Genetic and Phenotypic Characterization of a Rabies Virus Strain Isolated from a Dog in Tokyo, Japan in the 1940s

 , , ,
, , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cells and Virus Strains
2.2. Complete Genome Sequencing of Koma Strain
2.3. Genetic and Phylogenetic Analyses
2.4. Growth of Each Virus Strain in NA Cells
2.5. Focus Formation by Each Virus Strain in NA Cells
2.6. Pathogenicity of Each Virus Strain In Vivo
2.7. Examination of Incubation Period in Mice
2.8. Immunohistochemical Analysis
2.9. Statistical Analysis
3. Results
3.1. Genome Organization of Koma Strain
3.2. Phylogenetic Relationships between Koma Strain and Other RABV Strains
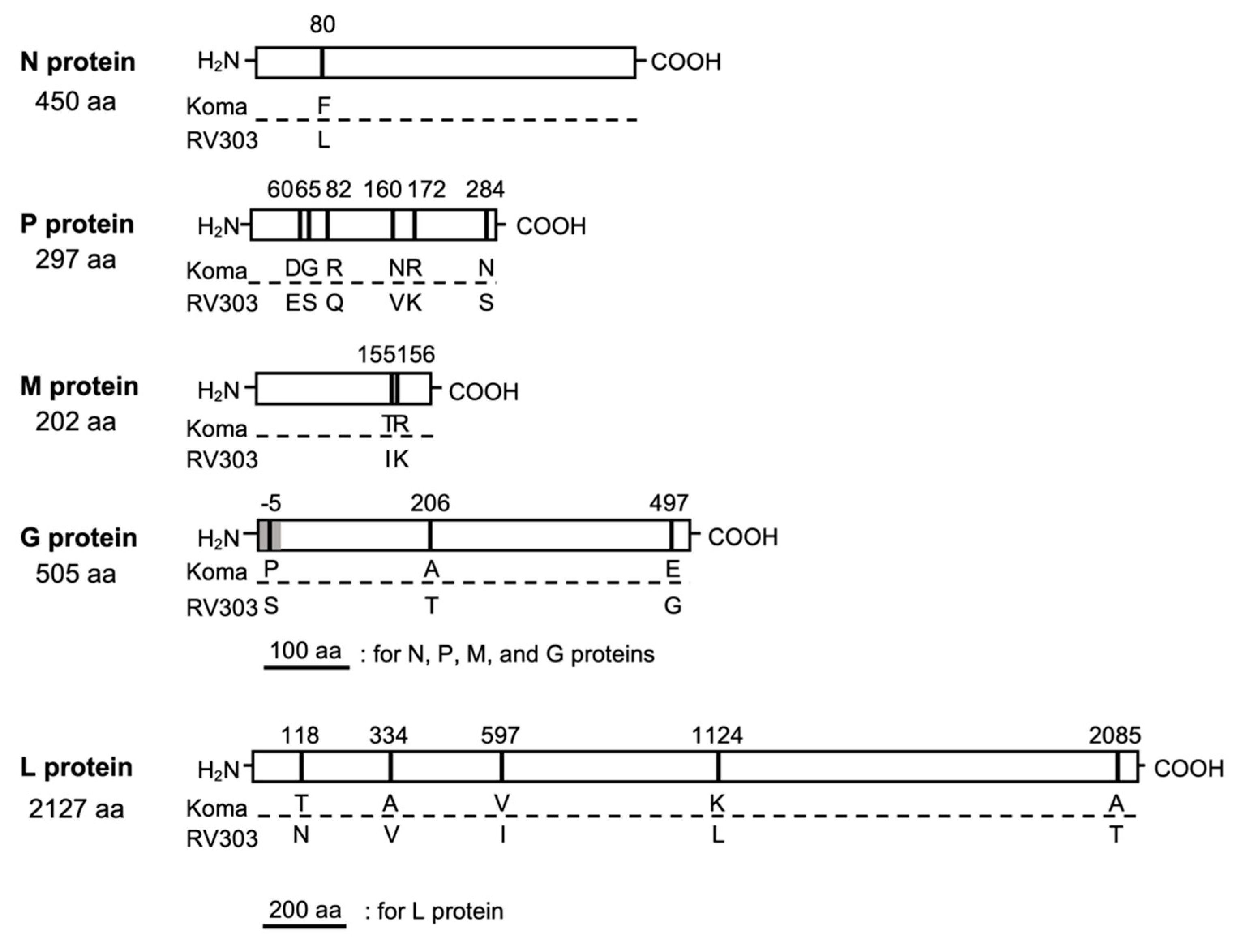
3.3. Genetic Properties of Koma Strain
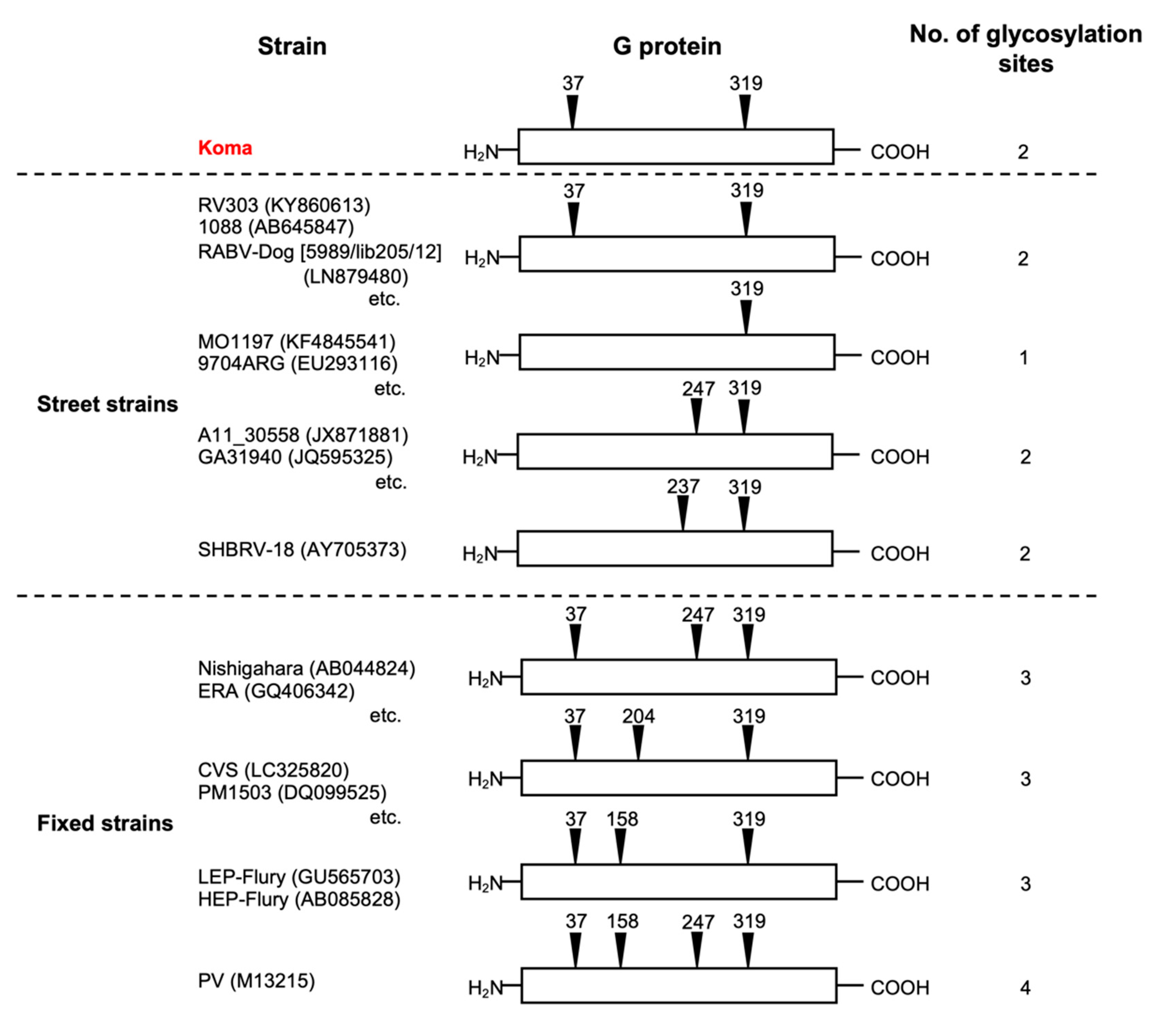
3.4. Number and Position of Potential N-Glycosylation Sites on the Koma G Protein
3.5. Growth of Koma Strain in Neuroblastoma NA Cells
3.6. Cell-to-Cell Spread of Koma Strain in NA Cells
3.7. Neurovirulence and Neuroinvasiveness of Koma Strain in Mice
3.8. Incubation Period in Koma-Infected Mice
3.9. Distribution of Virus in the Brains of Mice Infected with Koma Strain via the i.m. Route
4. Discussion
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
Appendix A

References
- Jackson, A.C.; Fu, Z.F. Pathogenesis. In Rabies; Elsevier Inc.: Amsterdam, The Netherlands, 2013; pp. 299–349. ISBN 9780123965479. [Google Scholar]
- WHO/Department of Control of Neglected Tropical Diseases. Human Rabies Transmitted by Dogs: Current Status of Global Data, 2015. Wkly. Epidemiol. Rec. 2016, 2, 13–20. [Google Scholar]
- Hampson, K.; Coudeville, L.; Lembo, T.; Sambo, M.; Kieffer, A.; Attlan, M.; Barrat, J.; Blanton, J.D.; Briggs, D.J.; Cleaveland, S.; et al. Estimating the Global Burden of Endemic Canine Rabies. PLoS Negl. Trop. Dis. 2015, 9. [Google Scholar] [CrossRef] [Green Version]
- WHO/Department of Control of Neglected Tropical Diseases. Rabies Vaccines: WHO Position Paper–April 2018. Wkly. Epidemiol. Rec. 2018, 16, 201–220. [Google Scholar]
- Jackson, A.C.; Fu, Z.F. Rabies virus. In Rabies; Elsevier Inc.: Amsterdam, The Netherlands, 2013; pp. 17–60. ISBN 9780123965479. [Google Scholar]
- Vidy, A.; Chelbi-Alix, M.; Blondel, D. Rabies Virus P Protein Interacts with STAT1 and Inhibits Interferon Signal Transduction Pathways. J. Virol. 2005, 79, 14411–14420. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vidy, A.; El Bougrini, J.; Chelbi-Alix, M.K.; Blondel, D. The Nucleocytoplasmic Rabies Virus P Protein Counteracts Interferon Signaling by Inhibiting Both Nuclear Accumulation and DNA Binding of STAT1. J. Virol. 2007, 81, 4255–4263. [Google Scholar] [CrossRef] [Green Version]
- Masatani, T.; Ito, N.; Shimizu, K.; Ito, Y.; Nakagawa, K.; Abe, M.; Yamaoka, S.; Sugiyama, M. Amino Acids at Positions 273 and 394 in Rabies Virus Nucleoprotein Are Important for Both Evasion of Host RIG-I-mediated Antiviral Response and Pathogenicity. Virus Res. 2011, 155, 168–174. [Google Scholar] [CrossRef]
- Brzózka, K.; Finke, S.; Conzelmann, K.-K. Identification of the Rabies Virus Alpha/Beta Interferon Antagonist: Phosphoprotein P Interferes with Phosphorylation of Interferon Regulatory Factor 3. J. Virol. 2005, 79, 7673–7681. [Google Scholar] [CrossRef] [Green Version]
- Masatani, T.; Ito, N.; Shimizu, K.; Ito, Y.; Nakagawa, K.; Sawaki, Y.; Koyama, H.; Sugiyama, M. Rabies Virus Nucleoprotein Functions to Evade Activation of the RIG-I-Mediated Antiviral Response. J. Virol. 2010, 84, 4002–4012. [Google Scholar] [CrossRef] [Green Version]
- Sonthonnax, F.; Besson, B.; Bonnaud, E.; Jouvion, G.; Merino, D.; Larrous, F.; Bourhy, H. Lyssavirus Matrix Protein Cooperates with Phosphoprotein to Modulate the Jak-Stat Pathway. Sci. Rep. 2019, 9. [Google Scholar] [CrossRef]
- Yamaoka, S.; Ito, N.; Ohka, S.; Kaneda, S.; Nakamura, H.; Agari, T.; Masatani, T.; Nakagawa, K.; Okada, K.; Okadera, K.; et al. Involvement of the Rabies Virus Phosphoprotein Gene in Neuroinvasiveness. J. Virol. 2013, 87, 12327–12338. [Google Scholar] [CrossRef] [Green Version]
- Okada, K.; Ito, N.; Yamaoka, S.; Masatani, T.; Ebihara, H.; Goto, H.; Nakagawa, K.; Mitake, H.; Okadera, K.; Sugiyama, M. Roles of the Rabies Virus Phosphoprotein Isoforms in Pathogenesis. J. Virol. 2016, 90, 8226–8237. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Prehaud, C.; Coulon, P.; LaFay, F.; Thiers, C.; Flamand, A. Antigenic Site II of the Rabies Virus Glycoprotein: Structure and Role in Viral Virulence. J. Virol. 1988, 62, 1–7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dietzschold, B.; Wunner, W.H.; Wiktor, T.J.; Lopes, A.D.; Lafon, M.; Smith, C.L.; Koprowski, H. Characterization of an Antigenic Determinant of the Glycoprotein That Correlates with Pathogenicity of Rabies Virus. Proc. Natl. Acad. Sci. USA 1983, 80, 70–74. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Faber, M.; Faber, M.-L.; Papaneri, A.; Bette, M.; Weihe, E.; Dietzschold, B.; Schnell, M.J. A Single Amino Acid Change in Rabies Virus Glycoprotein Increases Virus Spread and Enhances Virus Pathogenicity. J. Virol. 2005, 79, 14141–14148. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takayama-Ito, M.; Inoue, K.; Shoji, Y.; Inoue, S.; Iijima, T.; Sakai, T.; Kurane, I.; Morimoto, K. A Highly Attenuated Rabies Virus HEP-Flury Strain Reverts to Virulent by Single Amino Acid Substitution to Arginine at Position 333 in Glycoprotein. Virus Res. 2006, 119, 208–215. [Google Scholar] [CrossRef] [PubMed]
- Seif, I.; Coulon, P.; Rollin, P.E.; Flamand, A. Rabies Virulence: Effect on Pathogenicity and Sequence Characterization of Rabies Virus Mutations Affecting Antigenic Site III of the Glycoprotein. J. Virol. 1985, 53, 926–934. [Google Scholar] [CrossRef] [Green Version]
- Shimizu, K.; Ito, N.; Mita, T.; Yamada, K.; Hosokawa-Muto, J.; Sugiyama, M.; Minamoto, N. Involvement of Nucleoprotein, Phosphoprotein, and Matrix Protein Genes of Rabies Virus in Virulence for Adult Mice. Virus Res. 2007, 123, 154–160. [Google Scholar] [CrossRef]
- Takayama-Ito, M.; Ito, N.; Yamada, K.; Sugiyama, M.; Minamoto, N. Multiple Amino Acids in the Glycoprotein of Rabies Virus Are Responsible for Pathogenicity in Adult Mice. Virus Res. 2006, 115, 169–175. [Google Scholar] [CrossRef]
- Yamada, K.; Ito, N.; Takayama-Ito, M.; Sugiyama, M.; Minamoto, N. Multigenic Relation to the Attenuation of Rabies Virus. Microbiol. Immunol. 2006, 50, 25–32. [Google Scholar] [CrossRef] [Green Version]
- Schnell, M.J.; Tan, G.S.; Dietzschold, B. The Application of Reverse Genetics Technology in the Study of Rabies Virus (RV) Pathogenesis and for the Development of Novel RV Vaccines. J. Neurovirol. 2005, 11, 76–81. [Google Scholar] [CrossRef]
- Yamada, K.; Park, C.H.; Noguchi, K.; Kojima, D.; Kubo, T.; Komiya, N.; Matsumoto, T.; Mitui, M.T.; Ahmed, K.; Morimoto, K.; et al. Serial Passage of a Street Rabies Virus in Mouse Neuroblastoma Cells Resulted in Attenuation: Potential Role of the Additional N-Glycosylation of a Viral Glycoprotein in the Reduced Pathogenicity of Street Rabies Virus. Virus Res. 2012, 165, 34–45. [Google Scholar] [CrossRef] [PubMed]
- Potratz, M.; Zaeck, L.; Christen, M.; te Kamp, V.; Klein, A.; Nolden, T.; Freuling, C.; Müller, T.; Finke, S. Astrocyte Infection during Rabies Encephalitis Depends on the Virus Strain and Infection Route as Demonstrated by Novel Quantitative 3D Analysis of Cell Tropism. Cells 2020, 9, 412. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nolden, T.; Pfaff, F.; Nemitz, S.; Freuling, C.M.; Höper, D.; Müller, T.; Finke, S. Reverse Genetics in High Throughput: Rapid Generation of Complete Negative Strand RNA Virus cDNA Clones and Recombinant Viruses Thereof. Sci. Rep. 2016, 6, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Isomura, M.; Yamada, K.; Noguchi, K.; Nishizono, A. Near-Infrared Fluorescent Protein iRFP720 Is Optimal for in vivo Fluorescence Imaging of Rabies Virus Infection. J. Gen. Virol. 2017, 98, 2689–2698. [Google Scholar] [CrossRef] [PubMed]
- Kimitsuki, K.; Yamada, K.; Shiwa, N.; Inoue, S.; Nishizono, A.; Park, C.H. Pathological Lesions in the Central Nervous System and Peripheral Tissues of ddY Mice with Street Rabies Virus (1088 strain). J. Vet. Med. Sci. 2017, 79, 970–978. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Virojanapirom, P.; Khawplod, P.; Sawangvaree, A.; Wacharapluesadee, S.; Hemachudha, T.; Yamada, K.; Morimoto, K.; Nishizono, A. Molecular Analysis of the Mutational Effects of Thai Street Rabies Virus with Increased Virulence in Mice after Passages in the BHK Cell Line. Arch. Virol. 2012, 157, 2201–2205. [Google Scholar] [CrossRef] [PubMed]
- Tian, Q.; Wang, Y.; Zhang, Q.; Luo, J.; Jiang, H.; Zhang, B.; Mei, M.; Wu, F.; Wu, Y.; Peng, J.; et al. Phosphoprotein Gene Contributes to the Enhanced Apoptosis Induced by Wild-Type Rabies Virus GD-SH-01 in vitro. Front. Microbiol. 2017, 8. [Google Scholar] [CrossRef] [Green Version]
- Faber, M.; Pulmanausahakul, R.; Nagao, K.; Prosniak, M.; Rice, A.B.; Koprowski, H.; Schnell, M.J.; Dietzschold, B. Identification of Viral Genomic Elements Responsible for Rabies Virus Neuroinvasiveness. Proc. Natl. Acad. Sci. USA 2004, 101, 16328–16332. [Google Scholar] [CrossRef] [Green Version]
- Troupin, C.; Dacheux, L.; Tanguy, M.; Sabeta, C.; Blanc, H.; Bouchier, C.; Vignuzzi, M.; Duchene, S.; Holmes, E.C.; Bourhy, H. Large-Scale Phylogenomic Analysis Reveals the Complex Evolutionary History of Rabies Virus in Multiple Carnivore Hosts. PLoS Pathog. 2016, 12, 1–20. [Google Scholar] [CrossRef]
- Miyamoto, K. Electron Microscopic Studies of the Negri Body. Uirusu 1966, 16, 13–31. [Google Scholar] [CrossRef] [Green Version]
- Arai, Y. Phylogenetic Analysis of Two Rabies Viruses, Takamen and Komatsugawa Strains Isolated in Japan in the 1940’s. J. Jpn. Assoc. Infect. Dis. 2004, 78, 815–822. [Google Scholar] [CrossRef]
- Ito, N.; Sugiyama, M.; Oraveerakul, K.; Piyaviriyakul, P.; Lumlertdacha, B.; Arai, Y.T.; Tamura, Y.; Mori, Y.; Minamoto, N. Molecular Epidemiology of Rabies in Thailand. Microbiol. Immunol. 1999, 43, 551–559. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arai, Y.T.; Yamada, K.; Kameoka, Y.; Horimoto, T.; Yamamoto, K.; Yabe, S.; Nakayama, M.; Tashiro, M. Nucleoprotein Gene Analysis of Fixed and Street Rabies Virus Variants Using RT-PCR. Arch. Virol. 1997, 142, 1787–1796. [Google Scholar] [CrossRef] [PubMed]
- Luo, T.R.; Minamoto, N.; Hishida, M.; Yamamoto, K.; Fujise, T.; Hiraga, S.; Ito, N.; Sugiyama, M.; Kinjo, T. Antigenic and Functional Analyses of Glycoprotein of Rabies Virus Using Monoclonal Antibodies. Microbiol. Immunol. 1998, 42, 187–193. [Google Scholar] [CrossRef]
- Hatakeyama, K.; Sadamasu, K.; Kai, A. Phylogenetic Analysis of Rabies Viruses Isolated from Animals in Tokyo in the 1950s. J. Jpn. Assoc. Infect. Dis. 2011, 85, 238–243. [Google Scholar] [CrossRef]
- Kuzmin, I.V.; Botvinkin, A.D.; McElhinney, L.M.; Smith, J.S.; Orciari, L.A.; Hughes, G.J.; Fooks, A.R.; Rupprecht, C.E. Molecular Epidemiology of Terrestrial Rabies in the Former Soviet Union. J. Wildl. Dis. 2004, 40, 617–631. [Google Scholar] [CrossRef] [Green Version]
- McMorris, F.A.; Ruddle, F.H. Expression of Neuronal Phenotypes in Neuroblastoma Cell Hybrids. Dev. Biol. 1974, 39, 226–246. [Google Scholar] [CrossRef]
- Ito, N.; Takayama-Ito, M.; Yamada, K.; Hosokawa, J.; Sugiyama, M.; Minamoto, N. Improved Recovery of Rabies Virus from Cloned cDNA Using a Vaccinia Virus-Free Reverse Genetics System. Microbiol. Immunol. 2003, 47, 613–617. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular Evolutionary Genetics Analysis across Computing Platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef]
- Nakagawa, K.; Kobayashi, Y.; Ito, N.; Suzuki, Y.; Okada, K.; Makino, M.; Goto, H.; Takahashi, T.; Sugiyama, M. Molecular Function Analysis of Rabies Virus RNA Polymerase L Protein by Using an L Gene-Deficient Virus. J. Virol. 2017, 91, 1–16. [Google Scholar] [CrossRef] [Green Version]
- Reed, L.J.; Muench, H. A Simple Method of Estimating Fifty Percent Endpoints. Am. J. Epidemiol. 1938, 27, 493–497. [Google Scholar] [CrossRef]
- Mita, T.; Shimizu, K.; Ito, N.; Yamada, K.; Ito, Y.; Sugiyama, M.; Minamoto, N. Amino Acid at Position 95 of the Matrix Protein Is a Cytopathic Determinant of Rabies Virus. Virus Res. 2008, 137, 33–39. [Google Scholar] [CrossRef] [PubMed]
- Ito, N.; Moseley, G.W.; Blondel, D.; Shimizu, K.; Rowe, C.L.; Ito, Y.; Masatani, T.; Nakagawa, K.; Jans, D.A.; Sugiyama, M. Role of Interferon Antagonist Activity of Rabies Virus Phosphoprotein in Viral Pathogenicity. J. Virol. 2010, 84, 6699–6710. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wirblich, C.; Tan, G.S.; Papaneri, A.; Godlewski, P.J.; Orenstein, J.M.; Harty, R.N.; Schnell, M.J. PPEY Motif within the Rabies Virus (RV) Matrix Protein Is Essential for Efficient Virion Release and RV Pathogenicity. J. Virol. 2008, 82, 9730–9738. [Google Scholar] [CrossRef] [Green Version]
- Hamamoto, N.; Uda, A.; Tobiume, M.; Park, C.H.; Noguchi, A.; Kaku, Y.; Okutani, A.; Morikawa, S.; Inoue, S. Association between RABV G Proteins Transported from the Perinuclear Space to the Cell Surface Membrane and N-Glycosylation of the Sequon Asn204. Jpn. J. Infect. Dis. 2015, 68, 387–393. [Google Scholar] [CrossRef] [Green Version]
- Gautret, P.; Carrara, P.; Parola, P. Long Incubation in Imported Human Rabies. Ann. Neurol. 2014, 75, 324–325. [Google Scholar] [CrossRef]
- Charlton, K.M.; Nadin-Davis, S.; Casey, G.A.; Wandeler, A.I. The Long Incubation Period in Rabies: Delayed Progression of Infection in Muscle at the Site of Exposure. Acta Neuropathol. 1997, 94, 73–77. [Google Scholar] [CrossRef]
- Smith, J.S.; Fishbein, D.B.; Rupprecht, C.E.; Clark, K. Unexplained Rabies in Three Immigrants in the United States a Virologic Investigation. N. Engl. J. Med. 1991, 324, 205–211. [Google Scholar] [CrossRef]
- Brzózka, K.; Finke, S.; Conzelmann, K.-K. Inhibition of Interferon Signaling by Rabies Virus Phosphoprotein P: Activation-Dependent Binding of STAT1 and STAT2. J. Virol. 2006, 80, 2675–2683. [Google Scholar] [CrossRef] [Green Version]
- Rieder, M.; Brzózka, K.; Pfaller, C.K.; Cox, J.H.; Stitz, L.; Conzelmann, K.-K. Genetic Dissection of Interferon-Antagonistic Functions of Rabies Virus Phosphoprotein: Inhibition of Interferon Regulatory Factor 3 Activation Is Important for Pathogenicity. J. Virol. 2011, 85, 842–852. [Google Scholar] [CrossRef] [Green Version]
- Lépine, P. On the Evolution of Fixed Strains of Rabies Virus. J. Hyg. 1938, 38, 180–184. [Google Scholar] [CrossRef] [PubMed]
- Yamada, K.; Noguchi, K.; Nishizono, A. Efficient N-Glycosylation at Position 37, but Not at Position 146, in the Street Rabies Virus Glycoprotein Reduces Pathogenicity. Virus Res. 2014, 179, 169–176. [Google Scholar] [CrossRef] [PubMed]
- Yamada, K.; Noguchi, K.; Nishizono, A. Characterization of Street Rabies Virus Variants with an Additional N-Glycan at Position 247 in the Glycoprotein. Arch. Virol. 2014, 159, 207–216. [Google Scholar] [CrossRef] [PubMed]
- Baer, G.M.; Cleary, W.F. Model in Mice for the Pathogenesis and Treatment of Rabies. J. Infect. Dis. 1972, 125, 520–527. [Google Scholar] [CrossRef]
- LeDoux, J. A Few Degrees of Separation. In The emotional Brain: The Mysterious Underpinnings of Emotional Life; Simon and Schuster: New York, NY, USA, 1998; pp. 138–178. ISBN 1439126380. [Google Scholar]
- Jackson, A.C.; Fu, Z.F. Human disease. In Rabies; Elsevier Inc.: Amsterdam, The Netherlands, 2013; pp. 269–298. ISBN 9780123965479. [Google Scholar]
- Jackson, A.C.; Fu, Z.F. Rabies in Terrestrial Animals. In Rabies; Elsevier Inc.: Amsterdam, The Netherlands, 2013; pp. 179–214. ISBN 9780123965479. [Google Scholar]
- Zaeck, L.; Potratz, M.; Freuling, C.M.; Müller, T.; Finke, S. High-Resolution 3D Imaging of Rabies Virus Infection in Solvent-Cleared Brain Tissue. J. Vis. Exp. 2019, 146. [Google Scholar] [CrossRef] [PubMed]
- Kuzmin, I.V.; Hughes, G.J.; Botvinkin, A.D.; Gribencha, S.G.; Rupprecht, C.E. Arctic and Arctic-like Rabies Viruses: Distribution, Phylogeny and Evolutionary History. Epidemiol. Infect. 2008, 136, 509–519. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Total | N Gene | P Gene | M Gene | G Gene | L Gene | |
|---|---|---|---|---|---|---|
| Nucleotide | 98.2% | 98.5% | 97.2% | 97.7% | 98.1% | 98.3% |
| Amino acid | 99.5% 1 | 99.8% | 98.0% | 99.0% | 99.4% | 99.8% |
| Dose | Koma | 1088 | RABV-Dog | Nishigahara | CVS | ERA |
|---|---|---|---|---|---|---|
| 1.0 × 103 FFU | NT | NT | NT | 100% (5/5) 1 | 100% (5/5) | 60% (3/5) |
| 1.0 × 102 FFU | 100% (10/10) | 100% (10/10) | 70% (7/10) | 80% (4/5) | 60% (3/5) | 100% (5/5) |
| 1.0 × 101 FFU | 80% (8/10) | 50% (5/10) | 50% (5/10) | 40% (2/5) | 20% (1/5) | 20% (1/5) |
| 1.0 × 100 FFU | 0% (0/10) | 0% (0/10) | 10% (1/10) | 0% (0/5) | 0% (0/5) | 0% (0/5) |
| LD50 (FFU) | 4.2 | 10.0 | 15.3 | 20.8 | 48.1 | ND |
| Dose | Koma | 1088 | RABV-Dog | Nishigahara | CVS | ERA |
|---|---|---|---|---|---|---|
| 1.0 × 106 FFU | NT | NT | NT | 100% (5/5) 2 | 100% (5/5) | 40% (2/5) |
| 1.0 × 105 FFU | NT | NT | NT | 40% (2/5) | 20% (1/5) | 20% (1/5) |
| 1.0 × 104 FFU | NT | 100% (10/10) | 60% (6/10) | 0% (0/5) | 0% (0/5) | 0% (0/5) |
| 1.0 × 103 FFU | 80% (8/10) | 20% (2/10) | 30% (3/10) | NT | NT | NT |
| 1.0 × 102 FFU | 20% (2/10) | 0% (0/10) | 10% (1/10) | NT | NT | NT |
| LD50 (FFU) | 3.2 × 102 | 2.4 × 103 | 3.3 × 103 | 1.5 × 105 | 2.4 × 105 | >1.0 × 106 |
| Fixed Viruses (FVs) vs. Street Viruses (SVs) vs. Koma Strain | |
|---|---|
| Growth in neuroblastoma cells | FVs > SVs = Koma |
| Cell-to-cell spread in neuroblastoma cells | FVs 1 > SVs = Koma |
| Neurovirulence in mice (i.c. route) | FVs = SVs = Koma |
| Neuroinvasiveness in mice (i.m. route) | FVs < SVs = Koma |
| Incubation period in mice (i.m. route) | FVs (Constant) < SVs = Koma (Inconstant) |
| Virus spread in the mouse brain | FVs > SVs 2 = Koma 2 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Takahashi, T.; Inukai, M.; Sasaki, M.; Potratz, M.; Jarusombuti, S.; Fujii, Y.; Nishiyama, S.; Finke, S.; Yamada, K.; Sakai, H.; et al. Genetic and Phenotypic Characterization of a Rabies Virus Strain Isolated from a Dog in Tokyo, Japan in the 1940s. Viruses 2020, 12, 914. https://doi.org/10.3390/v12090914
Takahashi T, Inukai M, Sasaki M, Potratz M, Jarusombuti S, Fujii Y, Nishiyama S, Finke S, Yamada K, Sakai H, et al. Genetic and Phenotypic Characterization of a Rabies Virus Strain Isolated from a Dog in Tokyo, Japan in the 1940s. Viruses. 2020; 12(9):914. https://doi.org/10.3390/v12090914
Chicago/Turabian StyleTakahashi, Tatsuki, Maho Inukai, Michihito Sasaki, Madlin Potratz, Supasiri Jarusombuti, Yuji Fujii, Shoko Nishiyama, Stefan Finke, Kentaro Yamada, Hiroki Sakai, and et al. 2020. "Genetic and Phenotypic Characterization of a Rabies Virus Strain Isolated from a Dog in Tokyo, Japan in the 1940s" Viruses 12, no. 9: 914. https://doi.org/10.3390/v12090914