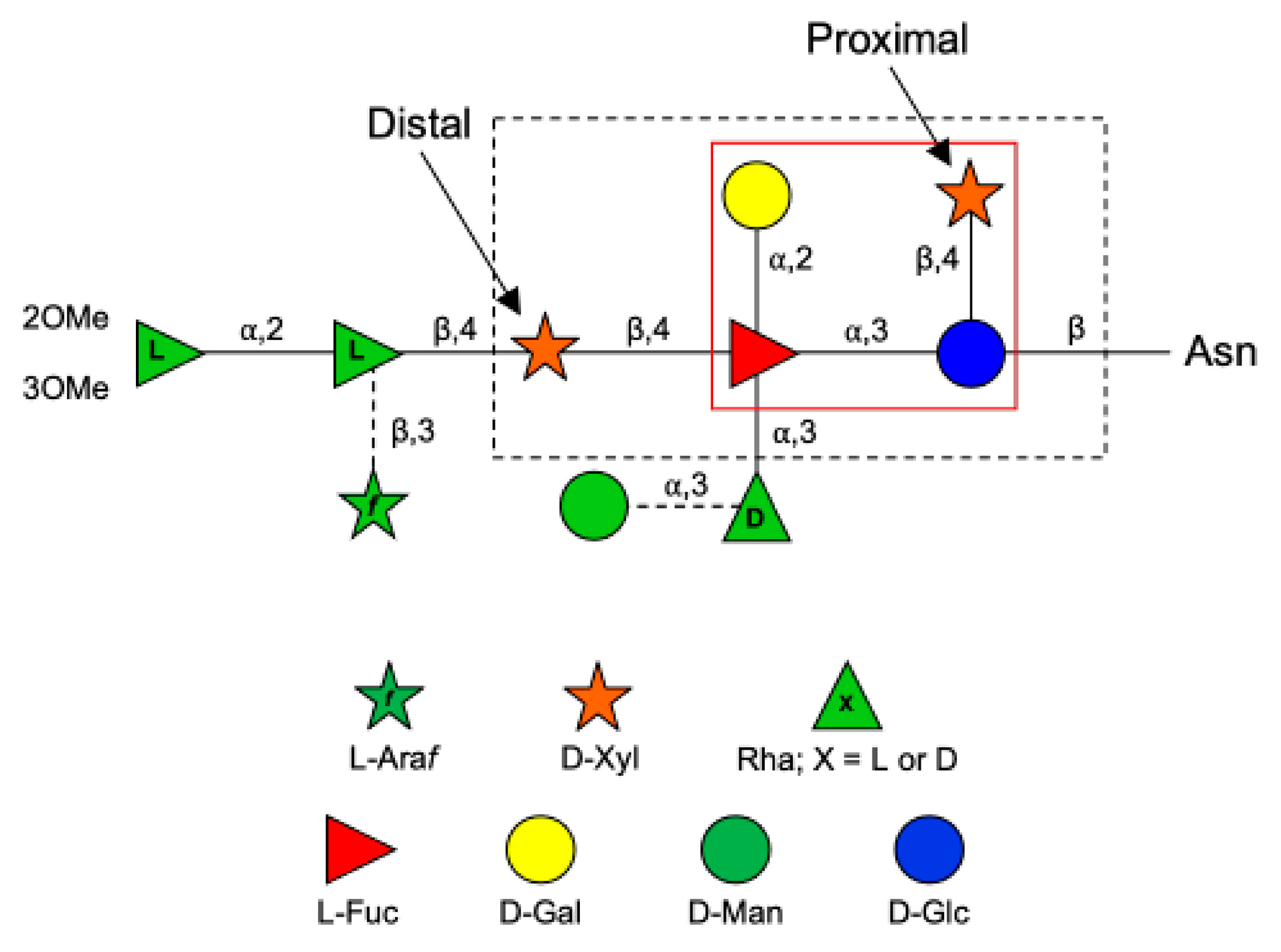
:1. Introduction
2. Materials and Methods
2.1. Protein Modeling
2.2. Cloning and Expression
2.3. Purification of Recombinant Enzymes
2.4. UDP-GloTM and GDP-GloTM GT Assays
3. Results and Discussion
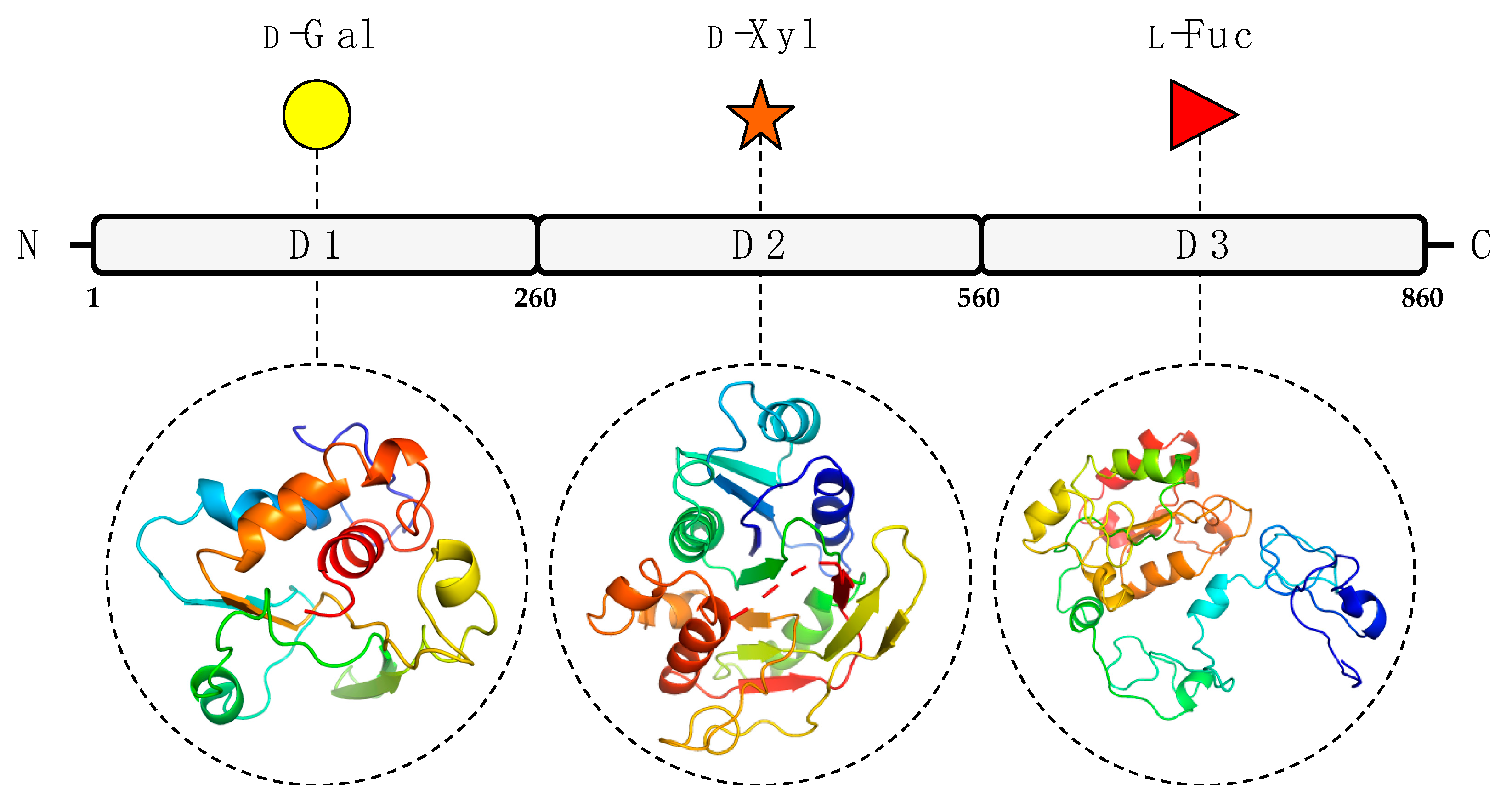
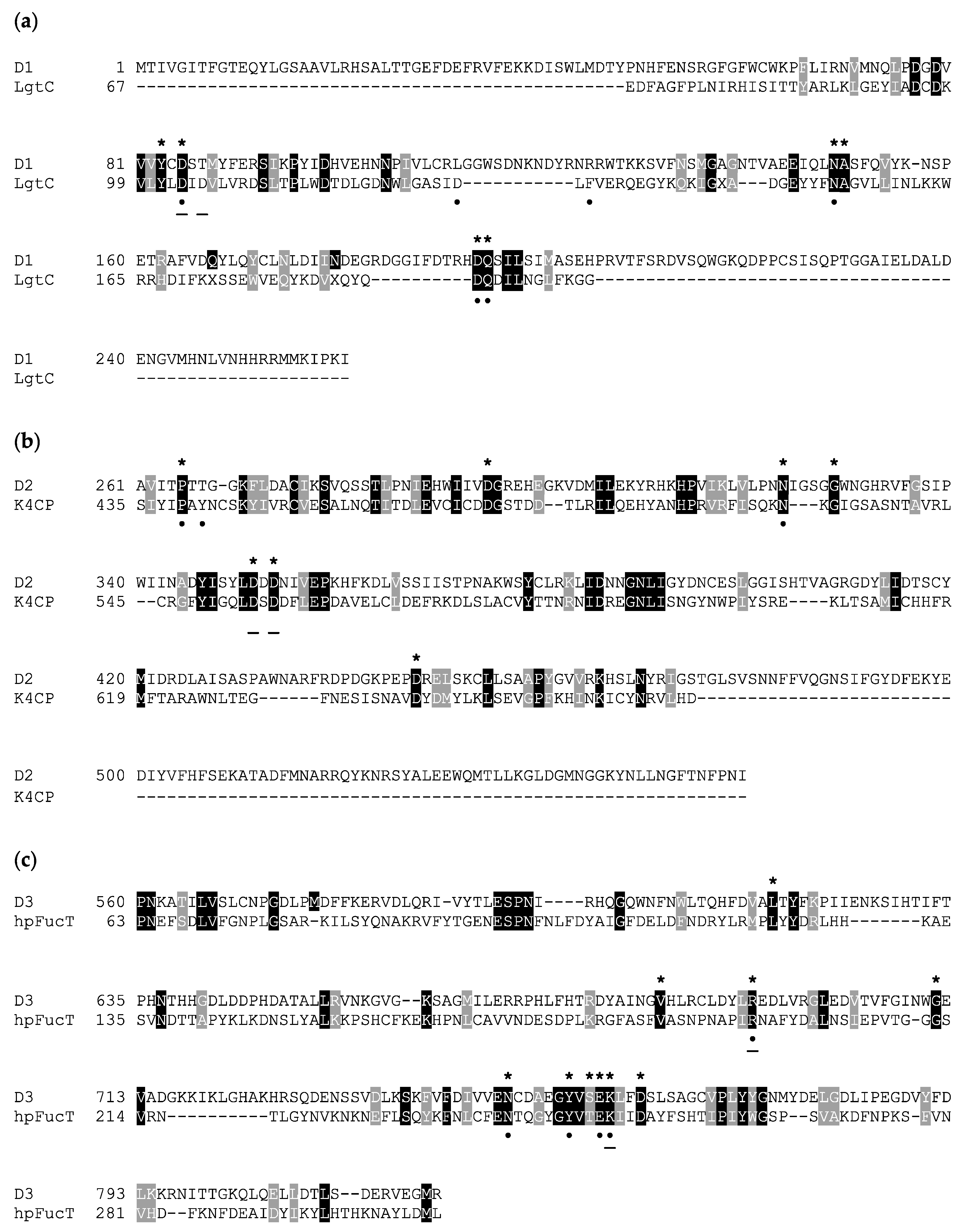
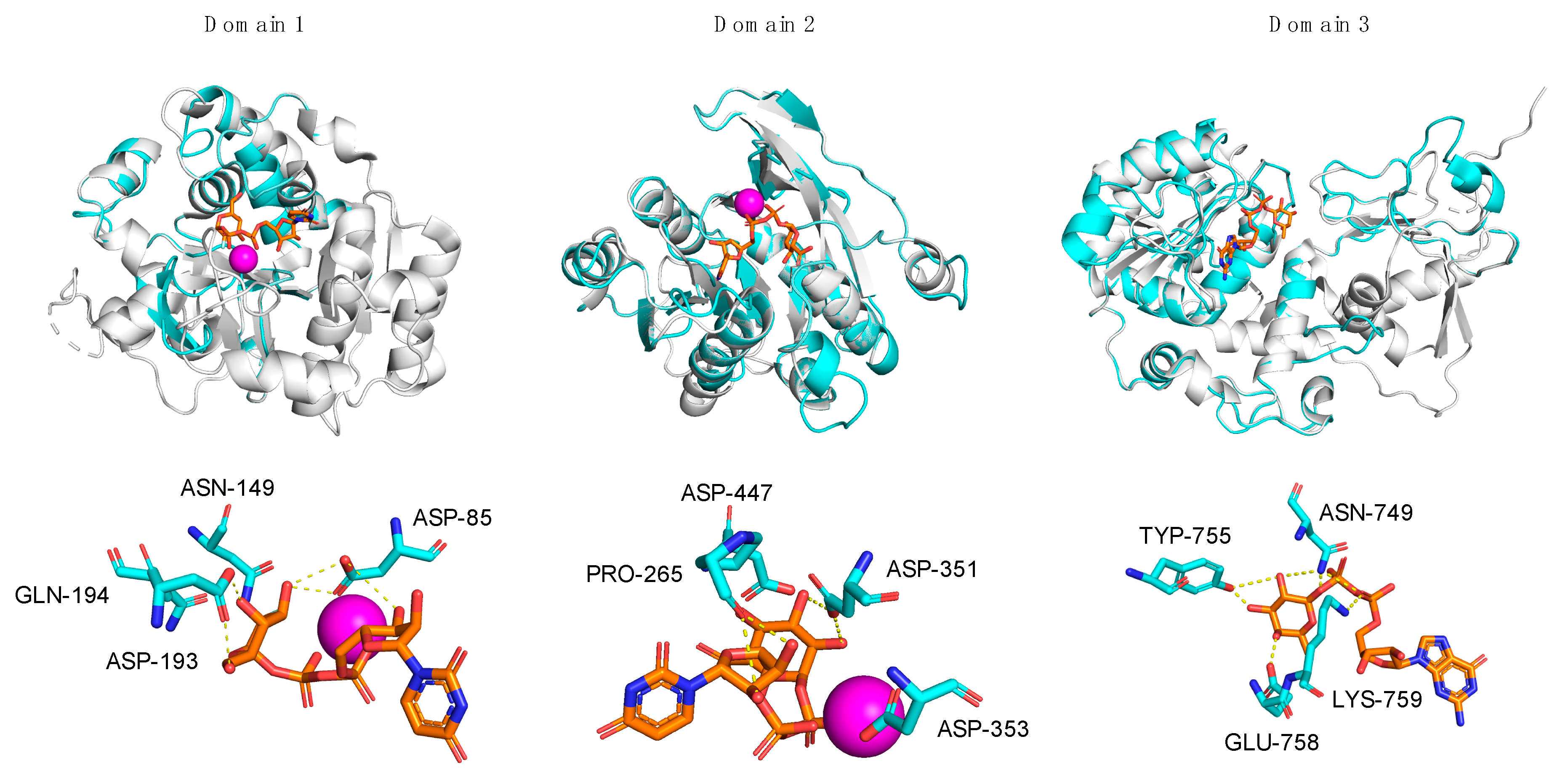
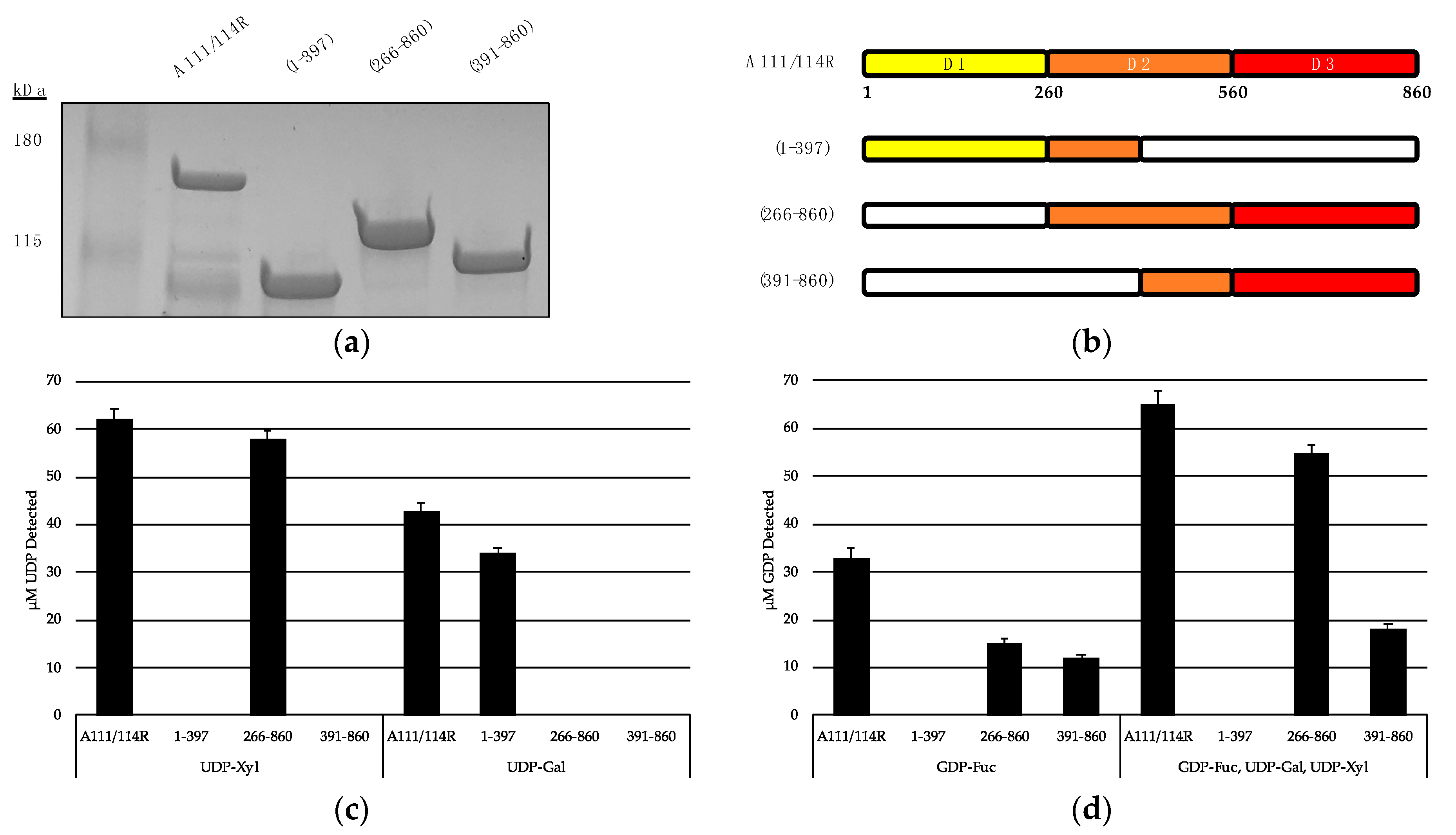
3.1. In Silico Analysis of A111/114R Domain 1
3.2. In Silico Analysis of A111/114R Domain 2
3.3. In Silico Analysis of A111/114R Domain 3
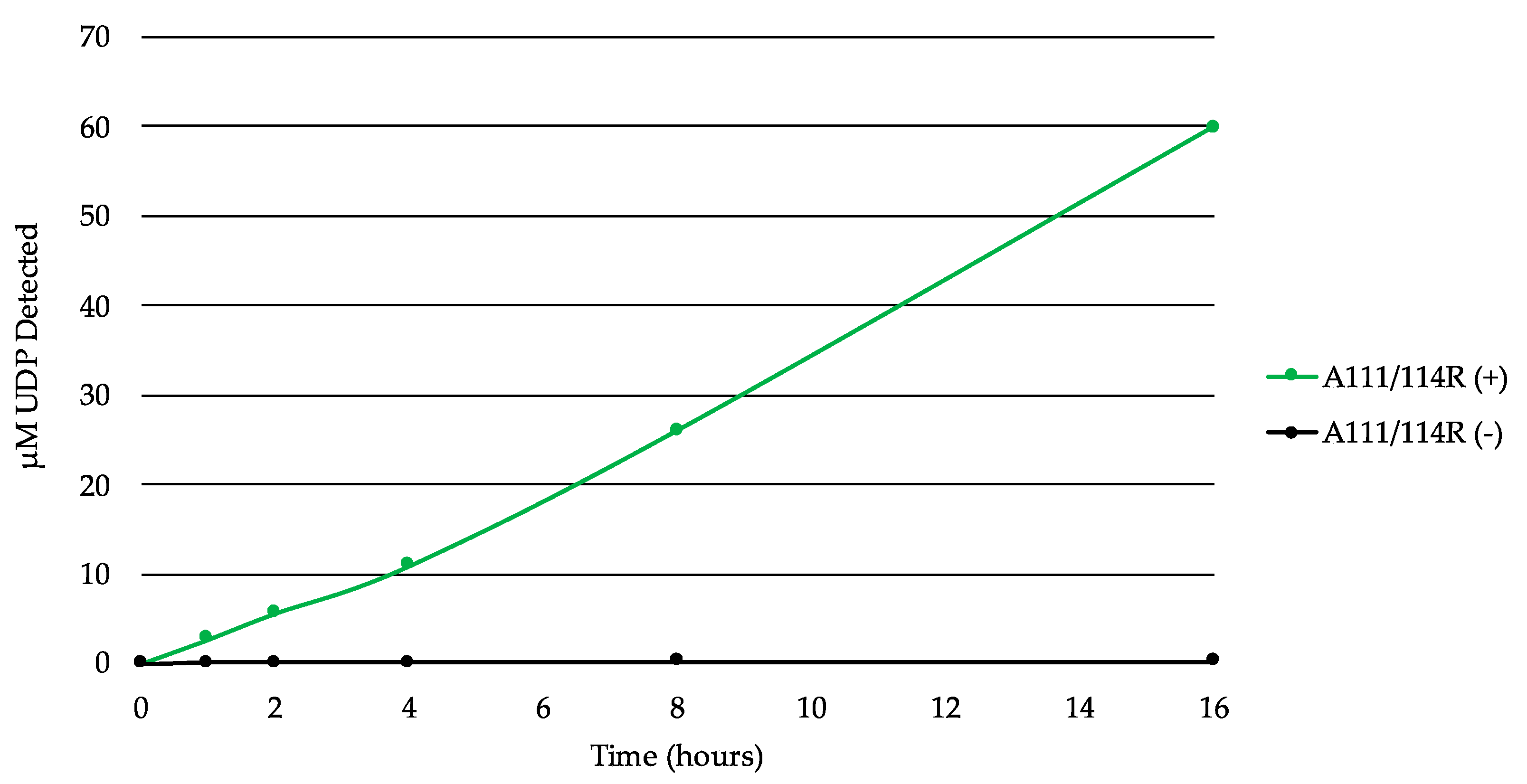
3.4. In Vitro Evidence of A111/114R Hydrolytic Activity
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bieberich, E. Synthesis, Processing, and Function of N-Glycans in N-Glycoproteins. In Glycobiology of the Nervous System; Springer: Berlin/Heidelberg, Germany, 2014; pp. 47–70. [Google Scholar]
- Vigerust, D.J.; Shepherd, V.L. Virus Glycosylation: Role in Virulence and Immune Interactions. Trends Microbiol. 2007, 15, 211–218. [Google Scholar] [CrossRef] [PubMed]
- Van Etten, J.L.; Gurnon, J.R.; Yanai-Balser, G.M.; Dunigan, D.D.; Graves, M.V. Chlorella Viruses Encode Most, If Not All, of the Machinery to Glycosylate Their Glycoproteins Independent of the Endoplasmic Reticulum and Golgi. Biochim. Biophys. Acta (BBA) Gen. Subj. 2010, 1800, 152–159. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Etten, J.L.; Agarkova, I.; Dunigan, D.D.; Tonetti, M.; De Castro, C.; Duncan, G.A. Chloroviruses Have a Sweet Tooth. Viruses 2017, 9, 88. [Google Scholar] [CrossRef] [PubMed]
- Van Etten, J.L.; Agarkova, I.V.; Dunigan, D.D. Chloroviruses. Viruses 2019, 12, 20. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Speciale, I.; Duncan, G.A.; Unione, L.; Agarkova, I.V.; Garozzo, D.; Jimenez-Barbero, J.; Lin, S.; Lowary, T.L.; Molinaro, A.; Noel, E.; et al. The N-Glycan Structures of the Antigenic Variants of Chlorovirus PBCV-1 Major Capsid Protein Help to Identify the Virus-Encoded Glycosyltransferases. J. Biol. Chem. 2019, 294, 5688–5699. [Google Scholar] [CrossRef] [PubMed]
- Nandhagopal, N.; Simpson, A.A.; Gurnon, J.R.; Yan, X.; Baker, T.S.; Graves, M.V.; Van Etten, J.L.; Rossmann, M.G. The Structure and Evolution of the Major Capsid Protein of a Large, Lipid-Containing DNA Virus. Proc. Natl. Acad. Sci. USA 2002, 99, 14758–14763. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Castro, C.; Klose, T.; Speciale, I.; Lanzetta, R.; Molinaro, A.; Van Etten, J.L.; Rossmann, M.G. Structure of the Chlorovirus PBCV-1 Major Capsid Glycoprotein Determined by Combining Crystallographic and Carbohydrate Molecular Modeling Approaches. Proc. Natl. Acad. Sci. USA 2018, 115, E44–E52. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Castro, C.; Molinaro, A.; Piacente, F.; Gurnon, J.R.; Sturiale, L.; Palmigiano, A.; Lanzetta, R.; Parrilli, M.; Garozzo, D.; Tonetti, M.G.; et al. Structure of N-Linked Oligosaccharides Attached to Chlorovirus PBCV-1 Major Capsid Protein Reveals Unusual Class of Complex N-Glycans. Proc. Natl. Acad. Sci. USA 2013, 110, 13956–13960. [Google Scholar] [CrossRef] [Green Version]
- Wieland, F.; Heitzer, R.; Schaefer, W. Asparaginylglucose: Novel Type of Carbohydrate Linkage. Proc. Natl. Acad. Sci. USA 1983, 80, 5470–5474. [Google Scholar] [CrossRef] [Green Version]
- Mengele, R.; Sumper, M. Drastic Differences in Glycosylation of Related S-Layer Glycoproteins from Moderate and Extreme Halophiles. J. Biol. Chem. 1992, 267, 8182–8185. [Google Scholar] [CrossRef]
- Schreiner, R.; Schnabel, E.; Wieland, F. Novel N-Glycosylation in Eukaryotes: Laminin Contains the Linkage Unit Beta-Glucosylasparagine. J. Cell Biol. 1994, 124, 1071–1081. [Google Scholar] [CrossRef] [Green Version]
- Gross, J.; Grass, S.; Davis, A.E.; Gilmore-Erdmann, P.; Townsend, R.R.; Geme, J.W.S. The Haemophilus Influenzae HMW1 Adhesin Is a Glycoprotein with an Unusual N-Linked Carbohydrate Modification. J. Biol. Chem. 2008, 283, 26010–26015. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Quispe, C.F.; Esmael, A.; Sonderman, O.; McQuinn, M.; Agarkova, I.; Battah, M.; Duncan, G.A.; Dunigan, D.D.; Smith, T.P.; De Castro, C. Characterization of a New Chlorovirus Type with Permissive and Non-Permissive Features on Phylogenetically Related Algal Strains. Virology 2017, 500, 103–113. [Google Scholar] [CrossRef] [PubMed]
- Speciale, I.; Agarkova, I.; Duncan, G.A.; Van Etten, J.L.; De Castro, C. Structure of the N-Glycans from the Chlorovirus Ne-Jv-1. Antonie Van Leeuwenhoek 2017, 110, 1391–1399. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Castro, C.; Speciale, I.; Duncan, G.; Dunigan, D.D.; Agarkova, I.; Lanzetta, R.; Sturiale, L.; Palmigiano, A.; Garozzo, D.; Molinaro, A.; et al. N-Linked Glycans of Chloroviruses Sharing a Core Architecture without Precedent. Angew. Chem. 2016, 55, 654–658. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Landstein, D.; Burbank, D.E.; Nietfeldt, J.W.; Van Etten, J.L. Large Deletions in Antigenic Variants of the Chlorella Virus PBCV-1. Virology 1995, 214, 413–420. [Google Scholar] [CrossRef] [PubMed]
- Kelley, L.A.; Mezulis, S.; Yates, C.M.; Wass, M.N.; Sternberg, M.J.E. The Phyre2 Web Portal for Protein Modeling, Prediction and Analysis. Nat. Protoc. 2015, 10, 845–858. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Söding, J.; Biegert, A.; Lupas, A.N. The Hhpred Interactive Server for Protein Homology Detection and Structure Prediction. Nucl. Acids Res. 2005, 33 (Suppl. 2), W244–W248. [Google Scholar]
- Altschul, S.F.; Madden, T.L.; Schäffer, A.A.; Zhang, J.; Zhang, Z.; Miller, W.; Lipman, D.J. Gapped Blast and Psi-Blast: A New Generation of Protein Database Search Programs. Nucl. Acids Res. 1997, 25, 3389–3402. [Google Scholar] [CrossRef] [Green Version]
- Culbertson, A.T.; Ehrlich, J.J.; Choe, J.-Y.; Honzatko, R.B.; Zabotina, O.A. Structure of Xyloglucan Xylosyltransferase 1 Reveals Simple Steric Rules That Define Biological Patterns of Xyloglucan Polymers. Proc. Natl. Acad. Sci. USA 2018, 115, 6064–6069. [Google Scholar] [CrossRef] [Green Version]
- Haltiwanger, R.S.; Yu, H.; Takeuchi, M.; LeBarron, J.; Kantharia, J.; London, E.; Bakker, H.; Li, H.; Takeuchi, H. Regulation of Notch Signaling by O-Glucosylation: Notch-Modifying Xylosyltransferase-Substrate Complexes Support an Sni-Like Retaining Mechanism. FASEB 2016, 30 (Suppl. 1), 624.3. [Google Scholar]
- Gibbons, B.J.; Roach, P.J.; Hurley, T.D. Crystal Structure of the Autocatalytic Initiator of Glycogen Biosynthesis, Glycogenin. J. Mol. Biol. 2002, 319, 463–477. [Google Scholar] [CrossRef]
- Persson, K.; Ly, H.D.; Dieckelmann, M.; Wakarchuk, W.W.; Withers, S.G.; Strynadka, N.C. Crystal Structure of the Retaining Galactosyltransferase Lgtc from Neisseria Meningitidis in Complex with Donor and Acceptor Sugar Analogs. Nat. Struct. Biol. 2001, 8, 166–175. [Google Scholar] [CrossRef] [PubMed]
- Campbell, J.A.; Davies, G.J.; Bulone, V.; Henrissat, B. A Classification of Nucleotide-Diphospho-Sugar Glycosyltransferases Based on Amino Acid Sequence Similarities. Biochem. J. 1997, 326, 929–939. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Y.-L.; Jin, H.; Yang, H.-B.; Zhao, R.-L.; Wang, S.; Chen, Y.; Zhou, C.-Z. Defining the Enzymatic Pathway for Polymorphic O-Glycosylation of the Pneumococcal Serine-Rich Repeat Protein Psrp. J. Biol. Chem. 2017, 292, 6213–6224. [Google Scholar] [CrossRef] [Green Version]
- Clarke, B.R.; Ovchinnikova, O.G.; Sweeney, R.P.; Kamski-Hennekam, E.R.; Gitalis, R.; Mallette, E.; Kelly, S.D.; Lowary, T.L.; Kimber, M.S.; Whitfield, C. A Bifunctional O-Antigen Polymerase Structure Reveals a New Glycosyltransferase Family. Nat. Chem. Biol. 2020, 16, 450–457. [Google Scholar] [CrossRef]
- Ninomiya, T.; Sugiura, N.; Tawada, A.; Sugimoto, K.; Watanabe, H.; Kimata, K. Molecular Cloning and Characterization of Chondroitin Polymerase from Escherichia Coli Strain K4. J. Biol. Chem. 2002, 277, 21567–21575. [Google Scholar] [CrossRef] [Green Version]
- Sun, H.-Y.; Lin, S.-W.; Ko, T.-P.; Pan, J.-F.; Liu, C.-L.; Lin, C.-N.; Wang, A.H.-J.; Lin, C.-H. Structure and Mechanism of Helicobacter Pylori Fucosyltransferase a Basis for Lipopolysaccharide Variation and Inhibitor Design. J. Biol. Chem. 2007, 282, 9973–9982. [Google Scholar] [CrossRef] [Green Version]
- Speciale, I.; Laugieri, M.E.; Noel, E.; Lin, S.; Lowary, T.L.; Molinaro, A.; Duncan, G.A.; Agarkova, I.V.; Garozzo, D.; Tonetti, M.G. Chlorovirus PBCV-1 Protein A064R Has Three of the Transferase Activities Necessary to Synthesize Its Capsid Protein N-Linked Glycans. Proc. Natl. Acad. Sci. USA 2020, 117, 28735–28742. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| A111/114R Residues | H-Bond Distance of Nucleotide-Sugar (Å) | |
|---|---|---|
| Domain 1 | Tyr-83 | <4 |
| Asp-85 | 2.6, 3.0, 3.6 | |
| Asn-149 | <4 | |
| Ala-150 | <4 | |
| Asp-193 | 2.4, 2.5 | |
| Gln-194 | <4 | |
| Domain 2 | Pro-265 | 2.8, 3.3 |
| Asp-294 | <4 | |
| Asn-323 | <4 | |
| Gly-328 | <4 | |
| Asp-351 | 3.2, 3.3 | |
| Asp-353 | <4 | |
| Asp-447 | <4 | |
| Domain 3 | Leu-617 | <4 |
| Val-684 | <4 | |
| Arg-693 | <4 | |
| Gly-711 | <4 | |
| Asn-749 | 1.6, 3.1, 3.3 | |
| Tyr-755 | 1.9, 3.4 | |
| Ser-757 | <4 | |
| Glu-758 | 2.2 | |
| Lys-759 | 2.5 | |
| Asp-762 | <4 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Noel, E.; Notaro, A.; Speciale, I.; Duncan, G.A.; De Castro, C.; Van Etten, J.L. Chlorovirus PBCV-1 Multidomain Protein A111/114R Has Three Glycosyltransferase Functions Involved in the Synthesis of Atypical N-Glycans. Viruses 2021, 13, 87. https://doi.org/10.3390/v13010087
Noel E, Notaro A, Speciale I, Duncan GA, De Castro C, Van Etten JL. Chlorovirus PBCV-1 Multidomain Protein A111/114R Has Three Glycosyltransferase Functions Involved in the Synthesis of Atypical N-Glycans. Viruses. 2021; 13(1):87. https://doi.org/10.3390/v13010087
Chicago/Turabian StyleNoel, Eric, Anna Notaro, Immacolata Speciale, Garry A. Duncan, Cristina De Castro, and James L. Van Etten. 2021. "Chlorovirus PBCV-1 Multidomain Protein A111/114R Has Three Glycosyltransferase Functions Involved in the Synthesis of Atypical N-Glycans" Viruses 13, no. 1: 87. https://doi.org/10.3390/v13010087