Prolonged Detection of Bovine Viral Diarrhoea Virus Infection in the Semen of Bulls

 , , , and
, , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Specimens
2.2. Serology
2.3. Virus and Nucleic Acid Detection
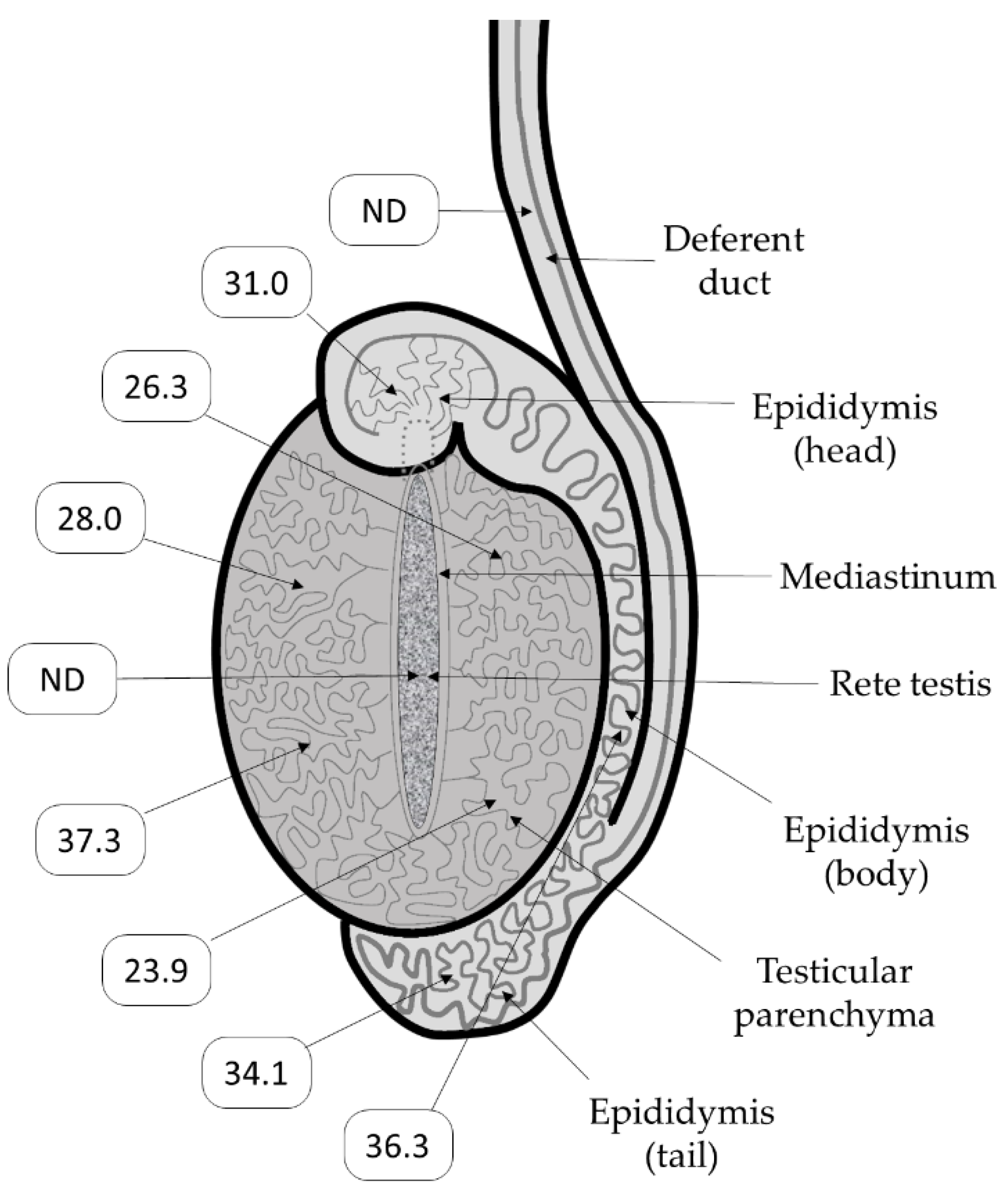
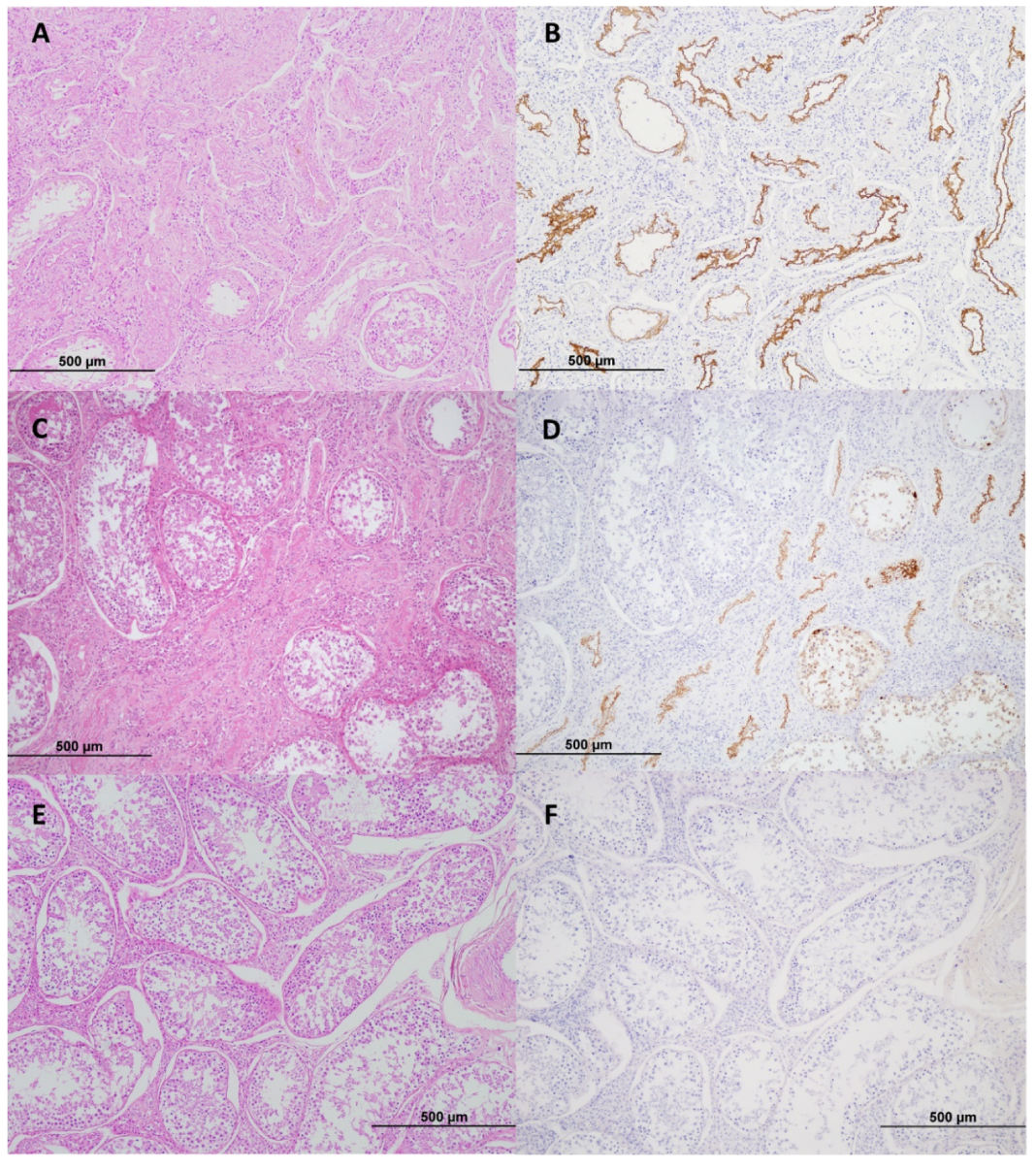
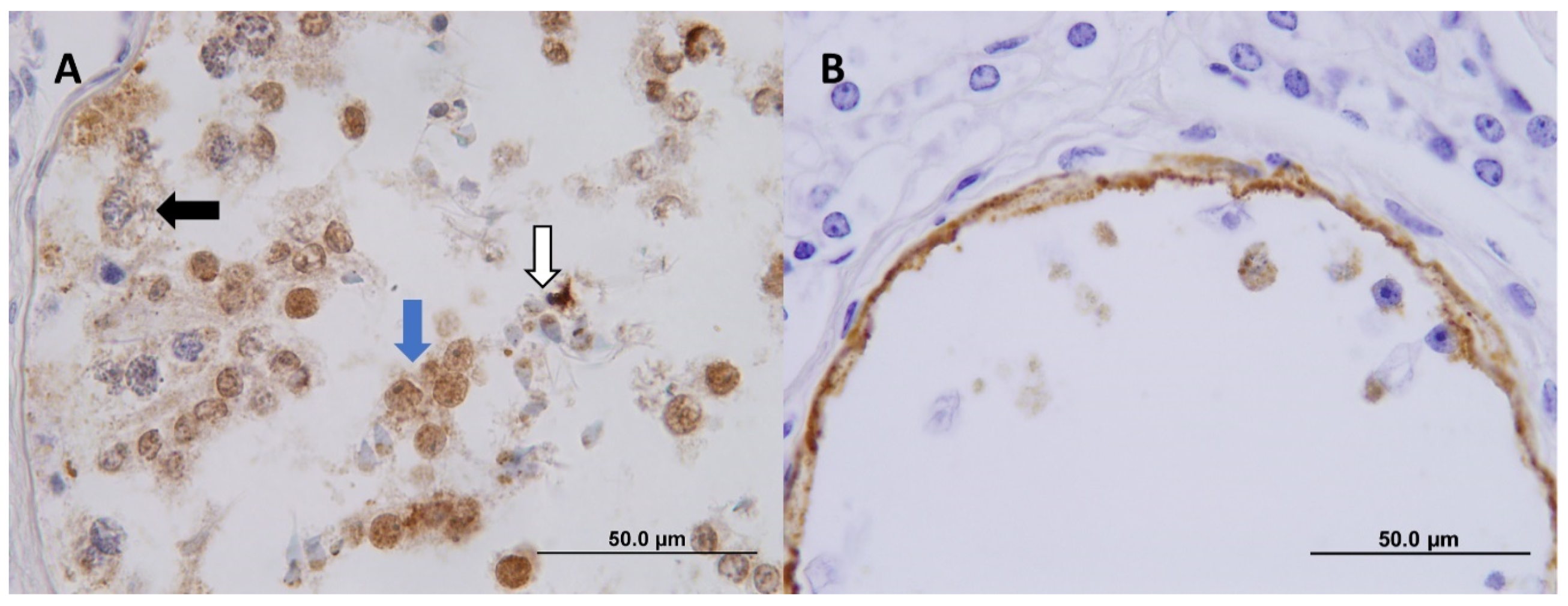
2.4. Immunohistochemistry (IHC)
3. Results
4. Discussion
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
| Sample Date | Sample Type | Age (Months) | qRT-PCR (Ct) | Virus Isolation | Antigen ELISA | VNT (Titre) |
|---|---|---|---|---|---|---|
| 28/08/2012 | Serum | 21 | Negative | 512 | ||
| 22/10/2012 | Serum | 23 | Negative | ≥512 | ||
| 12/11/2012 | Extended semen | 23 | Positive (35.4) | Negative | ||
| 23/11/2012 | Extended semen | 24 | Positive (35.4) | Negative | ||
| 26/11/2012 | Extended semen | 24 | Positive (35.8) | Negative | ||
| 28/11/2012 | Extended semen | 24 | Positive (32.8) | Negative | ||
| 30/11/2012 | Extended semen | 24 | Positive (35.5) | Negative | ||
| 6/12/2012 | Serum | 24 | ≥512 |
| Sample Date | Sample Type | Age (Months) | qRT-PCR (Ct) | Virus Isolation | Antigen ELISA | AGID | VNT (Titre) |
|---|---|---|---|---|---|---|---|
| 23/01/2012 | Serum | 10 | Negative | 3 | |||
| 13/03/2012 | Serum | 12 | Negative | 2 | |||
| 17/04/2012 | Extended semen | 13 | Negative a | Negative | |||
| 17/04/2012 | Extended semen | 15 | Negative a | ||||
| 19/06/2012 | Extended semen | 15 | Negative a | ||||
| 29/06/2012 | Extended semen | 16 | Negative a | ||||
| 20/07/2012 | Extended semen | 17 | Negative a | ||||
| 8/08/2012 | Extended semen | 17 | Negative a | ||||
| 20/08/2012 | Extended semen | 17 | Negative a | ||||
| 24/08/2012 | Extended semen | 17 | Negative a | ||||
| 31/08/2012 | Extended semen | 18 | Negative a | ||||
| 4/09/2012 | Extended semen | 18 | Negative a | ||||
| 7/09/2012 | Extended semen | 18 | Negative a | ||||
| 11/09/2012 | Extended semen | 18 | Negative a | ||||
| 21/09/2012 | Extended semen | 18 | Negative a | ||||
| 28/09/2012 | Extended semen | 19 | Negative a | ||||
| 26/10/2012 | Extended semen | 20 | Positive (36.5) a | ||||
| 13/11/2012 | Serum | 26 | Negative | >3 | |||
| 7/05/2013 | Serum | 38 | Negative | 3 | |||
| 5/05/2014 | Serum | 50 | Negative | 3 | |||
| 4/05/2015 | Serum | 62 | Negative | 3 | |||
| 2/05/2016 | Serum | 74 | Negative | ≥512 | |||
| 1/05/2017 | Extended semen | 82 | Positive (29.7) | Positive | |||
| 18/01/2018 | Extended semen | 82 | Positive (32.6) | Negative | |||
| 23/01/2018 | Extended semen | 82 | Positive (32.4) | Negative | |||
| 25/01/2018 | Extended semen | 83 | Positive (30.9) | Positive | |||
| 1/02/2018 | Extended semen | 83 | Positive (35.9) | Negative | |||
| 6/02/2018 | Extended semen | 83 | Positive (32.5) | Negative | |||
| 8/02/2018 | Serum | 83 | Negative | ||||
| 12/02/2018 | Extended semen | 83 | Positive (33.3) | Negative | |||
| 13/02/2018 | Extended semen | 83 | Positive (34.4) | Negative | |||
| 15/02/2018 | Extended semen | 83 | Positive (31.58) | Negative | |||
| 16/02/2018 | Extended semen | 83 | Positive (38.46) | ||||
| 20/02/2018 | Extended semen | 83 | Positive (36.82) | ||||
| 22/02/2018 | Extended semen | 83 | Negative | ||||
| 27/02/2018 | Extended semen | 84 | Negative |
| Sample Date | Sample Type | Age (Months) | qRT-PCR (Ct) | Virus Isolation | Antigen ELISA | VNT (Titre) |
|---|---|---|---|---|---|---|
| 20/02/2013 | Serum | 15 | Negative | |||
| 22/05/2013 | Extended semen | 18 | Positive (29.2) | |||
| 24/05/2013 | Extended semen | 18 | Positive (29.7) | Negative | ||
| 28/05/2013 | Extended semen | 18 | Positive (29.3) | Negative | ||
| 30/05/2013 | Extended semen | 18 | Positive (29.1) | |||
| 3/06/2013 | Extended semen | 18 | Positive (32.6) | |||
| 7/06/2013 | Extended semen | 18 | Positive (30.0) | |||
| 10/06/2013 | Serum | 18 | Negative | ≥512 | ||
| 12/06/2013 | Extended semen | 18 | Positive (31.0) | |||
| 14/06/2013 | Extended semen | 19 | Positive (30.5) | |||
| 18/06/2013 | Extended semen | 19 | Positive (29.5) | |||
| 20/06/2013 | Extended semen | 19 | Positive (30.0) | |||
| 24/06/2013 | Extended semen | 19 | Positive (30.4) | |||
| 26/06/2013 | Extended semen | 19 | Positive (30.5) | |||
| 3/07/2013 | Extended semen | 19 | Positive (30.5) | |||
| 8/07/2013 | Extended semen | 19 | Positive (32.0) | |||
| 12/07/2013 | Extended semen | 19 | Positive (31.0) | |||
| 12/07/2013 | Extended semen | 19 | Positive (30.5) | Negative | ||
| 16/07/2013 | Extended semen | 20 | Positive (30.4) | |||
| 18/07/2013 | Extended semen | 20 | Positive (31.4) | |||
| 23/07/2013 | Extended semen | 20 | Positive (32.2) | |||
| 15/08/2013 | Extended semen | 21 | Positive (30.7) | Negative |
| Sample Date | Sample Type | Age (Months) | qRT-PCR (Ct) | Virus Isolation | Antigen ELISA | AGID | VNT (Titre) |
|---|---|---|---|---|---|---|---|
| 3/09/2013 | Serum | 18 | Negative | Negative | |||
| 8/10/2013 | Serum | 19 | Negative | >3 | |||
| 22/10/2013 | Extended semen | 20 | Positive (31.7) | Positive | |||
| 24/10/2013 | Extended semen | 20 | Positive (32.0) | Negative | |||
| 16/12/2013 | Seminal fluid | 22 | Positive (30.2) | Negative | |||
| 16/02/2014 a | Extended semen | 24 | Positive (30.1) | ||||
| 16/02/2014 a | Serum | 24 | ≥512 |
| Sample Date | Sample Type | Age (Months) | qRT-PCR (Ct) | Virus Isolation | Antigen ELISA | VNT (Titre) |
|---|---|---|---|---|---|---|
| 31/03/2014 | Serum | 20 | Negative | 4 | ||
| 12/05/2014 | Serum | 21 | Negative | 256 | ||
| 15/05/2014 | Extended semen | 21 | Positive (32.6) | |||
| 24/05/2014 | Neat semen | 22 | Positive (34.2) | Negative | ||
| 11/06/2014 | Extended semen | 22 | Positive (34.8) | Negative | ||
| 2/07/2014 | Neat semen | 23 | Positive (34.1) | Negative | ||
| 17/07/2014 | Extended semen | 23 | Positive (33.9) | Negative | ||
| 20/02/2015 | Serum | 31 | Negative | ≥512 | ||
| 8/04/2015 | Extended semen | 32 | Negative | |||
| 20/04/2015 | Serum | 32 | Negative | ≥512 | ||
| 22/04/2015 | Extended semen | 33 | Negative | |||
| 27/04/2015 | Extended semen | 33 | Negative | |||
| 14/05/2015 | Extended semen | 33 | Negative | |||
| 19/05/2015 | Extended semen | 33 | Negative | |||
| 21/05/2015 | Extended semen | 34 | Negative | |||
| 25/05/2015 | Extended semen | 34 | Negative | |||
| 28/05/2015 | Extended semen | 34 | Negative | |||
| 2/06/2015 | Extended semen | 34 | Negative | |||
| 4/06/2015 | Extended semen | 34 | Negative | |||
| 8/06/2015 | Extended semen | 34 | Negative | |||
| 12/06/2015 | Extended semen | 34 | Negative | |||
| 16/06/2015 | Extended semen | 34 | Negative | |||
| 28/06/2015 | Extended semen | 35 | Negative | |||
| 18/07/2015 | Extended semen | 35 | Negative | |||
| 17/08/2015 | Extended semen | 36 | Negative | |||
| 21/08/2015 | Extended semen | 37 | Negative | |||
| 24/08/2015 | Extended semen | 37 | Negative | |||
| 31/08/2015 | Extended semen | 38 | Negative | |||
| 6/02/2017 | Serum | 54 | Negative | 512 | ||
| 3/04/2017 | Serum | 56 | 128 | |||
| 1/05/2017 | Extended semen | 57 | Negative | |||
| 4/05/2017 | Extended semen | 57 | Negative |
| Sample Date | Sample Type | Age (Months) | qRT-PCR (Ct) | Virus Isolation | Antigen ELISA | VNT |
|---|---|---|---|---|---|---|
| 14/12/2018 | Extended semen | 15 | Negative | |||
| 6/04/2019 a | Serum | 19 | Negative | |||
| 21/05/2019 | Extended semen | 20 | Positive (27.6) | Positive | ||
| 1/09/2019 | Serum | 24 | Negative | >512 | ||
| 24/09/2019 | Serum | 24 | >512 | |||
| 17/10/2019 | Extended semen | 25 | Positive (38.0) | Negative | ||
| 22/11/2019 | Extended semen | 26 | Positive (36.4) | |||
| 8/01/2020 | Extended semen | 28 | Positive (36.2) | |||
| 23/01/2020 | Extended semen | 28 | Negative |
References
- Richter, V.; Lebl, K.; Baumgartner, W.; Obritzhauser, W.; Kasbohrer, A.; Pinior, B. A systematic worldwide review of the direct monetary losses in cattle due to bovine viral diarrhoea virus infection. Vet. J. 2017, 220, 80–87. [Google Scholar] [CrossRef] [PubMed]
- Smith, D.B.; Meyers, G.; Bukh, J.; Gould, E.A.; Monath, T.; Scott Muerhoff, A.; Pletnev, A.; Rico-Hesse, R.; Stapleton, J.T.; Simmonds, P.; et al. Proposed revision to the taxonomy of the genus Pestivirus, family Flaviviridae. J. Gen. Virol. 2017, 98, 2106–2112. [Google Scholar] [CrossRef] [PubMed]
- Walker, P.J.; Siddell, S.G.; Lefkowitz, E.J.; Mushegian, A.R.; Dempsey, D.M.; Dutilh, B.E.; Harrach, B.; Harrison, R.L.; Hendrickson, R.C.; Junglen, S.; et al. Changes to virus taxonomy and the International Code of Virus Classification and Nomenclature ratified by the International Committee on Taxonomy of Viruses. Arch. Virol. 2019, 164, 2417–2429. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Walz, P.H.; Grooms, D.L.; Passler, T.; Ridpath, J.F.; Tremblay, R.; Step, D.L.; Callan, R.J.; Givens, M.D.; American College of Veterinary Internal, M. Control of bovine viral diarrhea virus in ruminants. J. Vet. Intern. Med. 2010, 24, 476–486. [Google Scholar] [CrossRef] [PubMed]
- Bielanski, A.; Dubuc, C.; Hare, W.C.D. Failure to Remove Bovine Diarrhea Virus (BVDV) from Bull Semen by Swim up and other Separatory Sperm Techniques Associated with in vitro Fertilization. Reprod. Domest. Anim. 1992, 27, 303–306. [Google Scholar] [CrossRef]
- Givens, M.D.; Riddell, K.P.; Edmondson, M.A.; Walz, P.H.; Gard, J.A.; Zhang, Y.; Galik, P.K.; Brodersen, B.W.; Carson, R.L.; Stringfellow, D.A. Epidemiology of prolonged testicular infections with bovine viral diarrhea virus. Vet. Microbiol. 2009, 139, 42–51. [Google Scholar] [CrossRef] [PubMed]
- Givens, M.D.; Riddell, K.P.; Walz, P.H.; Rhoades, J.; Harland, R.; Zhang, Y.; Galik, P.K.; Brodersen, B.W.; Cochran, A.M.; Brock, K.V.; et al. Noncytopathic bovine viral diarrhea virus can persist in testicular tissue after vaccination of peri-pubertal bulls but prevents subsequent infection. Vaccine 2007, 25, 867–876. [Google Scholar] [CrossRef]
- Kirkland, P.D.; McGowan, M.R.; Mackintosh, S.G.; Moyle, A. Insemination of cattle with semen from a bull transiently infected with pestivirus. Vet. Rec. 1997, 140, 124–127. [Google Scholar] [CrossRef]
- Kirkland, P.D.; Richards, S.G.; Rothwell, J.T.; Stanley, D.F. Replication of bovine viral diarrhoea virus in the bovine reproductive tract and excretion of virus in semen during acute and chronic infections. Vet. Rec. 1991, 128, 587–590. [Google Scholar] [CrossRef]
- Whitmore, H.L.; Gustafsson, B.K.; Havareshti, P.; Duchateau, A.B.; Mather, E.C. Inoculation of bulls with bovine virus diarrhea virus: Excretion of virus in semen and effects on semen quality. Theriogenology 1978, 9, 153–163. [Google Scholar] [CrossRef]
- McClurkin, A.W.; Coria, M.F.; Cutlip, R.C. Reproductive performance of apparently healthy cattle persistently infected with bovine viral diarrhea virus. J. Am. Vet. Med. Assoc. 1979, 174, 1116–1119. [Google Scholar] [PubMed]
- Barlow, R.M.; Nettleton, P.F.; Gardiner, A.C.; Greig, A.; Campbell, J.R.; Bonn, J.M. Persistent bovine virus diarrhoea virus infection in a bull. Vet. Rec. 1986, 118, 321–324. [Google Scholar] [CrossRef] [PubMed]
- Howard, T.H.; Bean, B.; Hillman, R.; Monke, D.R. Surveillance for persistent bovine viral diarrhea virus infection in four artificial insemination centers. J. Am. Vet. Med. Assoc. 1990, 196, 1951–1955. [Google Scholar] [PubMed]
- Givens, M.D.; Heath, A.M.; Brock, K.V.; Brodersen, B.W.; Carson, R.L.; Stringfellow, D.A. Detection of bovine viral diarrhea virus in semen obtained after inoculation of seronegative postpubertal bulls. Am. J. Vet. Res. 2003, 64, 428–434. [Google Scholar] [CrossRef] [PubMed]
- Kirkland, P.; Mackintosh, S.; Moyle, A. The outcome of widespread use of semen from a bull persistently infected with pestivirus. Vet. Rec. 1994, 135, 527–529. [Google Scholar] [CrossRef] [PubMed]
- Meyling, A.; Jensen, A.M. Transmission of bovine virus diarrhoea virus (BVDV) by artificial insemination (AI) with semen from a persistently-infected bull. Vet. Microbiol. 1988, 17, 97–105. [Google Scholar] [CrossRef]
- Newcomer, B.W.; Toohey-Kurth, K.; Zhang, Y.; Brodersen, B.W.; Marley, M.S.; Joiner, K.S.; Zhang, Y.; Galik, P.K.; Riddell, K.P.; Givens, M.D. Laboratory diagnosis and transmissibility of bovine viral diarrhea virus from a bull with a persistent testicular infection. Vet. Microbiol. 2014, 170, 246–257. [Google Scholar] [CrossRef] [PubMed]
- Voges, H.; Horner, G.W.; Rowe, S.; Wellenberg, G.J. Persistent bovine pestivirus infection localized in the testes of an immuno-competent, non-viraemic bull. Vet. Microbiol. 1998, 61, 165–175. [Google Scholar] [CrossRef]
- Dunser, M.; Altmann, M.; Dengg, J.; Eichinger, M.; Loitsch, A.; Revilla-Fernandez, S.; Schweighardt, H. Localised persistent infection of the genital tract with bovine viral diarrhoea virus in an immunocompetent bull. Wien. Tierarztl. Mon. 2005, 92, 227–232. [Google Scholar]
- International Office of Epizootics (OIE). Chapter 3.4.7. Bovine viral diarrhoea. In Manual of Diagnostic Tests and Vaccines for Terrestrial Animals; Office international des épizooties: Paris, France, 2019; pp. 1075–1096. [Google Scholar]
- International Office of Epizootics (OIE). Chapter 4.7. Collection and processing of bovine, small ruminant and porcine semen. In Terrestrial Animal Health Code; Office international des épizooties: Paris, France, 2019. [Google Scholar]
- Kirkland, P.D.; MacKintosh, S.G. Ruminant Pestivirus Infections. In Australia and New Zealand Standard Diagnostic Procedures; Department of Agriculture, Australian Government: Canberra, Australia, 2006; pp. 1–30. [Google Scholar]
- Mahony, T.J.; McCarthy, F.M.; Gravel, J.L.; Corney, B.; Young, P.L.; Vilcek, S. Genetic analysis of bovine viral diarrhoea viruses from Australia. Vet. Microbiol. 2005, 106, 1–6. [Google Scholar] [CrossRef]
- Gong, Y.; Trowbridge, R.; Macnaughton, T.B.; Westaway, E.G.; Shannon, A.D.; Gowans, E.J. Characterization of RNA synthesis during a one-step growth curve and of the replication mechanism of bovine viral diarrhoea virus. J. Gen. Virol. 1996, 77 Pt 11, 2729–2736. [Google Scholar] [CrossRef]
- Gu, X.; Davis, R.J.; Walsh, S.J.; Melville, L.F.; Kirkland, P.D. Longitudinal study of the detection of Bluetongue virus in bull semen and comparison of real-time polymerase chain reaction assays. J. Vet. Diagn. Investig. 2014, 26, 18–26. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hoffmann, B.; Depner, K.; Schirrmeier, H.; Beer, M. A universal heterologous internal control system for duplex real-time RT-PCR assays used in a detection system for pestiviruses. J. Virol. Methods 2006, 136, 200–209. [Google Scholar] [CrossRef] [PubMed]
- Brown, L.M.; Papa, R.A.; Frost, M.J.; Mackintosh, S.G.; Gu, X.; Dixon, R.J.; Shannon, A.D. A single amino acid is critical for the expression of B-cell epitopes on the helicase domain of the pestivirus NS3 protein. Virus Res. 2002, 84, 111–124. [Google Scholar] [CrossRef]
- Hueston, W.D.; Monke, D.R.; Milburn, R.J. Scrotal circumference measurements on young Holstein bulls. J. Am. Vet. Med. Assoc. 1988, 192, 766–768. [Google Scholar]
- Wolf, F.R.; Almquist, J.O.; Hale, E.B. Prepuberal Behavior and Puberal Characteristics of Beef Bulls on High Nutrient Allowance. J. Anim. Sci. 1965, 24, 761–765. [Google Scholar] [CrossRef] [PubMed]
- Brito, L.F.; Barth, A.D.; Rawlings, N.C.; Wilde, R.E.; Crews, D.H., Jr.; Mir, P.S.; Kastelic, J.P. Effect of nutrition during calfhood and peripubertal period on serum metabolic hormones, gonadotropins and testosterone concentrations, and on sexual development in bulls. Domest. Anim. Endocrinol. 2007, 33, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Dance, A.; Thundathil, J.; Wilde, R.; Blondin, P.; Kastelic, J. Enhanced early-life nutrition promotes hormone production and reproductive development in Holstein bulls. J. Dairy Sci. 2015, 98, 987–998. [Google Scholar] [CrossRef]
- Harstine, B.R.; Maquivar, M.; Helser, L.A.; Utt, M.D.; Premanandan, C.; DeJarnette, J.M.; Day, M.L. Effects of dietary energy on sexual maturation and sperm production in Holstein bulls. J. Anim. Sci. 2015, 93, 2759–2766. [Google Scholar] [CrossRef]
- Fawcett, D.W.; Leak, L.V.; Heidger, P.M., Jr. Electron microscopic observations on the structural components of the blood-testis barrier. J. Reprod Fertil Suppl. 1970, 10, 105–122. [Google Scholar]
- Mital, P.; Hinton, B.T.; Dufour, J.M. The blood-testis and blood-epididymis barriers are more than just their tight junctions. Biol. Reprod. 2011, 84, 851–858. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Setchell, B.P.; Pollanen, P.; Zupp, J.L. Development of the blood-testis barrier and changes in vascular permeability at puberty in rats. Int. J. 1988, 11, 225–233. [Google Scholar] [CrossRef] [PubMed]
- Niskanen, R.; Alenius, S.; Belak, K.; Baule, C.; Belak, S.; Voges, H.; Gustafsson, H. Insemination of susceptible heifers with semen from a non-viraemic bull with persistent bovine virus diarrhoea virus infection localized in the testes. Reprod. Domest. Anim. 2002, 37, 171–175. [Google Scholar] [CrossRef] [PubMed]



| Bull Number | Breed | Estimated Age at First Detection of Infection (Months) a | Estimated Duration of Shedding (Months) b | qRT-PCR Results in Semen (Ct) | Virus Isolation (Pos/Total Tested) |
|---|---|---|---|---|---|
| 1 | Main Anjou | 21 | 3 | 32.8–35.8 | 0/6 |
| 2 | Holstein | 10 | 73 | 29.7–38.4 | 2/10 |
| 3 | Angus | 18 | 3 | 29.1–32.6 | 0/5 |
| 4 | Angus | 19 | 4 | 30.1–32.0 | 1/3 |
| 5 | Angus | 20 | 6 | 32.6–34.8 | 0/6 |
| 6 | Wagyu | 21 | 8 | 27.6–38.0 | 1/2 c |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Read, A.J.; Gestier, S.; Parrish, K.; Finlaison, D.S.; Gu, X.; O’Connor, T.W.; Kirkland, P.D. Prolonged Detection of Bovine Viral Diarrhoea Virus Infection in the Semen of Bulls. Viruses 2020, 12, 674. https://doi.org/10.3390/v12060674
Read AJ, Gestier S, Parrish K, Finlaison DS, Gu X, O’Connor TW, Kirkland PD. Prolonged Detection of Bovine Viral Diarrhoea Virus Infection in the Semen of Bulls. Viruses. 2020; 12(6):674. https://doi.org/10.3390/v12060674
Chicago/Turabian StyleRead, Andrew J., Sarah Gestier, Kate Parrish, Deborah S. Finlaison, Xingnian Gu, Tiffany W. O’Connor, and Peter D. Kirkland. 2020. "Prolonged Detection of Bovine Viral Diarrhoea Virus Infection in the Semen of Bulls" Viruses 12, no. 6: 674. https://doi.org/10.3390/v12060674