Usutu Virus Infection of Embryonated Chicken Eggs and a Chicken Embryo-Derived Primary Cell Line

 ,
,  ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Viruses and Embryonated Chicken Eggs
2.2. In Ovo Characterization of USU-BE-Seraing/2017
2.3. Virulence of Other USUV Strains In Ovo
2.4. Preparation of Primary Chorioallantoic Membrane Cells
2.5. Characterization of USUV Strains Growth Kinetics in Chorioallantoic Membrane Cells
2.6. Statistical Analyses
3. Results
3.1. In Ovo Characterization of USUV USU-BE-Seraing/2017
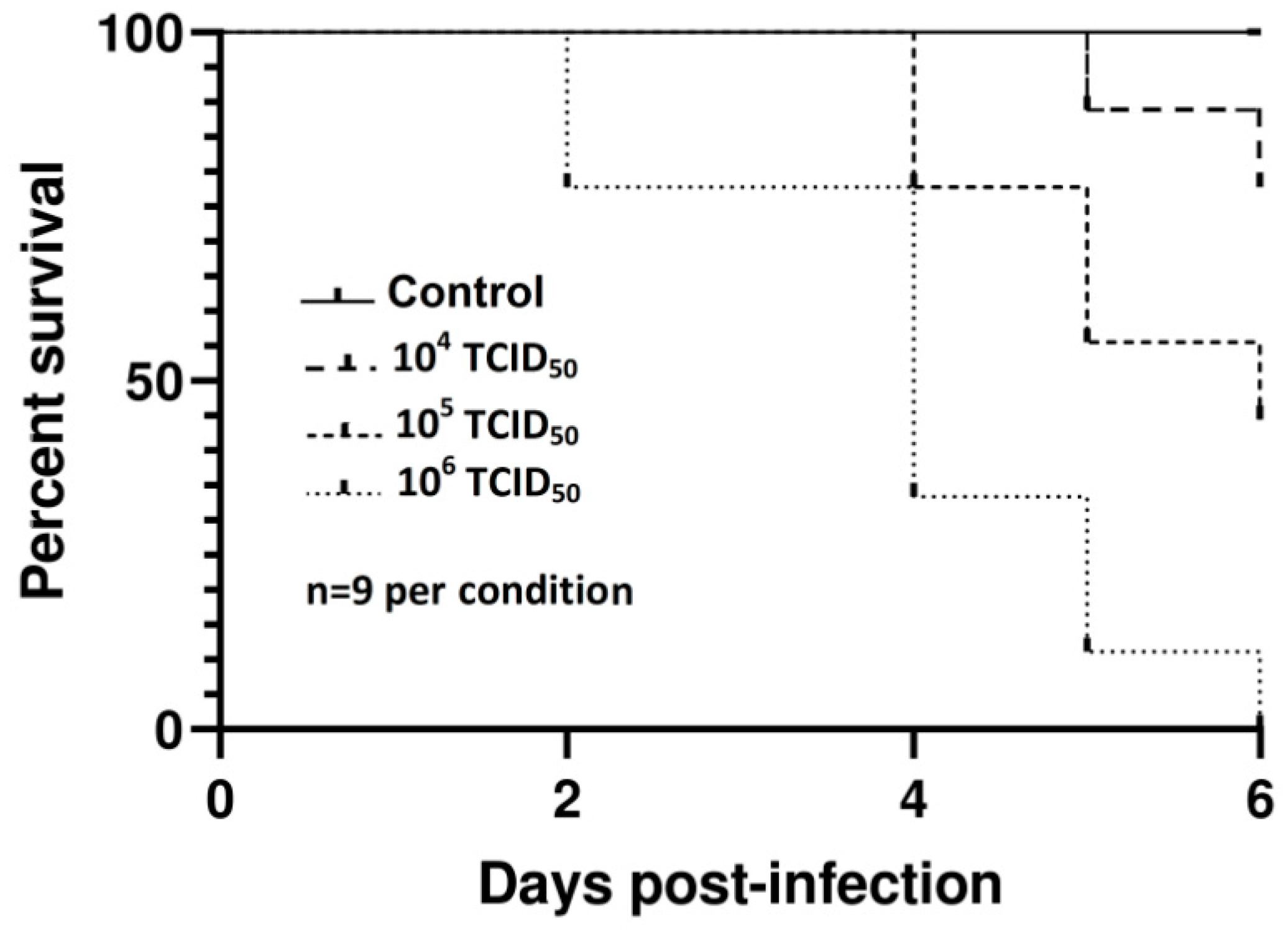
3.1.1. Survival Study
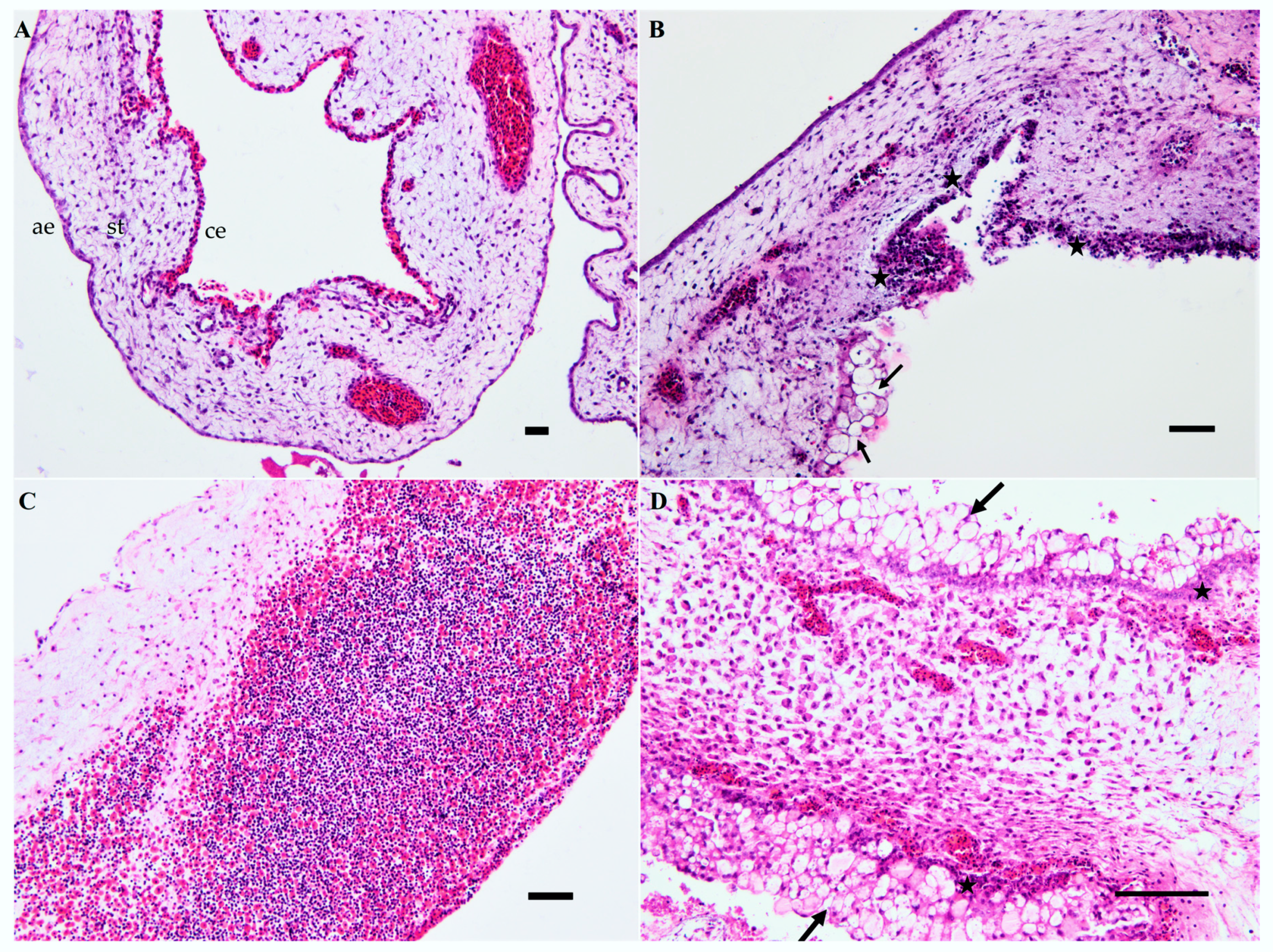
3.1.2. Course of Infection
3.2. Virulence of other USUV Strains In Ovo
3.3. Characterization of USUV Strains Growth Kinetics in Chorioallantoic Membrane Cells
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Kuno, G.; Chang, G.J.; Tsuchiya, K.R.; Karabatsos, N.; Cropp, C.B. Phylogeny of the genus flavivirus. J. Virol. 1998, 72, 73–83. [Google Scholar] [CrossRef] [Green Version]
- Nikolay, B. A review of West Nile and Usutu virus co-circulation in Europe: How much do transmission cycles overlap? Trans. R. Soc. Trop. Med. Hyg. 2015, 109, 609–618. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- IZSAM. CESME Integrated Surveillance of West Nile and Usutu Virus. Available online: https://westnile.izs.it/j6_wnd/home (accessed on 2 December 2019).
- Benzarti, E.; Linden, A.; Desmecht, D.; Garigliany, M. Mosquito-borne epornitic flaviviruses: An update and review. J. Gen. Virol. 2019, 100, 119–132. [Google Scholar] [CrossRef] [PubMed]
- Weidinger, P.; Kolodziejek, J.; Bakonyi, T.; Brunthaler, R.; Erdélyi, K.; Eissenböck, H.; Nowotny, N. Different dynamics of Usutu virus infections in Austria and Hungary, 2017–2018. Trans. Emerg. Dis. 2020, 67, 298–307. [Google Scholar] [CrossRef]
- Benzarti, E.; Garigliany, M.; Hauman, D.; Paternostre, J.; Linden, A.; Franssen, M.; Sarlet, M.; Cassart, D.; Desmecht, D. First evidence of fatal Usutu virus natural infections in an Anatidae, the common scoter (Melanitta nigra). Vector Borne Zoonotic Dis. 2019, 19, 777–780. [Google Scholar] [CrossRef]
- Lühken, R.; Jöst, H.; Cadar, D.; Thomas, S.M.; Bosch, S.; Tannich, E.; Becker, N.; Ziegler, U.; Lachmann, L.; Schmidt-chanasit, J. Distribution of Usutu virus in Germany and its effect on breeding bird populations. Emerg. Infect. Dis. 2017, 23, 1991–1998. [Google Scholar] [CrossRef] [Green Version]
- Savini, G.; Monaco, F.; Terregino, C.; Di Gennaro, A.; Bano, L.; Pinoni, C.; De Nardi, R.; Bonilauri, P.; Pecorari, M.; Di Gialleonardo, L.; et al. Usutu virus in Italy: An emergence or a silent infection? Vet. Microbiol. 2011, 151, 264–274. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Allering, L.; Jöst, H.; Emmerich, P.; Günther, S.; Lattwein, E.; Schmidt, M.; Seifried, E.; Sambri, V.; Hourfar, K.; Medizinische, E.; et al. Detection of Usutu virus infection in a healthy blood donor from south-west Germany, 2012. Euro Surveill. 2012, 17, 20341. [Google Scholar] [PubMed]
- Cvjetković, I.H.; Petrović, T.; Petrić, D.; Cvjetković, D.; Kovačević, G.; Radovanov, J.; Galović, A.J.; Patić, A.; Nikolić, N.; Mikić, S.S.; et al. Seroprevalence of mosquito-born and tick-born microorganisms in human population of South Backa District. Arh. Vet. Med. 2016, 9, 23–30. [Google Scholar]
- Grottola, A.; Marcacci, M.; Tagliazucchi, S.; Gennari, W.; Di Gennaro, A.; Orsini, M.; Marchegiano, P.; Marini, V.; Meacci, M.; Rumpianesi, F.; et al. Usutu virus infections in humans: A retrospective analysis in the municipality of Modena, Italy. Clin. Microbiol. Infect. 2017, 23, 33–37. [Google Scholar] [CrossRef] [Green Version]
- Percivalle, E.; Sassera, D.; Rovida, F.; Isernia, P.; Fabbi, M. Usutu virus antibodies in blood donors and healthy forestry workers in the Lombardy Region, Northern Italy. Vector Borne Zoonotic Dis. 2017, 17, 658–661. [Google Scholar] [CrossRef]
- Zaaijer, H.L.; Slot, E.; Molier, M.; Reusken, C.B.E.M.; Koppelman, M.H.G.M. Usutu virus infection in Dutch blood donors. Transfusion 2019, 59, 2931–2937. [Google Scholar] [CrossRef]
- Nagy, A.; Mezei, E.; Nagy, O.; Bakonyi, T.; Csonka, N.; Kaposi, M.; Koroknai, A.; Szomor, K.; Rigó, Z.; Molnár, Z.; et al. Extraordinary increase in West Nile virus cases and first confirmed human Usutu virus infection in Hungary, 2018. Eurosurveillance 2019, 24, 1900038. [Google Scholar] [CrossRef] [Green Version]
- Vilibic-Cavlek, T.; Savic, V.; Sabadi, D.; Peric, L.; Barbic, L.; Klobucar, A.; Miklausic, B.; Tabain, I.; Santini, M.; Vucelja, M.; et al. Prevalence and molecular epidemiology of West Nile and Usutu virus infections in Croatia in the ‘One health’ context, 2018. Transbound. Emerg. Dis. 2019, 66, 1946–1957. [Google Scholar] [CrossRef]
- Clé, M.; Salinas, S.; Lecollinet, S.; Beck, C.; Gutierrez, S.; Baldet, T.; Perre, P.V.; Foulongne, V.; Simonin, Y.; Usutu, L. Le virus Usutu: La menace fantôme. Med. Sci. 2018, 34, 709–716. [Google Scholar] [CrossRef] [Green Version]
- Cavrini, F.; Gaibani, P.; Longo, G.; Pierro, A.; Rossini, G.; Bonilauri, P.; Gerunda, G.; Di Benedetto, F.; Pasetto, A.; Girardis, M.; et al. Usutu virus infection in a patient who underwent orthotropic liver transplantation, Italy, August–September 2009. Euro Surveill. 2009, 14, 19448. [Google Scholar]
- Pecorari, M.; Longo, G.; Gennari, W.; Grottola, A.; Sabbatini, A.M.T.; Tagliazucchi, S. First human case of Usutu virus neuroinvasive infection, Italy, August–September 2009. Euro Surveill. 2009, 14, 19446. [Google Scholar]
- Santini, M.; Vilibic-Cavlek, T.; Barsic, B.; Barbic, L.; Savic, V.; Stevanovic, V.; Listes, E.; Di Gennaro, A.; Savini, G. First cases of human Usutu virus neuroinvasive infection in Croatia, August–September 2013: Clinical and laboratory features. J. Neurovirol. 2014, 21, 92–97. [Google Scholar] [CrossRef]
- Simonin, Y.; Sillam, O.; Carles, M.J.; Gutierrez, S.; Gil, P.; Constant, O.; Martin, M.F.; Girard, G.; Van De Perre, P.; Salinas, S.; et al. Human Usutu virus infection with atypical neurologic presentation, Montpellier, France, 2016. Emerg. Infect. Dis. 2018, 24, 875–878. [Google Scholar] [CrossRef] [Green Version]
- Heinz, F.X.; Stiasny, K. Flaviviruses and their antigenic structure. J. Clin. Virol. 2012, 55, 289–295. [Google Scholar] [CrossRef]
- Cook, C.L.; Huang, Y.J.S.; Lyons, A.C.; Alto, B.W.; Unlu, I.; Higgs, S.; Vanlandingham, D.L. North American Culex pipiens and Culex quinquefasciatus are competent vectors for Usutu virus. PLoS Negl. Trop. Dis. 2018, 12, e0006732. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roesch, F.; Fajardo, A.; Moratorio, G.; Vignuzzi, M. Usutu virus: An arbovirus on the rise. Viruses 2019, 11, 640. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blyden, B.E.R.; Watler, P.K. New approaches to improved vaccine manufacturing in embryonated eggs. BioPharm Int. Suppl. 2010, 2010, 4–9. [Google Scholar]
- Lin, J.; Liu, Y.; Wang, X.; Yang, B.; He, P.; Yang, Z.; Xie, J.; Zou, L.; Zhao, J.; Pan, J.; et al. Efficacy evaluation of an inactivated duck Tembusu virus vaccine. Avian Dis. 2015, 59, 244–248. [Google Scholar] [CrossRef] [PubMed]
- Ianconescu, M.; Hornstein, K.; Samberg, Y.; Aharonovici, A. Development of a new vaccine against turkey meningo-encephalitis using a virus passaged through the Japanese quail (Coturnix coturnix Japonica). Avian Pathol. 1975, 4, 119–131. [Google Scholar]
- Thawani, A.; Sirohi, D.; Kuhn, R.J.; Fekete, D.M. Zika virus can strongly infect and disrupt secondary organizers in the ventricular zone of the embryonic chicken brain. Physiol. Behav. 2017, 176, 139–148. [Google Scholar] [CrossRef] [Green Version]
- Goodfellow, F.T.; Tesla, B.; Simchick, G.; Zhao, Q.; Hodge, T.; Brindley, M.A.; Stice, S.L. Zika virus induced mortality and microcephaly in chicken embryos. Stem Cells Dev. 2016, 25, 1691–1697. [Google Scholar] [CrossRef]
- Ishikawa, T.; Yamanaka, A.; Konishi, E. A review of successful flavivirus vaccines and the problems with those flaviviruses for which vaccines are not yet available. Vaccine 2014, 32, 1326–1337. [Google Scholar] [CrossRef]
- De Abreu Manso, P.P.; Dias de Oliveira, B.C.E.P.; De Sequeira, P.C.; De Souza, Y.R.M.; Dos Santos Ferro, J.M.; Da Silva, I.J.; Caputo, L.F.G.; Guedes, P.T.; Dos Santos, A.A.C.; Da Silva Freire, M.; et al. Yellow fever 17DD vaccine virus infection causes detectable changes in chicken embryos. PLoS Negl. Trop. Dis. 2015, 9, e0004064. [Google Scholar]
- Bakonyi, T.; Lussy, H.; Weissenböck, H.; Hornyák, Á.; Nowotny, N. In vitro host-cell susceptibility to Usutu virus. Emerg. Infect. Dis. 2005, 11, 298–301. [Google Scholar] [CrossRef]
- Segura, N.A.; Sharma, S.; Neyts, J.; Kaptein, S.J.F. Favipiravir inhibits in vitro Usutu virus replication and delays disease progression in an infection model in mice. Antivir. Res. 2018, 160, 137–142. [Google Scholar] [CrossRef] [PubMed]
- Osorio, J.E.; Ciuoderis, K.A.; Lopera, J.G.; Piedrahita, L.D.; Murphy, D.; LeVasseur, J.; Carrillo, L.; Ocampo, M.C.; Hofmeister, E. Characterization of West Nile viruses isolated from captive American flamingoes (Phoenicopterus ruber) in Medellin, Colombia. Am. J. Trop. Med. Hyg. 2012, 87, 565–572. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, L.; Li, Y.; Zhang, Y.; Han, Z.; Xu, Y.; Kong, X.; Liu, S. Adaptation and attenuation of duck tembusu virus strain Du/CH/LSD/110128 following serial passage in chicken embryos. Clin. Vaccine Immunol. 2014, 21, 1046–1053. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Way, H.J.; Bowen, E.T.W.; Platt, G.S. Comparative studies of some African arboviruses in cell culture and in mice. J. Gen. Virol. 1976, 30, 123–130. [Google Scholar] [CrossRef]
- Chan, J.F.W.; Yip, C.C.Y.; Tsang, J.O.L.; Tee, K.M.; Cai, J.P.; Chik, K.K.H.; Zhu, Z.; Chan, C.C.S.; Choi, G.K.Y.; Sridhar, S.; et al. Differential cell line susceptibility to the emerging Zika virus: Implications for disease pathogenesis, non-vector-borne human transmission and animal reservoirs. Emerg. Microbes Infect. 2016, 5, e93. [Google Scholar] [CrossRef] [Green Version]
- Benzarti, E.; Sarlet, M.; Franssen, M.; Cadar, D.; Schmidt-Chanasit, J.; Rivas, J.; Linden, A.; Desmecht, D.; Garigliany, M. Usutu virus epizootic in Belgium in 2017 and 2018: Evidence of virus endemization and ongoing introduction events. Vector Borne Zoonotic Dis. 2019, 20, 43–50. [Google Scholar] [CrossRef]
- Garigliany, M.; Linden, A.; Gilliau, G.; Levy, E.; Sarlet, M.; Franssen, M.; Benzarti, E.; Derouaux, A.; Francis, F.; Desmecht, D. Usutu virus, Belgium, 2016. Infect. Genet. Evol. 2017, 48, 116–119. [Google Scholar] [CrossRef]
- Benzarti, E.; Rivas, J.; Sarlet, M.; Franssen, M.; Desmecht, D.; Schmidt-Chanasit, J.; Savini, G.; Lorusso, A.; Van Laere, A.-S.; Garigliany, M.-M. Experimental Usutu virus infection in domestic canaries Serinus canaria. Viruses 2020, 12, 164. [Google Scholar] [CrossRef] [Green Version]
- Schilling, M.A.; Katani, R.; Memari, S.; Cavanaugh, M.; Buza, J.; Basu, J.R.; Mpenda, F.N.; Deist, M.S.; Lamont, S.J.; Kapur, V. Transcriptional innate immune response of the developing chicken embryo to Newcastle disease virus infection. Front. Genet. 2018, 9, 61. [Google Scholar] [CrossRef]
- Deist, M.S.; Gallardo, R.A.; Bunn, D.A.; Dekkers, J.C.M.; Zhou, H.; Lamont, S.J. Resistant and susceptible chicken lines show distinctive responses to Newcastle disease virus infection in the lung transcriptome. BMC Genom. 2017, 18, 989. [Google Scholar] [CrossRef] [Green Version]
- Scagnolari, C.; Caputo, B.; Trombetti, S.; Spano, L.; Villari, P.; Cacciotti, G.; Solda, A.; Torre, A.; Nowotny, N.; Antonelli, G. Usutu virus growth in human cell lines: Induction of and sensitivity to type I and III interferons Communication Usutu virus growth in human cell lines: Induction of and sensitivity to type I and III interferons. J. Gen. Virol. 2013, 94, 789–795. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cacciotti, G.; Caputo, B.; Selvaggi, C.; Vitiello, L.; Diallo, D.; Ceianu, C.; Antonelli, G.; Nowotny, N.; Scagnolari, C. Variation in interferon sensitivity and induction between Usutu and West Nile (lineages 1 and 2) viruses. Virology 2015, 485, 189–198. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weissenbock, H.; Bakonyi, T.; Chvala, S.; Nowotny, N. Experimental Usutu virus infection of suckling mice causes neuronal and glial cell apoptosis and demyelination. Acta Neuropathol. 2004, 108, 453–460. [Google Scholar] [CrossRef] [PubMed]
- Blázquez, A.; Escribano-romero, E.; Martín-acebes, M.A.; Petrovic, T.; Saiz, J. Limited susceptibility of mice to Usutu virus (USUV) infection and induction of flavivirus cross-protective immunity. Virology 2015, 482, 67–71. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martín-Acebes, M.A.; Blázqueza, A.-B.; Canas-Arranz, R.; Vázquez-calvo, Á.; Merino-ramos, T.; Escribano-romero, E.; Sobrino, F.; Saiz, J. A recombinant DNA vaccine protects mice deficient in the alpha/beta interferon receptor against lethal challenge with Usutu virus. Vaccine 2016, 34, 2066–2073. [Google Scholar] [CrossRef]
- Takamatsu, Y.; Okamoto, K.; Dinh, D.T.; Yu, F.; Hayasaka, D.; Uchida, L.; Nabeshima, T.; Buerano, C.C.; Morita, K. NS1′ protein expression facilitates production of Japanese encephalitis virus in avian cells and embryonated chicken eggs. J. Gen. Virol. 2014, 95, 373–383. [Google Scholar] [CrossRef]
- Chvala, S.; Bakonyi, T.; Hackl, R.; Hess, M.; Nowotny, N.; Weissenböck, H. Limited pathogenicity of Usutu virus for the domestic goose (Anser anser f. domestica) following experimental inoculation. J. Vet. Med. 2006, 53, 171–175. [Google Scholar] [CrossRef]
- Crespo, R.; Shivaprasad, H.L.; França, M.; Woolcock, P.R.; Crespo, R.; Shivaprasad, H.L.; Franc, A.M.; Woolcock, P.R. Isolation and distribution of West Nile virus in embryonated chicken eggs. Avian Dis. 2009, 53, 608–612. [Google Scholar] [CrossRef]
- Yan, P.; Zhao, Y.; Zhang, X.; Xu, D.; Dai, X.; Teng, Q.; Yan, L. An infectious disease of ducks caused by a newly emerged Tembusu virus strain in mainland China. Virology 2011, 417, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Thontiravong, A.; Ninvilai, P.; Tunterak, W.; Nonthabenjawan, N.; Chaiyavong, S.; Angkabkingkaew, K.; Mungkundar, C.; Phuengpho, W.; Oraveerakul, K.; Amonsin, A.; et al. Tembusu-related flavivirus in ducks, Thailand. Emerg. Infect. Dis. 2015, 21, 2164–2167. [Google Scholar] [CrossRef] [Green Version]
- Salinas, S.; Constant, O.; Desmetz, C.; Barthelemy, J.; Lemaitre, M.; Milhavet, O.; Nagot, N.; Foulongne, V.; Perrin, E.; Saiz, J.; et al. Deleterious effect of Usutu virus on human neural cells. PLoS Negl. Trop. Dis. 2017, 11, e0005913. [Google Scholar] [CrossRef] [PubMed]
- Pauli, A.M.; Cruz-martinez, L.A.; Ponder, J.B.; Redig, P.T.; Glaser, A.L.; Klauss, G.; Schoster, J.V. Ophthalmologic and oculopathologic finding in red-tailed hawks and Cooper’s hawks with naturally acquired West Nile virus infection. J. Am. Vet. Med. Assoc. 2007, 231, 1240–1248. [Google Scholar] [CrossRef]
- Wünschmann, A.; Shivers, J.; Bender, J.; Carroll, L.; Fuller, S.; Saggese, M.; Van Wettere, A.; Redig, P.; Shivers, A.J.; Bender, A.J.; et al. Pathologic and immunohistochemical findings in goshawks (Accipiter gentilis) and great horned owls (Bubo virginianus) naturally infected with West Nile virus. Avian Dis. 2005, 49, 252–259. [Google Scholar] [CrossRef] [PubMed]
- Gamino, V.; Fernández-de-mera, I.G.; Ortíz, J.; Durán-martín, M.; De Fuente, J.; Gortázar, C.; Höfle, U. Natural Bagaza virus infection in game birds in southern Spain. Vet. Res. 2012, 43, 65. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gómez, C.C.; Llorente, F.; Ramírez, E.P.; Soriguer, R.C.; Sarasa, M.; Ángel, M.; Clavero, J. Experimental infection of grey partridges with Bagaza virus: Pathogenicity evaluation and potential role as a competent host. Vet. Res. 2018, 49, 44. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nemeth, N.M.; Young, G.R.; Burkhalter, K.L.; Brault, A.C.; Reisen, W.K.; Komar, N. West Nile virus detection in nonvascular feathers from avian carcasses. J. Vet. Diagn. Investig. 2009, 21, 616–622. [Google Scholar] [CrossRef] [Green Version]
- Davidson, I.; Natour-altory, A.; Raibstein, I.; Kin, E.; Dahan, Y.; Krispin, H. Monitoring the uptake of live avian vaccines by their detection in feathers. Vaccine 2018, 36, 637–643. [Google Scholar] [CrossRef]
- Freire, M.S.; Mann, G.F.; Marchevsky, R.S.; Yamamura, A.M.Y.; Almeida, L.F.C.; Jabor, A.V.; Malachias, J.M.N.; Coutinho, E.S.F.; Galler, R. Production of yellow fever 17DD vaccine virus in primary culture of chicken embryo fibroblasts: Yields, thermo and genetic stability, attenuation and immunogenicity. Vaccine 2005, 23, 2501–2512. [Google Scholar] [CrossRef] [Green Version]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Viral Dose (TCID50) | Log-Rank (Mantel-Cox) | Gehan–Breslow Wilcoxon | ||
|---|---|---|---|---|
| χ2 | p | χ2 | p | |
| 106 | 3.846 | 0.2752 | 3.537 | 0.316 |
| 105 | 2.033 | 0.5655 | 2.203 | 0.5113 |
| 104 | 0.03672 | 0.9981 | 8.845e-032 | >0.9999 |
| Tissue | Infection with USU-BE-Seraing/2017 | IHC Findings in Embryos Infected with Other USUV Strains * | ||||
|---|---|---|---|---|---|---|
| dpi | ||||||
| 1 | 2 | 3 | 4 | 5 | ||
| CAM | - | ++ | +++ | +++ | +++ | Common to all strains |
| Brain | - | - | - | - | - | Positive staining when infected with USU-BE-Grivegnee/2017 and UR-10-Tm strains (day 5 pi) |
| Heart | - | - | + | + | + | Common to all strains |
| Liver | - | - | + | - | - | Only with USU-BE-Seraing/2017 |
| Skeletal muscle | - | + | + | + | + | Common to all strains |
| Intestine | - | - | - | - | + | Positive staining with USU-BE-Grivegnee/2017 |
| Eye | - | + | + | - | - | Only with USU-BE-Seraing/2017 |
| Skin and feather follicles | - | - | + | ++ | +++ | Common to all strains |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Benzarti, E.; Rivas, J.; Sarlet, M.; Franssen, M.; Moula, N.; Savini, G.; Lorusso, A.; Desmecht, D.; Garigliany, M.-M. Usutu Virus Infection of Embryonated Chicken Eggs and a Chicken Embryo-Derived Primary Cell Line. Viruses 2020, 12, 531. https://doi.org/10.3390/v12050531
Benzarti E, Rivas J, Sarlet M, Franssen M, Moula N, Savini G, Lorusso A, Desmecht D, Garigliany M-M. Usutu Virus Infection of Embryonated Chicken Eggs and a Chicken Embryo-Derived Primary Cell Line. Viruses. 2020; 12(5):531. https://doi.org/10.3390/v12050531
Chicago/Turabian StyleBenzarti, Emna, José Rivas, Michaël Sarlet, Mathieu Franssen, Nassim Moula, Giovanni Savini, Alessio Lorusso, Daniel Desmecht, and Mutien-Marie Garigliany. 2020. "Usutu Virus Infection of Embryonated Chicken Eggs and a Chicken Embryo-Derived Primary Cell Line" Viruses 12, no. 5: 531. https://doi.org/10.3390/v12050531