Distribution and Phylogeny of Erythrocytic Necrosis Virus (ENV) in Salmon Suggests Marine Origin

 , , ,
, , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Fish Sampling
2.2. Data Collection
2.3. Metatranscriptomic Sequencing and Bioinformatics
2.4. Spatial Epidemiology Analysis
3. Results
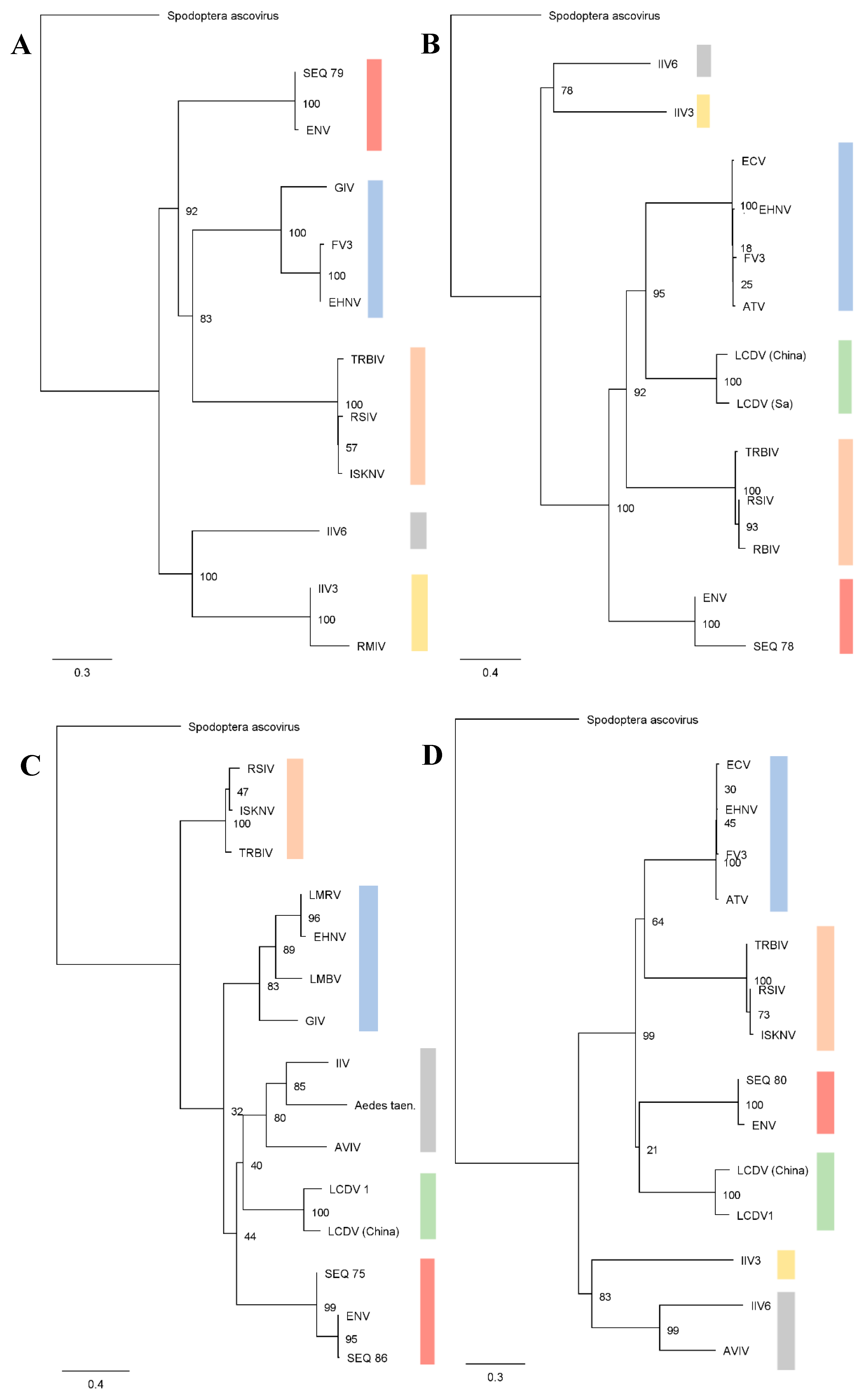
3.1. Genetic Characterization
3.2. Spatial Epidemiology
3.3. Seasonal and Yearly Variation
4. Discussion
Implications
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Hershberger, P.K.; Elder, N.E.; Grady, C.A.; Gregg, J.L.; Pacheco, C.A.; Greene, C.; Rice, C.; Meyers, T.R. Prevalence of Viral Erythrocytic Necrosis in Pacific Herring and Epizootics in Skagit Bay, Puget Sound, Washington. J. Aquat. Anim. Health 2009, 21, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Emmenegger, E.J.; Glenn, J.A.; Winton, J.R.; Batts, W.N.; Gregg, J.L.; Hershberger, P.K. Molecular Identification of Erythrocytic Necrosis Virus (ENV) from the Blood of Pacific Herring (Clupea pallasii). Vet. Microbiol. 2014, 174, 16–26. [Google Scholar] [CrossRef] [PubMed]
- Winton, J.R.; Hershberger, P.K. Viral Erythrocytic Necrosis: Chapter 2.2.7. In FHS Blue Book: Suggested Procedures for the Detection and Identification of Certain Finfish and Shellfish Pathogens; American Fisheries Society: Bethesda, MA, USA, 2014. [Google Scholar]
- Purcell, M.K.; Pearman-Gillman, S.; Thompson, R.L.; Gregg, J.L.; Hart, L.M.; Winton, J.R.; Emmenegger, E.J.; Hershberger, P.K. Identification of the Major Capsid Protein of Erythrocytic Necrosis Virus (ENV) and Development of Quantitative Real-Time PCR Assays for Quantification of ENV DNA. J. Vet. Diagn. Invest. 2016, 28, 382–391. [Google Scholar] [CrossRef] [PubMed]
- Evelyn, T.P.T.; Traxler, G.S. Viral Erythrocytic Necrosis: Natural Occurrence in Pacific Salmon and Experimental Transmission. J. Fish. Res. Board Can. 1978, 35, 903–907. [Google Scholar] [CrossRef]
- Rohovec, J.S.; Amandi, A. Incidence of Viral Erythrocytic Necrosis among Hatchery Reared Salmonids of Oregon. Fish Pathol. 1981, 15, 135–141. [Google Scholar] [CrossRef] [Green Version]
- Haney, D.C.; Hursh, D.A.; Mix, M.C.; Winton, J.R. Physiological and Hematological Changes in Chum Salmon Artificially Infected with Erythrocytic Necrosis Virus. J. Aquat. Anim. Health 1992, 4, 48–57. [Google Scholar] [CrossRef]
- MacMillian, J.R.; Mulcahy, D. Artificial Transmission to and Susceptibility of Puget Sound Fish to Viral Erythrocytic Necrosis (VEN). J. Fish. Res. Board Can. 1979, 36, 1097–1101. [Google Scholar] [CrossRef]
- Eaton, W.D. Artificial Transmission of Erythrocytic Necrosis Virus (ENV) from Pacific Herring in Alaska to Chum, Sockeye, and Pink Salmon. J. Appl. Ichthyol. 1990, 6, 136–141. [Google Scholar] [CrossRef]
- International Committee on Taxonomy of Viruses; King, A.M.Q.; International Union of Microbiological Societies; Virology Division. Virus Taxonomy: Classification and Nomenclature of Viruses: Ninth Report of the International Committee on Taxonomy of Viruses; Academic Press: London, UK, 2012; ISBN 978-0-12-384684-6. [Google Scholar]
- Huang, S.M.; Tu, C.; Tseng, C.H.; Huang, C.C.; Chou, C.-C.; Kuo, H.-C.; Chang, S.-K. Genetic Analysis of Fish Iridoviruses Isolated in Taiwan during 2001–2009. Arch. Virol. 2011, 156, 1505–1515. [Google Scholar] [CrossRef]
- Wang, C.S.; Shih, H.H.; Ku, C.C.; Chen, S.N. Studies on Epizootic Iridovirus Infection Among Red Sea Bream, Pagrus major (Temminck & Schlegel), Cultured in Taiwan. J. Fish Dis. 2003, 26, 127–133. [Google Scholar] [PubMed]
- Liu, H.I.; Lin, Y.C.; Chiou, P.P.; Chou, H.-Y. Temperature-Dependent Pathogenicity of Grouper Iridovirus of Taiwan (TGIV). J. Mar. Sci. Technol. 2016, 24, 637–644. [Google Scholar]
- Wang, Y.Q.; Lü, L.; Weng, S.P.; Huang, J.N.; Chan, S.-M.; He, J.G. Molecular Epidemiology and Phylogenetic Analysis of a Marine Fish Infectious Spleen and Kidney Necrosis Virus-like (ISKNV-like) Virus. Arch. Virol. 2007, 152, 763–773. [Google Scholar] [CrossRef] [PubMed]
- Meyers, T.R.; Hauck, A.K.; Blankenbeckler, W.D.; Minicucci, T. First report of Viral Erythrocytic Necrosis in Alaska, USA, Associated with Epizootic Mortality in Pacific Herring, Clupea harengus pallasi (Valenciennes). J. Fish Dis. 1986, 9, 479–491. [Google Scholar] [CrossRef]
- Nekouei, O.; Vanderstichel, R.; Ming, T.; Kaukinen, K.H.; Thakur, K.; Tabata, A.; Laurin, E.; Tucker, S.; Beacham, T.D.; Miller, K.M. Detection and Assessment of the Distribution of Infectious Agents in Juvenile Fraser River Sockeye Salmon, Canada, in 2012 and 2013. Front. Microbiol. 2018, 9. [Google Scholar] [CrossRef] [PubMed]
- Tucker, S.; Li, S.; Kaukinen, K.H.; Patterson, D.A.; Miller, K.M. Distinct Seasonal Infectious Agent Profiles in Life-history Variants of Juvenile Fraser River Chinook Salmon: An Application of High-Throughput Genomic Screening. PLoS ONE 2018, 13, e0195472. [Google Scholar] [CrossRef] [PubMed]
- Thakur, K.K.; Vanderstichel, R.; Li, S.; Laurin, E.; Tucker, S.; Neville, C.; Tabata, A.; Miller, K.M. A Comparison of Infectious Agents Between Hatchery-Enhanced and Wild Out-Migrating Juvenile Chinook Salmon (Oncorhynchus tshawytscha) from Cowichan River, British Columbia. FACETS 2018. [Google Scholar] [CrossRef]
- Laurin, E.; Jaramillo, D.; Vanderstichel, R.; Ferguson, H.; Kaukinen, K.H.; Schulze, A.D.; Keith, I.R.; Gardner, I.A.; Miller, K.M. Histopathological and Novel High-Throughput Molecular Monitoring Data from Farmed Salmon (Salmo salar and Oncorhynchus spp.) in British Columbia, Canada, from 2011–2013. Aquaculture 2019, 499, 220–234. [Google Scholar] [CrossRef]
- Teffer, A.K.; Bass, A.L.; Miller, K.M.; Patterson, D.A.; Juanes, F.; Hinch, S.G. Infections, Fisheries Capture, Temperature, and Host Responses: Multistressor Influences on Survival and Behaviour of Adult Chinook Salmon. Can. J. Fish. Aquat. Sci. 2018, 75, 2069–2083. [Google Scholar] [CrossRef]
- DFO. Stock Assessment Report on Pacific Herring in British Columbia. Canadian Science Advisory Secretariat Science Advisory Report 2009/059. Available online: http://www.dfo-mpo.gc.ca/Library/338390.pdf (accessed on 17 April 2019).
- Varpe, Ø.; Fiksen, Ø.; Slotte, A. Meta-Ecosystems and Biological Energy Transport from Ocean to Coast: The Ecological Importance of Herring Migration. Oecologia 2005, 146, 443. [Google Scholar] [CrossRef] [PubMed]
- Pacific Salmon Commission. 2010 Annual Report of Catches and Escapements; Report TCChinook (11)-2; Pacific Salmon Commission: Vancouver, BC, Canada, 2011. [Google Scholar]
- Government of Canada. Summary of COSEWIC Wildlife Species Assessments. November 2018. Available online: https://www.canada.ca/en/environment-climate-change/services/committee-status-endangered-wildlife/assessments/wildlife-species-assessment-summary-nov-2018.html (accessed on 28 January 2019).
- Miller, K.M.; Teffer, A.; Tucker, S.; Li, S.; Schulze, A.D.; Trudel, M.; Juanes, F.; Tabata, A.; Kaukinen, K.H.; Ginther, N.G.; et al. Infectious Disease, Shifting Climates, and Opportunistic Predators: Cumulative Factors Potentially Impacting Wild Salmon Declines. Evol. Appl. 2014, 7, 812–855. [Google Scholar] [CrossRef] [PubMed]
- Miller, K.M.; Gardner, I.A.; Vanderstichel, R.; Burnley, T.; Schulze, A.D.; Li, S.; Tabata, A.; Kaukinen, K.H.; Ming, T.J.; Ginther, N.G. Report on the Performance Evaluation of the Fluidigm BioMark Platform for High Throughput Microbe Monitoring in Salmon; Fisheries and Oceans Canada: Ottawa, ON, Canada, 2016.
- Di Cicco, E.; Ferguson, H.W.; Kaukinen, K.H.; Schulze, A.D.; Li, S.; Tabata, A.; Günther, O.P.; Mordecai, G.; Suttle, C.A.; Miller, K.M. The Same Strain of Piscine Orthoreovirus (PRV-1) is Involved in the Development of Different, but Related, Diseases in Atlantic and Pacific Salmon in British Columbia. FACETS 2018, 3, 599–641. [Google Scholar] [CrossRef]
- Bass, A.L.; Hinch, S.G.; Teffer, A.K.; Patterson, D.A.; Miller, K.M. A Survey of Microparasites Present in Adult Migrating Chinook Salmon (Oncorhynchus tshawytscha) in South-Western British Columbia Determined by High-Throughput Quantitative Polymerase Chain Reaction. J. Fish Dis. 2017, 40, 453–477. [Google Scholar] [CrossRef]
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A Flexible Trimmer for Illumina Sequence Data. Bioinforma. Oxf. Engl. 2014, 30, 2114–2120. [Google Scholar] [CrossRef] [PubMed]
- Davidson, W.S.; Koop, B.F.; Jones, S.J.; Iturra, P.; Vidal, R.; Maass, A.; Jonassen, I.; Lien, S.; Omholt, S.W. Sequencing the Genome of the Atlantic Salmon (Salmo salar). Genome Biol. 2010, 11, 403. [Google Scholar] [PubMed]
- Li, H.; Durbin, R. Fast and Accurate Short Read Alignment with Burrows-Wheeler Transform. Bioinform. Oxf. Engl. 2009, 25, 1754–1760. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Handsaker, B.; Wysoker, A.; Fennell, T.; Ruan, J.; Homer, N.; Marth, G.; Abecasis, G.; Durbin, R. 1000 Genome Project Data Processing Subgroup the Sequence Alignment/Map Format and SAMtools. Bioinforma. Oxf. Engl. 2009, 25, 2078–2079. [Google Scholar] [CrossRef] [PubMed]
- Bankevich, A.; Nurk, S.; Antipov, D.; Gurevich, A.A.; Dvorkin, M.; Kulikov, A.S.; Lesin, V.M.; Nikolenko, S.I.; Pham, S.; Prjibelski, A.D.; et al. SPAdes: A New Genome Assembly Algorithm and its Applications to Single-Cell Sequencing. J. Comput. Biol. J. Comput. Mol. Cell Biol. 2012, 19, 455–477. [Google Scholar] [CrossRef] [PubMed]
- Buchfink, B.; Xie, C.; Huson, D.H. Fast and Sensitive Protein Alignment using DIAMOND. Nat. Methods 2015, 12, 59–60. [Google Scholar] [CrossRef] [PubMed]
- Caporaso, J.G.; Kuczynski, J.; Stombaugh, J.; Bittinger, K.; Bushman, F.D.; Costello, E.K.; Fierer, N.; Peña, A.G.; Goodrich, J.K.; Gordon, J.I.; et al. QIIME Allows Analysis of High-Throughput Community Sequencing Data. Nat. Methods 2010, 7, 335–336. [Google Scholar] [CrossRef]
- Borodovsky, M.; Mills, R.; Besemer, J.; Lomsadze, A. Prokaryotic Gene Prediction Using GeneMark and GeneMark.hmm. Curr. Protoc. Bioinform. 2003, 4, 4.5.1–4.5.16. [Google Scholar] [CrossRef] [PubMed]
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic Local Alignment Search Tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef]
- Yutin, N.; Wolf, Y.I.; Raoult, D.; Koonin, E.V. Eukaryotic Large Nucleo-Cytoplasmic DNA Viruses: Clusters of Orthologous Genes and Reconstruction of Viral Genome Evolution. Virol. J. 2009, 6, 223. [Google Scholar] [CrossRef] [PubMed]
- Kearse, M.; Moir, R.; Wilson, A.; Stones-Havas, S.; Cheung, M.; Sturrock, S.; Buxton, S.; Cooper, A.; Markowitz, S.; Duran, C.; et al. Geneious Basic: An Integrated and Extendable Desktop Software Platform for the Organization and Analysis of Sequence Data. Bioinformatics 2012, 28, 1647–1649. [Google Scholar] [CrossRef] [PubMed]
- Guindon, S.; Dufayard, J.-F.; Lefort, V.; Anisimova, M.; Hordijk, W.; Gascuel, O. New Algorithms and Methods to Estimate Maximum-Likelihood Phylogenies: Assessing the Performance of PhyML 3.0. Syst. Biol. 2010, 59, 307–321. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thompson, J.D.; Higgins, D.G.; Gibson, T.J. CLUSTAL W: Improving the Sensitivity of Progressive Multiple Sequence Alignment Through Sequence Weighting, Position-Specific Gap Penalties and Weight Matrix Choice. Nucleic Acids Res. 1994, 22, 4673–4680. [Google Scholar] [CrossRef] [PubMed]
- R Core Team R: A Language and Environment for Statistical Computing. Ed. Found. Stat. Comput. Vienna Austria 2013.
- Pebesma, E.J. Multivariable Geostatistics in S: The gstat Package. Comput. Geosci. 2004, 30, 683–691. [Google Scholar] [CrossRef]
- Heuvelink, G.B.M. Spatio-Temporal Interpolation Using gstat by Benedikt Gräler, Edzer Pebesma and Gerard Heuvelink. R J. 2016, 8, 204–218. [Google Scholar]
- Kahle, D.; Wickham, H. ggmap: Spatial Visualization with ggplot2. R J. 2013, 5, 144. [Google Scholar] [CrossRef]
- Mangiafico, S. rcompanion: Functions to Support Extension Education Program Evaluation [R Statistical Package]. 2016. Available online: https://cran.r-project.org/web/packages/rcompanion/ (accessed on 17 April 2018).
- Bates, D.; Mächler, M.; Bolker, B.; Walker, S. Fitting Linear Mixed-Effects Models Using lme4. J. Stat. Softw. 2015, 1. [Google Scholar] [CrossRef]
- Hothorn, T.; Bretz, F.; Westfall, P. Simultaneous Inference in General Parametric Models. Biom. J. 2008, 50, 346–363. [Google Scholar] [CrossRef] [Green Version]
- Teng, Y.; Hou, Z.; Gong, J.; Liu, H.; Xie, X.; Zhang, L.; Chen, X.; Qin, Q.W. Whole-Genome Transcriptional Profiles of a Novel Marine Fish Iridovirus, Singapore Grouper Iridovirus (SGIV) in Virus-Infected Grouper Spleen Cell Cultures and in Orange-Spotted Grouper, Epinephulus coioides. Virology 2008, 377, 39–48. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.M.; Wang, F.; Song, W.; Hew, C.L. Temporal and Differential Gene Expression of Singapore Grouper Iridovirus. J. Gen. Virol. 2006, 87, 2907–2915. [Google Scholar] [CrossRef] [PubMed]
- İnce, İ.A.; Özcan, O.; Ilter-Akulke, A.Z.; Scully, E.D.; Özgen, A. Invertebrate Iridoviruses: A Glance Over the Last Decade. Viruses 2018, 10, 161. [Google Scholar] [CrossRef]
- Sample, R.; Bryan, L.; Long, S.; Majji, S.; Hoskins, G.; Sinning, A.; Olivier, J.; Chinchar, V.G. Inhibition of Iridovirus Protein Synthesis and Virus Replication by Antisense Morpholino Oligonucleotides Targeted to the Major Capsid Protein, the 18 kDa Immediate-Early Protein, and a Viral Homolog of RNA Polymerase II. Virology 2007, 358, 311–320. [Google Scholar] [CrossRef] [PubMed]
- Kurobe, T.; MacConnell, E.; Hudson, C.; McDowell, T.S.; Mardones, F.O.; Hedrick, R.P. Iridovirus Infections among Missouri River Sturgeon: Initial Characterization, Transmission, and Evidence for Establishment of a Carrier State. J. Aquat. Anim. Health 2011, 23, 9–18. [Google Scholar] [CrossRef]
- Georgiadis, M.P.; Hedrick, R.P.; Johnson, W.O.; Yun, S.; Gardner, I.A. Risk Factors for Outbreaks of Disease Attributable to White Sturgeon Iridovirus and White Sturgeon Herpesvirus-2 at a Commercial Sturgeon Farm. Am. J. Vet. Res. 2000, 61, 1232–1240. [Google Scholar] [CrossRef]
- Traxler, G.; Bell, G. Pathogens Associated with Impounded Pacific Herring Clupea harengus pallasi, with Emphasis on Viral Erythrocytic Necrosis (VEN) and Atypical Aeromonas salmonicida. Dis. Aquat. Organ. 1988, 5, 93–100. [Google Scholar] [CrossRef]
- Hershberger, P.; Hart, A.; Gregg, J.; Elder, N.; Winton, J. Dynamics of Viral Hemorrhagic Septicemia, Viral Erythrocytic Necrosis and Ichthyophoniasis in Confined Juvenile Pacific Herring Clupea pallasii. Dis. Aquat. Organ. 2006, 70, 201–208. [Google Scholar] [CrossRef]
- Weitkamp, L.A. Marine Distributions of Chinook Salmon from the West Coast of North America Determined by Coded Wire Tag Recoveries. Trans. Am. Fish. Soc. 2010, 139, 147–170. [Google Scholar] [CrossRef]
- Myers, K.W.; Klovach, N.V.; Gritsenko, O.F.; Urawa, S.; Royer, T.C. Stock-Specific Distributions of Asian and North American Salmon in the Open Ocean, Interannual Changes, and Oceanographic Conditions; NPAFC: Vancouver, BC, Canada, 2007. [Google Scholar]
- Hourston, A.S.; Haegele, C.W. Herring on Canada’s Pacific Coast; Department of Fisheries and Oceans: Longueuil, QC, USA, 1980.
- Schroeder, D.C.; Martin, S.J. Deformed wing virus. Virulence 2012, 3, 589–591. [Google Scholar] [CrossRef] [PubMed]
- Bell, Q.R.; Traxler, G.S. First Record of Viral Erythrocytic Necrosis and Ceratomyxa Shasta Noble, 1950 (Myxozoa: Myxosporea) in Feral Pink Salmon (Oncorhynchus gorbuscha Walbaum). J. Wildl. Dis. 1985, 21, 169–171. [Google Scholar] [CrossRef] [PubMed]
- Peterson, W.; Morgan, C.; Casillas, E.; Peterson, J.; Fisher, J.; W Ferguson, J. Ocean Ecosystem Indicators of Salmon Marine Survival in the Northern California Current. 2019. Available online: https://www.fwspubs.org/doi/suppl/10.3996/042010-JFWM-009/suppl_file/10.3996_042010-jfwm-009.s6.pdf (accessed on 17 April 2019).
- Goniea, T.; Bjornn, T.C.; Peery, C.A.; Bennett, D.H.; Stuehrenberg, L.C. Behavioral Thermoregulation and Slowed Migration by Adult Fall Chinook Salmon in Response to High Columbia River Water Temperatures. Trans. Am. Fish. Soc. 2006, 135, 408–419. [Google Scholar] [CrossRef] [Green Version]
- Beacham, T.D.; Murray, C.B. Effect of Female Size, Egg Size, and Water Temperature on Developmental Biology of Chum Salmon (Oncorhynchus keta) from the Nitinat River, British Columbia. Can. J. Fish. Aquat. Sci. 1985, 42, 1755–1765. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Reference Sequence | Sequence ID | % Identity (Nucleotide) | % Identity (Amino Acid) | Sequence Alignment Length (AA) |
|---|---|---|---|---|
| ATPase KJ730210.1 (partial) | 80 (Aq.) | 99.9 | 100 | 849 |
| 46 (Wild) | 99.6 | 98.7 | 74 | |
| 57 (Wild) | 99.5 | 98.5 | 67 | |
| 54 (Wild) | 100 | 100 | 77 | |
| DNA-dependent DNA polymerase KJ756347.1 (partial) | 73 (Wild) | 99.9 | 98.3 | 59 |
| 79 (Aq.) | 99.8 | 99.7 | 897 | |
| 19 (Wild) | 99.5 | 99.5 | 213 | |
| MCP KT211480.1 (partial, Puget Sound) | 86 (Aq.) | 99.6 | 100 | 1436 |
| 75 (Wild) | 100 | 100 | 175 | |
| DNA-dependent RNA polymerase KJ756346.1 (partial) | 78 (Aq.) | 99.9 | 96.8 | 972 |
| Abbreviation | Accession Numbers | Taxon | |||
|---|---|---|---|---|---|
| DNA-Dependent DNA Polymerase | DNA-Dependent RNA Polymerase | MCP | ATPase | ||
| NA | AAC54632.1 | YP_762407.1 | NC_008361.1 | NC_008361.1 | Spodoptera ascovirus |
| ENV | AIQ77732.1 | AIQ77731.1 | KT211480.1 | AIN76233.1 | Erythrocytic necrosis |
| Megalocytivirus | |||||
| RSIV | BAA28669.1 | BAK14252.1 | AB109371.1 | BAK14298.1 | Red Sea Bream iridovirus |
| ISKNV | CAZ73994.1 | AF370008.1 | NP_612331.1 | Infectious spleen and kidney necrosis virus | |
| TRBIV | ADE34365.1 | ADE34378.1 | AY590687.2 | ADE34443.1 | Turbot reddish body iridovirus |
| RBIV | AAT71848.1 | Rock Bream iridovirus | |||
| Ranavirus | |||||
| FV3 | NC_005946.1 | ASH99239.1 | AHM26101.1 | Frog virus 3 | |
| EHNV | ACO25234.1 | YP_009182042.1 | AY187045.1 | YP_009182084.1 | Epizootic haematopoietic necrosis virus |
| GIV | AY666015.1 | KX284838.1 | Grouper iridovirus | ||
| ECV | AMZ05024.1 | YP_006347705.1 | European catfish virus | ||
| ATV | ALN36639.1 | YP_003852.1 | Ambystoma tigrinum virus | ||
| LMRV | KM516719.1 | Lacerta monticola ranavirus | |||
| LMBV | KU507317.1 | Largemouth bass ranavirus | |||
| Lymphocystivirus | |||||
| LCDV(Sa) | YP_009342128.1 | AY823414.1 | Lymphocystis disease virus (various strains) | ||
| LCDV(China) | YP_073534.1 | NC_005902.1 | YP_073585.1 | ||
| LCDV1 | AAX54510.1 | ||||
| Iridovirus | |||||
| IIV6 | NC_003038.1 | AAK82288.1 | NP_149647.1 | Invertebrate iridescent virus 6 | |
| IIV | NC_023615.1 | Invertebrate iridescent virus | |||
| AVIV | NC_024451.1 | NC_024451.1 | Armadillium vulgare iridescent virus | ||
| Aedes taen. | NC_008187.1 | Aedes taeniorhynchus iridescent virus | |||
| Chloriridovirus | |||||
| IIV3 | YP_654692.1 | YP_654581.1 | YP_654693.1 | Invertebrate iridescent virus 3 | |
| RMIV | CAC84133.1 | Regular mosquito iridescent virus | |||
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pagowski, V.A.; Mordecai, G.J.; Miller, K.M.; Schulze, A.D.; Kaukinen, K.H.; Ming, T.J.; Li, S.; Teffer, A.K.; Tabata, A.; Suttle, C.A. Distribution and Phylogeny of Erythrocytic Necrosis Virus (ENV) in Salmon Suggests Marine Origin. Viruses 2019, 11, 358. https://doi.org/10.3390/v11040358
Pagowski VA, Mordecai GJ, Miller KM, Schulze AD, Kaukinen KH, Ming TJ, Li S, Teffer AK, Tabata A, Suttle CA. Distribution and Phylogeny of Erythrocytic Necrosis Virus (ENV) in Salmon Suggests Marine Origin. Viruses. 2019; 11(4):358. https://doi.org/10.3390/v11040358
Chicago/Turabian StylePagowski, Veronica A., Gideon J. Mordecai, Kristina M. Miller, Angela D. Schulze, Karia H. Kaukinen, Tobi J. Ming, Shaorong Li, Amy K. Teffer, Amy Tabata, and Curtis A. Suttle. 2019. "Distribution and Phylogeny of Erythrocytic Necrosis Virus (ENV) in Salmon Suggests Marine Origin" Viruses 11, no. 4: 358. https://doi.org/10.3390/v11040358