Evidence Supporting That RNA Polymerase II Catalyzes De Novo Transcription Using Potato Spindle Tuber Viroid Circular RNA Templates



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
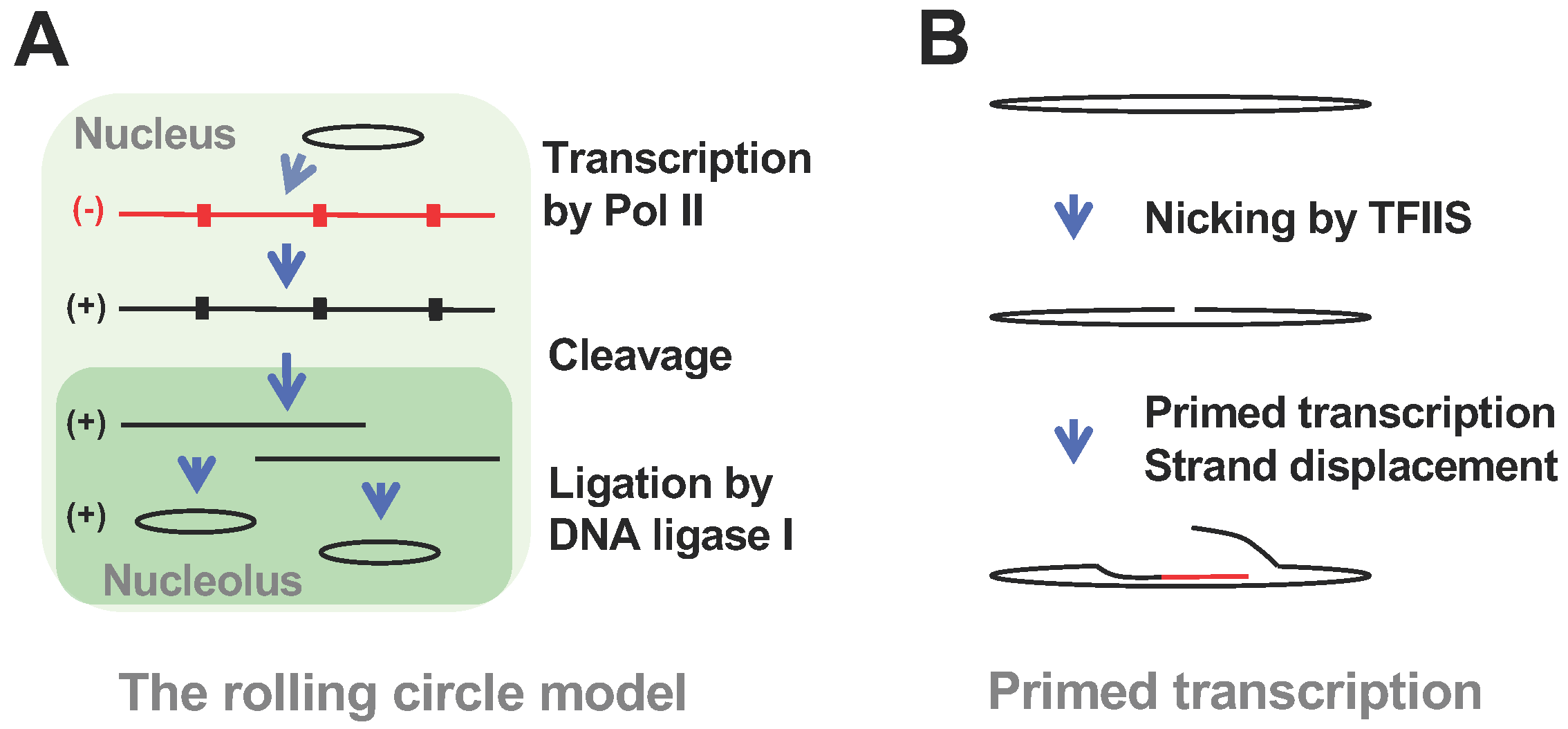
:1. Introduction
2. Materials and Methods
2.1. Plant Growth and Protoplast Assays
2.2. Recombinant Protein Purification
2.3. Purification of Pol II from Wheat Germ
2.4. Immunoblotting
2.5. RNA Preparation In Vitro
2.6. Pol II-catalyzed In Vitro Transcription
2.7. RNA Purification. RNA Gel Blots, and Dot Blots
3. Results
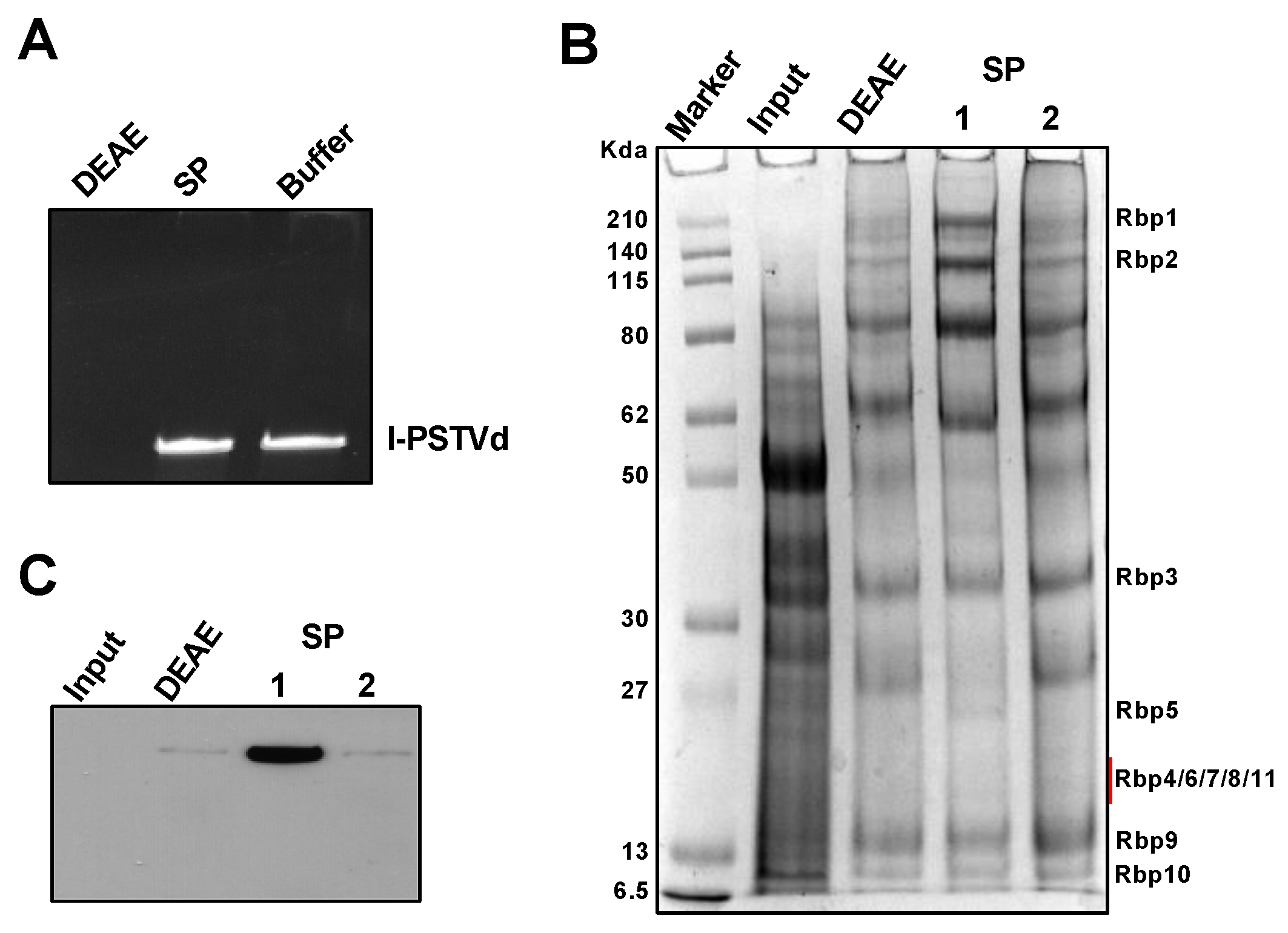
3.1. An Efficient Method to Partially Purify Functional Pol II
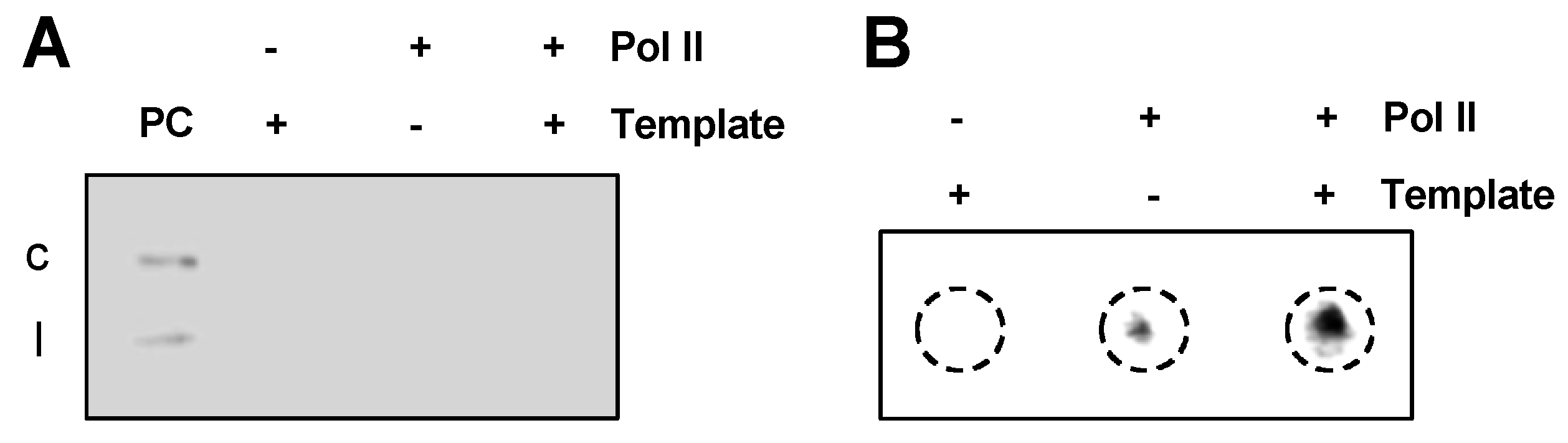
3.2. Circular Templates are Critical for Generating Longer-Than-Unit-Length Intermediates
3.3. PSTVd Replication is Independent of TFIIS
4. Discussion
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Spitz, F.; Furlong, E.E. Transcription factors: From enhancer binding to developmental control. Nat. Rev. Genet. 2012, 13, 613–626. [Google Scholar] [CrossRef]
- Levine, M.; Tjian, R. Transcription regulation and animal diversity. Nature 2003, 424, 147–151. [Google Scholar] [CrossRef]
- Crick, F.H. On protein synthesis. Symp. Soc. Exp. Biol. 1958, 12, 138–163. [Google Scholar]
- Crick, F. Central dogma of molecular biology. Nature 1970, 227, 561–563. [Google Scholar] [CrossRef]
- Khatter, H.; Vorlander, M.K.; Muller, C.W. RNA polymerase I and III: Similar yet unique. Curr. Opin. Struct. Biol. 2017, 47, 88–94. [Google Scholar] [CrossRef]
- Thomas, M.C.; Chiang, C.M. The general transcription machinery and general cofactors. Crit. Rev. Biochem. Mol. Biol. 2006, 41, 105–178. [Google Scholar] [CrossRef]
- Ream, T.S.; Haag, J.R.; Wierzbicki, A.T.; Nicora, C.D.; Norbeck, A.D.; Zhu, J.K.; Hagen, G.; Guilfoyle, T.J.; Pasa-Tolic, L.; Pikaard, C.S. Subunit compositions of the RNA-silencing enzymes Pol IV and Pol V reveal their origins as specialized forms of RNA polymerase II. Mol. Cell 2009, 33, 192–203. [Google Scholar] [CrossRef] [Green Version]
- Roeder, R.G.; Rutter, W.J. Multiple forms of DNA-dependent RNA polymerase in eukaryotic organisms. Nature 1969, 224, 234–237. [Google Scholar] [CrossRef]
- Hantsche, M.; Cramer, P. Conserved RNA polymerase II initiation complex structure. Curr. Opin. Struct. Biol. 2017, 47, 17–22. [Google Scholar] [CrossRef] [Green Version]
- Martinez-Rucobo, F.W.; Cramer, P. Structural basis of transcription elongation. Biochim. Biophys. Acta 2013, 1829, 9–19. [Google Scholar] [CrossRef] [Green Version]
- Vannini, A.; Cramer, P. Conservation between the RNA polymerase I, II, and III transcription initiation machineries. Mol. Cell 2012, 45, 439–446. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hahn, S.; Young, E.T. Transcriptional regulation in Saccharomyces cerevisiae: Transcription factor regulation and function, mechanisms of initiation, and roles of activators and coactivators. Genetics 2011, 189, 705–736. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grunberg, S.; Warfield, L.; Hahn, S. Architecture of the RNA polymerase II preinitiation complex and mechanism of ATP-dependent promoter opening. Nat. Struct. Mol. Biol. 2012, 19, 788–796. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, Y.; Bernecky, C.; Lee, C.T.; Maier, K.C.; Schwalb, B.; Tegunov, D.; Plitzko, J.M.; Urlaub, H.; Cramer, P. Architecture of the RNA polymerase II-Paf1C-TFIIS transcription elongation complex. Nat. Commun. 2017, 8, 15741. [Google Scholar] [CrossRef] [Green Version]
- Dezelee, S.; Sentenac, A.; Fromageot, P. Role of deoxyribonucleic acid-ribonucleic acid hybrids in eukaryotes. Synthetic ribo- and deoxyribopolynucleotides as template for yeast ribonucleic acid polymerase B (or II). J. Biol. Chem. 1974, 249, 5978–5983. [Google Scholar]
- Flores, R.; Owens, R.A.; Taylor, J. Pathogenesis by subviral agents: Viroids and hepatitis delta virus. Curr. Opin. Virol. 2016, 17, 87–94. [Google Scholar] [CrossRef]
- Navarro, J.A.; Vera, A.; Flores, R. A chloroplastic RNA polymerase resistant to tagetitoxin is involved in replication of avocado sunblotch viroid. Virology 2000, 268, 218–225. [Google Scholar] [CrossRef] [Green Version]
- Warrilow, D.; Symons, R.H. Citrus exocortis viroid RNA is associated with the largest subunit of RNA polymerase II in tomato in vivo. Arch. Virol. 1999, 144, 2367–2375. [Google Scholar] [CrossRef]
- Muhlbach, H.P.; Sanger, H.L. Viroid replication is inhibited by alpha-amanitin. Nature 1979, 278, 185–188. [Google Scholar] [CrossRef]
- Flores, R.; Semancik, J.S. Properties of a cell-free system for synthesis of citrus exocortis viroid. Proc. Natl. Acad. Sci. USA 1982, 79, 6285–6288. [Google Scholar] [CrossRef] [Green Version]
- Schindler, I.M.; Mühlbach, H.P. Involvement of nuclear DNA-dependent RNA polymerases in potato spindle tuber viroid replication: A reevaluation. Plant Sci. 1992, 84, 221–229. [Google Scholar] [CrossRef]
- Rackwitz, H.R.; Rohde, W.; Sanger, H.L. DNA-dependent RNA polymerase II of plant origin transcribes viroid RNA into full-length copies. Nature 1981, 291, 297–301. [Google Scholar] [CrossRef] [PubMed]
- Yamaguchi, Y.; Filipovska, J.; Yano, K.; Furuya, A.; Inukai, N.; Narita, T.; Wada, T.; Sugimoto, S.; Konarska, M.M.; Handa, H. Stimulation of RNA polymerase II elongation by hepatitis delta antigen. Science 2001, 293, 124–127. [Google Scholar] [CrossRef] [PubMed]
- Fu, T.B.; Taylor, J. The RNAs of hepatitis delta virus are copied by RNA polymerase II in nuclear homogenates. J. Virol. 1993, 67, 6965–6972. [Google Scholar] [CrossRef] [Green Version]
- Chang, J.; Nie, X.; Chang, H.E.; Han, Z.; Taylor, J. Transcription of hepatitis delta virus RNA by RNA polymerase II. J. Virol. 2008, 82, 1118–1127. [Google Scholar] [CrossRef] [Green Version]
- Greco-Stewart, V.S.; Schissel, E.; Pelchat, M. The hepatitis delta virus RNA genome interacts with the human RNA polymerases I and III. Virology 2009, 386, 12–15. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.J.; Macnaughton, T.; Gao, L.; Lai, M.M. RNA-templated replication of hepatitis delta virus: Genomic and antigenomic RNAs associate with different nuclear bodies. J. Virol. 2006, 80, 6478–6486. [Google Scholar] [CrossRef] [Green Version]
- Macnaughton, T.B.; Shi, S.T.; Modahl, L.E.; Lai, M.M. Rolling circle replication of hepatitis delta virus RNA is carried out by two different cellular RNA polymerases. J. Virol. 2002, 76, 3920–3927. [Google Scholar] [CrossRef] [Green Version]
- Modahl, L.E.; Macnaughton, T.B.; Zhu, N.; Johnson, D.L.; Lai, M.M. RNA-dependent replication and transcription of hepatitis delta virus RNA involve distinct cellular RNA polymerases. Mol. Cell. Biol. 2000, 20, 3030–3039. [Google Scholar] [CrossRef] [Green Version]
- Tseng, C.H.; Jeng, K.S.; Lai, M.M. Transcription of subgenomic mRNA of hepatitis delta virus requires a modified hepatitis delta antigen that is distinct from antigenomic RNA synthesis. J. Virol. 2008, 82, 9409–9416. [Google Scholar] [CrossRef] [Green Version]
- Taylor, J.M. Chapter 3. Replication of the hepatitis delta virus RNA genome. Adv. Virus Res. 2009, 74, 103–121. [Google Scholar] [PubMed]
- Taylor, J.M. Hepatitis D virus replication. Cold Spring Harb. Perspect. Med. 2015, 5, a021568. [Google Scholar] [CrossRef] [Green Version]
- Wassarman, K.; Saecker, R. Synthesis-mediated release of a small RNA inhibitor of RNA polymerase. Science 2006, 314, 1601–1603. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wagner, S.D.; Yakovchuk, P.; Gilman, B.; Ponicsan, S.L.; Drullinger, L.F.; Kugel, J.F.; Goodrich, J.A. RNA polymerase II acts as an RNA-dependent RNA polymerase to extend and destabilize a non-coding RNA. EMBO J. 2013, 32, 781–790. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kennedy, W.P.; Momand, J.R.; Yin, Y.W. Mechanism for de novo RNA synthesis and initiating nucleotide specificity by t7 RNA polymerase. J. Mol. Biol. 2007, 370, 256–268. [Google Scholar] [CrossRef] [PubMed]
- Lehmann, E.; Brueckner, F.; Cramer, P. Molecular basis of RNA-dependent RNA polymerase II activity. Nature 2007, 450, 445–449. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Beard, M.R.; MacNaughton, T.B.; Gowans, E.J. Identification and characterization of a hepatitis delta virus RNA transcriptional promoter. J. Virol. 1996, 70, 4986–4995. [Google Scholar] [CrossRef] [Green Version]
- Filipovska, J.; Konarska, M.M. Specific HDV RNA-templated transcription by pol II in vitro. RNA 2000, 6, 41–54. [Google Scholar] [CrossRef] [Green Version]
- Gudima, S.O.; Chang, J.; Taylor, J.M. Reconstitution in cultured cells of replicating HDV RNA from pairs of less than full-length RNAs. RNA 2005, 11, 90–98. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Qu, J.; Ji, S.; Wallace, A.J.; Wu, J.; Li, Y.; Gopalan, V.; Ding, B. A land plant-specific transcription factor directly enhances transcription of a pathogenic noncoding RNA template by DNA-dependent RNA polymerase II. Plant Cell 2016, 28, 1094–1107. [Google Scholar] [CrossRef] [Green Version]
- Jiang, J.; Smith, H.N.; Ren, D.; Dissanayaka Mudiyanselage, S.D.; Dawe, A.L.; Wang, L.; Wang, Y. Potato spindle tuber viroid modulates its replication through a direct interaction with a splicing regulator. J. Virol. 2018, 92, e01004-18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kolonko, N.; Bannach, O.; Aschermann, K.; Hu, K.H.; Moors, M.; Schmitz, M.; Steger, G.; Riesner, D. Transcription of potato spindle tuber viroid by RNA polymerase II starts in the left terminal loop. Virology 2006, 347, 392–404. [Google Scholar] [CrossRef] [PubMed]
- Bojic, T.; Beeharry, Y.; Zhang, D.J.; Pelchat, M. Tomato RNA polymerase II interacts with the rod-like conformation of the left terminal domain of the potato spindle tuber viroid positive RNA genome. J. Gen. Virol. 2012, 93 (Pt 7), 1591–1600. [Google Scholar] [CrossRef] [Green Version]
- Zhong, X.; Archual, A.J.; Amin, A.A.; Ding, B. A genomic map of viroid RNA motifs critical for replication and systemic trafficking. Plant Cell 2008, 20, 35–47. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qi, Y.; Ding, B. Replication of Potato spindle tuber viroid in cultured cells of tobacco and Nicotiana benthamiana: The role of specific nucleotides in determining replication levels for host adaptation. Virology 2002, 302, 445–456. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Branch, A.D.; Robertson, H.D.; Dickson, E. Longer-than-unit-length viroid minus strands are present in RNA from infected plants. Proc. Natl. Acad. Sci. USA 1981, 78, 6381–6385. [Google Scholar] [CrossRef] [Green Version]
- Branch, A.D.; Robertson, H.D. A replication cycle for viroids and other small infectious RNA’s. Science 1984, 223, 450–455. [Google Scholar] [CrossRef]
- Branch, A.D.; Benenfeld, B.J.; Robertson, H.D. Evidence for a single rolling circle in the replication of potato spindle tuber viroid. Proc. Natl. Acad. Sci. USA 1988, 85, 9128–9132. [Google Scholar] [CrossRef] [Green Version]
- Wu, F.H.; Shen, S.C.; Lee, L.Y.; Lee, S.H.; Chan, M.T.; Lin, C.S. Tape-Arabidopsis Sandwich—A simpler Arabidopsis protoplast isolation method. Plant Methods 2009, 5, 16. [Google Scholar] [CrossRef] [Green Version]
- Jiang, J.; Ma, J.; Liu, B.; Wang, Y. Combining a simple method for DNA/RNA/Protein co-purification and Arabidopsis protoplast assay to facilitate viroid research. Viruses 2019, 11, 324. [Google Scholar] [CrossRef] [Green Version]
- Itaya, A.; Zhong, X.; Bundschuh, R.; Qi, Y.; Wang, Y.; Takeda, R.; Harris, A.R.; Molina, C.; Nelson, R.S.; Ding, B. A structured viroid RNA serves as a substrate for dicer-like cleavage to produce biologically active small RNAs but is resistant to RNA-induced silencing complex-mediated degradation. J. Virol. 2007, 81, 2980–2994. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hu, Y.; Feldstein, P.A.; Hammond, J.; Hammond, R.W.; Bottino, P.J.; Owens, R.A. Destabilization of potato spindle tuber viroid by mutations in the left terminal loop. J. Gen. Virol. 1997, 78 (Pt 6), 1199–1206. [Google Scholar] [CrossRef] [Green Version]
- Kang, T.J.; Yang, M.S. Rapid and reliable extraction of genomic DNA from various wild-type and transgenic plants. BMC Biotechnol. 2004, 4, 20. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jendrisak, J.J.; Burgess, R.R. A new method for the large-scale purification of wheat germ DNA-dependent RNA polymerase II. Biochemistry 1975, 14, 4639–4645. [Google Scholar] [CrossRef] [PubMed]
- Beaudry, D.; Perreault, J.P. An efficient strategy for the synthesis of circular RNA molecules. Nucleic Acids Res. 1995, 23, 3064–3066. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhong, X.; Leontis, N.; Qian, S.; Itaya, A.; Qi, Y.; Boris-Lawrie, K.; Ding, B. Tertiary structural and functional analyses of a viroid RNA motif by isostericity matrix and mutagenesis reveal its essential role in replication. J. Virol. 2006, 80, 8566–8581. [Google Scholar] [CrossRef] [Green Version]
- Onodera, Y.; Nakagawa, K.; Haag, J.R.; Pikaard, D.; Mikami, T.; Ream, T.; Ito, Y.; Pikaard, C.S. Sex-biased lethality or transmission of defective transcription machinery in Arabidopsis. Genetics 2008, 180, 207–218. [Google Scholar] [CrossRef] [Green Version]
- Weil, P.A.; Luse, D.S.; Segall, J.; Roeder, R.G. Selective and accurate initiation of transcription at the Ad2 major late promotor in a soluble system dependent on purified RNA polymerase II and DNA. Cell 1979, 18, 469–484. [Google Scholar] [CrossRef]
- Hammond, C.I.; Holland, M.J. Purification of yeast RNA polymerases using heparin agarose affinity chromatography. Transcriptional properties of the purified enzymes on defined templates. J. Biol. Chem. 1983, 258, 3230–3241. [Google Scholar]
- McClements, W.L.; Kaesberg, P. Size and secondary structure of potato spindle tuber viroid. Virology 1977, 76, 477–484. [Google Scholar] [CrossRef]
- Owens, R.A.; Erbe, E.; Hadidi, A.; Steere, R.L.; Diener, T.O. Separation and infectivity of circular and linear forms of potato spindle tuber viroid. Proc. Natl. Acad. Sci. USA 1977, 74, 3859–3863. [Google Scholar] [CrossRef] [Green Version]
- Hadidi, A.; Hashimoto, J.; Diener, T.O. Potato spindle tuber viroid-specific double-stranded RNA in extracts from infected tomato leaves. Ann. Inst. Pasteur Virol. 1982, 133, 15–31. [Google Scholar] [CrossRef]
- Ishikawa, M.; Meshi, T.; Ohno, T.; Okada, Y.; Sano, T.; Ueda, I.; Shikata, E. A revised replication cycle for viroids: The role of longer than unit length RNA in viroid replication. Mol. Gen. Genet. 1984, 196, 421–428. [Google Scholar] [CrossRef] [PubMed]
- Grasser, M.; Kane, C.M.; Merkle, T.; Melzer, M.; Emmersen, J.; Grasser, K.D. Transcript elongation factor TFIIS is involved in arabidopsis seed dormancy. J. Mol. Biol. 2009, 386, 598–611. [Google Scholar] [CrossRef] [PubMed]
- Antosz, W.; Deforges, J.; Begcy, K.; Bruckmann, A.; Poirier, Y.; Dresselhaus, T.; Grasser, K.D. Critical role of transcript cleavage in Arabidopsis RNA polymerase II transcriptional elongation. Plant Cell 2020. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Al Khateeb, W.M.; Sher, A.A.; Marcus, J.M.; Schroeder, D.F. UVSSA, UBP12, and RDO2/TFIIS contribute to Arabidopsis UV tolerance. Front. Plant Sci. 2019, 10, 516. [Google Scholar] [CrossRef]
- Zhong, X.; Tao, X.; Stombaugh, J.; Leontis, N.; Ding, B. Tertiary structure and function of an RNA motif required for plant vascular entry to initiate systemic trafficking. EMBO J. 2007, 26, 3836–3846. [Google Scholar] [CrossRef]
- Kalantidis, K.; Denti, M.A.; Tzortzakaki, S.; Marinou, E.; Tabler, M.; Tsagris, M. Virp1 is a host protein with a major role in Potato spindle tuber viroid infection in Nicotiana plants. J. Virol. 2007, 81, 12872–12880. [Google Scholar] [CrossRef] [Green Version]
- Ding, B. The biology of viroid-host interactions. Annu. Rev. Phytopathol. 2009, 47, 105–131. [Google Scholar] [CrossRef]
- Dissanayaka Mudiyanselage, S.D.; Qu, J.; Tian, N.; Jiang, J.; Wang, Y. Potato spindle tuber viroid RNA-templated transcription: Factors and regulation. Viruses 2018, 10, 503. [Google Scholar] [CrossRef] [Green Version]
- Flores, R.; Gago-Zachert, S.; Serra, P.; Sanjuan, R.; Elena, S.F. Viroids: Survivors from the RNA world? Annu. Rev. Microbiol. 2014, 68, 395–414. [Google Scholar] [CrossRef] [PubMed] [Green Version]





© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dissanayaka Mudiyanselage, S.D.; Wang, Y. Evidence Supporting That RNA Polymerase II Catalyzes De Novo Transcription Using Potato Spindle Tuber Viroid Circular RNA Templates. Viruses 2020, 12, 371. https://doi.org/10.3390/v12040371
Dissanayaka Mudiyanselage SD, Wang Y. Evidence Supporting That RNA Polymerase II Catalyzes De Novo Transcription Using Potato Spindle Tuber Viroid Circular RNA Templates. Viruses. 2020; 12(4):371. https://doi.org/10.3390/v12040371
Chicago/Turabian StyleDissanayaka Mudiyanselage, Shachinthaka D., and Ying Wang. 2020. "Evidence Supporting That RNA Polymerase II Catalyzes De Novo Transcription Using Potato Spindle Tuber Viroid Circular RNA Templates" Viruses 12, no. 4: 371. https://doi.org/10.3390/v12040371