The Importance of Being Non-Defective: A Mini Review Dedicated to the Memory of Jan Svoboda


Department of Molecular Medicine, The Scripps Research Institute, La Jolla, CA 92037, USA
Viruses 2019, 11(1), 80; https://doi.org/10.3390/v11010080
Submission received: 27 December 2018
/
Revised: 15 January 2019
/
Accepted: 16 January 2019
/
Published: 18 January 2019
(This article belongs to the Special Issue Homage to Jan Svoboda: Recent Advances in Topics Pioneered by his Prague School)
{kind=link}
Abstract
:Jan Svoboda triggered investigations on non-defective avian sarcoma viruses. These viruses were a critical factor in the genetic understanding of retroviruses. They provided the single and unique access to the field and facilitated the discovery of the first oncogene src and of the cellular origin of retroviral oncogenes. They continue to be of importance as singularly effective expression vectors that have provided insights into the molecular functions of numerous oncogenes. Combined with the contributions to the validation of the provirus hypothesis, Jan Svoboda’s investigations of non-defective avian sarcoma viruses have shaped a large and important part of retrovirology.
At the beginning of experimental tumor virology stands a new technique: the focus assay for Rous sarcoma virus (RSV) [1]. It transformed tumor virology into a cell culture-based science. This assay provided the technical framework for a strategy adopted from phage research which included a reductionist approach and quantitation rather than description. It also led to an emphasis on work with single cells and single virus particles. These were novel and groundbreaking ideas which were quickly applied to a broad spectrum of tumor viruses, including an increasing number of cancer-causing viruses identified in mammals [2,3,4,5,6,7,8,9,10,11,12,13].
The original focus assay used the Bryan high-titer strain of RSV, a strain that had been selected because it generated high viral titers in chicken tumors from which working stocks were prepared at the time [14,15]. As the focus assay was widely adopted, especially in US labs, the Bryan high-titer strain of RSV gained almost exclusive dominance.
1. Defectiveness in Replication: The Bryan High-Titer Strain as the Prototype
Using the focus assay, Temin followed up on a hypothetical analogy between RSV and lysogenic phage [16]. He studied cell foci transformed by a single particle of the Bryan high-titer strain and discovered foci that did not release a detectable infectious virus [17]. Hanafusa carried out an extensive analysis of this non-producing state, and the conclusion of these studies was that the Bryan high-titer strain of RSV was defective in replication, requiring the assistance of a helper virus to generate infectious progeny [18]. The replication defectiveness of the Bryan high-titer strain also led to the broader conclusion that virus production was not required for oncogenic transformation. However, that extrapolation was soon challenged by electron-microscopic observations that showed the release of virus particles from non-producer cells and the recovery of infectious virus from non-producing cultures [19,20]. Most of the virus synthesized by non-producer cells turned out to carry a novel surface glycoprotein provided by an endogenous avian leukosis virus that acted as a helper virus. The corresponding cell surface receptor encoded by the S1 allele of the Tvb gene [21,22,23] was not present on the chicken “non-producer” cells, making the virus undetectable in the standard assay [24]. However, this subgroup E envelope protein readily mediated entry into cells of the Japanese quail, Coturnix coturnix japonica. The virus released from chicken “non-producer” cells is fully infectious and capable of reproducing in quail cell cultures. Scheele and Hanafusa precisely defined the genetic defect in the Bryan high-titer strain of RSV by showing that the viral genome lacks genetic information for the envelope protein. This information is provided by the helper virus [25].
The Bryan high-titer strain of RSV became the prototype for oncogene-carrying retroviruses. Virtually all of these acutely transforming viruses are replication-defective, because they have lost viral replicative information in exchange for acquiring cell-derived oncogenic sequences.
2. Non-Defective Avian Sarcoma Viruses
The discovery of defectiveness in RSV was an exciting and important event in the evolution of tumor virology. However, there were other strains of avian sarcoma in circulation, notably in Europe. These included the Prague strain of RSV [26], the Carr–Zilber and Schmidt–Ruppin strains [27,28] and the independently isolated avian sarcoma virus referred to as Bratislava 77 (B77) [29]. Several of these strains were studied for their activity in mammals [27,28,30,31,32,33,34]. In contrast to the Bryan high-titer strain, they showed significant ability to induce tumors in rodents and other laboratory animals and to transform mammalian cells in culture. Jan Svoboda was a pioneer in this area. In one of his groundbreaking studies, he showed that rat cells transformed by the Prague strain of RSV failed to release infectious virus. However, co-cultivation of these cells with chicken embryo fibroblasts led to the production of infectious RSV [35,36,37]. This observation on mammalian cells provided more compelling evidence than could be derived from chicken “non-producing” cells for the general conclusion that infectious virus replication was not required for oncogenic transformation. Since no helper virus had to be added to achieve virus replication in the chicken–rat co-cultures, these experiments also suggested that the Prague strain of RSV was non-defective, capable of both replication and transformation. The non-defectiveness of the Prague strain of RSV was soon confirmed in avian cell culture [38]. RNA analysis also showed the Schmidt–Ruppin and Carr–Zilber strains of RSV and the B77 avian sarcoma virus to be non-defective [39,40].
The finding of non-defectiveness in several strains of avian sarcoma virus changed the focus of the field. Non-defective avian sarcoma viruses carry all replicative and transforming functions on the same RNA molecule, allowing the molecular identification and characterization of these functions and their interactions. Since non-defectiveness is a unique attribute of avian sarcoma viruses and has not been observed with any other oncogene-carrying retrovirus, avian sarcoma viruses provided the only key to establishing the foundation of retrovirus genetics. In contrast, replication-defective oncogenic retroviruses can be propagated only with a helper virus, as two-virus systems, greatly complicating genetic analysis.
3. Conditional and Non-Conditional Mutants
The first conditional mutants of a non-defective avian sarcoma virus that are temperature-sensitive for their ability to transform cells in culture were isolated by Kumao Toyoshima. These mutants carried multiple mutations in replicative and oncogenic functions [41,42]. Steven Martin then isolated a temperature-sensitive mutant that affected only oncogenic activity, delivering genetic evidence for a specific viral oncogene, later termed src [43]. The existence of this oncogene could also be deduced from biochemical data. During the replication of cloned non-defective avian sarcoma viruses, derivative viruses are generated that are no longer oncogenic but reproduce and are fully infectious [44]. At high-virus dilution, they can be separated from the parental sarcoma virus and grown independently in the cell culture. They are transformation-defective. The size of their genome is smaller than that of the parental sarcoma virus. They have lost the specific viral gene encoding oncogenic activity [45]. Temperature-sensitive mutants affecting oncogenicity map to the genome section that is deleted in the transformation-defective viruses [46]. Oncogenic and transformation-defective avian sarcoma viruses are isogenic pairs, except for the deletion of the src gene in the non-transforming derivative [44]. Such isogenic pairs provide the opportunity to generate a specific probe for src using subtractive hybridization with DNA transcripts of transforming and transformation-defective genomes. This src probe showed that homologous sequences were present in the genome of normal avian as well as mammalian cells [47]. Other retroviral oncogenes were also identified as cellular genes [48]. The cellular origin of oncogenes was a revolutionary discovery with far-reaching consequences. It showed that the genomes of normal cells encompass genes with oncogenic potential. In the ensuing years, the functional characteristics of these genes illuminated mechanisms of cellular signaling and regulation. This became the core of a genetic interpretation of human cancer [13,49,50,51].
Conditional and non-conditional mutants in replicative genes also provided a first insight into retrovirus gene function and genome structure [52,53,54,55,56]. The group of Peter Duesberg applied RNA fingerprinting to non-conditional and conditional mutants of avian sarcoma viruses and constructed the first map of a retroviral genome [57,58].
4. The Continuing Significance of Oncogenic Retroviruses
The discovery of the cellular origin of oncogenes demoted oncogenic retroviruses from originators to transport vehicles or insertional mutagenic activators of cellular oncogenic information [47,48,59,60]. Many tumor virologists became cell biologists and oncologists, but retroviruses retained significant importance as sources of novel oncogenes, as animal pathogens and as models for the construction of expression vectors.
Replication-defective, acutely oncogenic retroviruses were identified repeatedly in spontaneous tumors in avian and mammalian species and led to the discovery of new oncogenes. Among the avian viruses, an early example is myc, first discovered in avian myelocytomatosis virus MC29 [61,62]. The oncogene jun, identified in the genome of avian sarcoma virus 17, showed the importance of transcriptional regulation in oncogenesis [63,64]. The catalytic isoform of PI 3-kinase drives tumor formation by avian sarcoma virus 16 [65] and plays a major role in human cancer [66]. The genomes of several acutely oncogenic mammalian retroviruses have also yielded important oncogenes, most notably abl, ras, raf, fos, and kit [13].
Avian retroviruses also remain potent pathogens in chickens, causing major losses in the poultry industry. A recent example is the outbreak of epidemics of hemangiomas and myelocytomas induced by subgroup J avian leukosis viruses in China. This subgroup of avian leukosis was first identified in 1991 in Britain [67,68], but the significance of this finding has emerged only recently [69,70,71] as this virus is spreading world-wide [72,73,74]. The evolution of avian leukosis viruses is continuing, and new envelope subgroups are likely to emerge [75]. Since poultry is one of the main sources of animal protein, these viruses are of serious concern for the world food supply.
The genome of the Schmidt–Ruppin strain of RSV has been re-engineered into a widely used and exceptionally efficient vector, referred to as RCAS, that can be used to express high levels of any protein in avian cells [76,77,78,79,80]. RCAS retains the unique features of non-defective avian sarcoma viruses: active replication and spreading to adjacent cells and expression of an inserted gene. The efficiency of the initial DNA transfection that often limits the utility of expression vectors does not affect outcome with RCAS, because this expression vector is a complete retrovirus that replicates and eventually will infect every avian cell in the culture. There is also a replication-defective version of RCAS, derived from the Bryan high-titer strain of RSV. It is dependent on an avian leukosis helper virus which allows easy changes in the envelope protein and thus its host range. The RCAS vector tool has been adapted in a limited way to mammalian cells that are engineered to express the avian retroviral receptor TVA. An RCAS version with a broader host range in mammalian cells uses the envelope gene of a mouse retrovirus [80], but mammalian cells do not support virus replication and spread relies solely on cell replication.
5. Idle Speculation
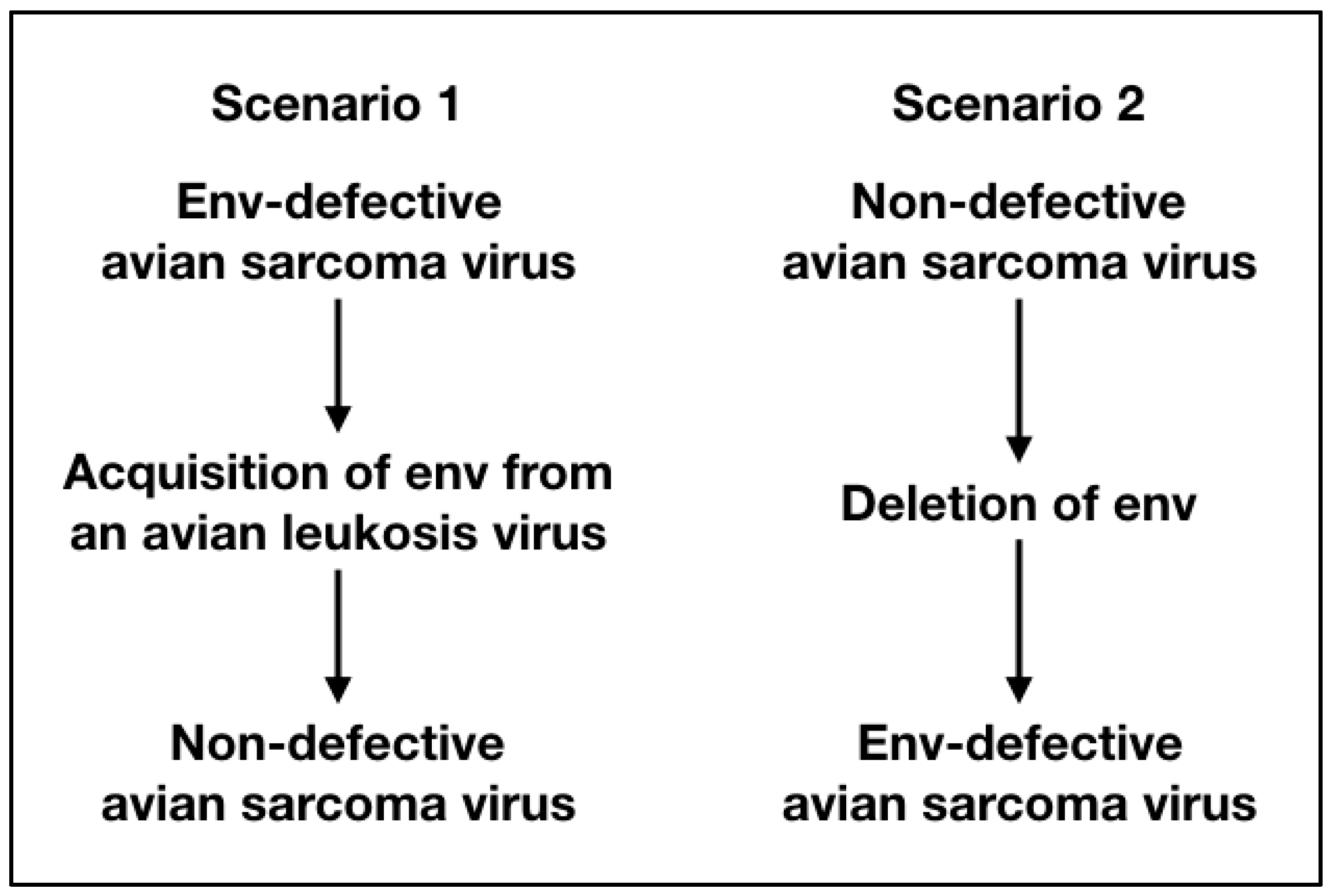
It has long been tempting to speculate on the origin of non-defective avian sarcoma viruses, because they are unique among acutely oncogenic, oncogene-carrying retroviruses [81]. Even among all the RSV strains, they form a separate category. The passage histories of the major RSV strains, Bryan high-titer, Schmidt–Ruppin, Carr–Zilber and Prague strains of RSV, are not known and cannot be reconstructed. Was Peyton Rous’ original isolate replication-defective or non-defective? Figure 1 summarizes these two possible scenarios.
In scenario 1, the original Rous isolate is replication-defective, lacking the envelope gene. It would have to acquire this gene to become non-defective. Although an acquisition is theoretically possible, given the high frequency of recombination between avian retroviruses, such a capture has never been observed. To detect a rare non-defective recombinant, it would be necessary to screen hundreds and possibly thousands of cell clones transformed by a single viral particle. It is unlikely for such a recombinant to become the prevalent component of an RSV stock, as defective viruses with their small genomes tend to have a selective advantage, because they replicate faster. Yet, in the non-defective avian sarcoma viruses, the non-defective component is prevalent.
In scenario 2, the env gene would have been deleted at some time in the passage history of certain RSV strains, including the Bryan high-titer strain. The deletions of replicative genes in avian retroviruses have been observed, so there is precedence for such a genetic change [52,82]. Combined with a selection for high virus titers, the smaller genome of a replication-defective virus could rapidly become the prevalent component during passage.
The sequence analysis of replication-defective RSV29, believed to be an early passage of the original Rous isolate, suggests that this virus is a direct precursor of the Bryan high-titer strain [83]. The conclusion that this also indicates defectiveness of the original isolate is not warranted, as deletion of the env gene could have occurred and could be selected for within the presumably short passage history of RSV29 [83].
These considerations favor scenario 2. It should be noted that a replication-defective version of the src-carrying B77 avian retrovirus has never been identified, even in the earliest passages. This virus appears to have been non-defective at the time of its isolation from a spontaneous chicken sarcoma.
The issue of the defectiveness or non-defectiveness of the original Rous isolate brings up the more important question of the acquisition of cellular oncogenes by retroviruses. There have been reports of “recovered” or “rescued” sarcoma viruses [84,85,86] that arose after infection with a transformation-defective leukosis virus. However, in all these cases, the capture of cellular src can be explained by homologous recombination between viral [85,86] or cellular sequences [84]. These are not de novo acquisitions as they are mediated by residual src sequences in the virus or retroviral sequences persisting from a previous infection adjacent to cellular src. The recombination between viral and cellular genomes resulting in the incorporation of an oncogene in the virus so far has occurred only in vivo. There is no experimental system that can reproducibly generate such virus–cell recombinants.
6. Jan Svoboda, the Catalyst
Jan Svoboda has made important, prescient contributions to the origin and validation of the provirus hypothesis [30,35,87,88,89,90,91,92]. However, he has also triggered the development that gave the field the unique tool of non-defective, acutely oncogenic retroviruses. This resulted in important insights into retroviral genetics. It provided the first biochemical evidence that a viral oncogene is not required for viral replication. It yielded a series of conditional and non-conditional mutants that illuminated viral gene functions and served as building blocks to construct a genetic map of a simple retrovirus. The non-defective avian sarcoma viruses led to the discovery of transformation-defective derivatives, and an isogenic pair of non-defective and transformation-defective viruses yielded a src probe that revealed the presence of src sequences in the cellular genome.
Personal note: I first met Jan Svoboda in 1964 at the International Conference on Avian Tumor Viruses at Duke University in Durham, North Carolina. Another particularly memorable encounter was in 1981 on the occasion of a symposium held by the International Association for Comparative Research on Leukemia and Related Diseases in Los Angeles. Jan and his wife Karin visited my home in Pasadena, and we had a long discussion about the Prague spring and its armed suppression. After Prague was liberated from Soviet domination, I visited Jan several times. Our friendship was very cordial. Since I was born in Czechoslovakia and spent my childhood there, the historical rivalry between Czechs and Germans in Czechoslovakia could have divided us, but it never entered our relationship. We were aware of it, but for both of us, the common heritage was the precious bond that prevailed.
Funding
This research received no external funding.
Conflicts of Interest
The author declares no conflict of interest.
References
- Temin, H.M.; Rubin, H. Characteristics of an assay for Rous sarcoma virus and Rous sarcoma cells in tissue culture. Virology 1958, 6, 669–688. [Google Scholar] [CrossRef]
- Hartley, J.W.; Rowe, W.P. Production of altered cell foci in tissue culture by defective Moloney sarcoma virus particles. Proc. Natl. Acad. Sci. USA 1966, 55, 780–786. [Google Scholar] [CrossRef] [PubMed]
- Huebner, R.J.; Hartley, J.W.; Rowe, W.P.; Lane, W.T.; Capps, W.I. Rescue of the defective genome of Moloney sarcoma virus from a noninfectious hamster tumor and the production of pseudotype sarcoma viruses with various murine leukemia viruses. Proc. Natl. Acad. Sci. USA 1966, 56, 1164–1169. [Google Scholar] [CrossRef] [PubMed]
- Somers, K.D.; Kirsten, W.H. Focus formation by murine sarcoma virus: Enhancement by DEAE-dextran. Virology 1968, 36, 155–157. [Google Scholar] [CrossRef]
- Klement, V.; Rowe, W.P.; Hartley, J.W.; Pugh, W.E. Mixed culture cytopathogenicity: A new test for growth of murine leukemia viruses in tissue culture. Proc. Natl. Acad. Sci. USA 1969, 63, 753–758. [Google Scholar] [CrossRef] [PubMed]
- Klement, V.; Hartley, J.W.; Rowe, W.P.; Huebner, R.J. Recovery of a hamster-specific, focus-forming, and sarcomagenic virus from a “noninfectious” hamster tumor induced by the Kirsten Mouse sarcoma virus. J. Natl. Cancer Inst. 1969, 43, 925–934. [Google Scholar]
- Kirsten, W.H.; Schauf, V.; McCoy, J. Properties of a murine sarcoma virus. Bibl. Haematol. 1970, 36, 246–249. [Google Scholar]
- Rowe, W.P.; Pugh, W.E.; Hartley, J.W. Plaque assay techniques for murine leukemia viruses. Virology 1970, 42, 1136–1139. [Google Scholar] [CrossRef]
- Levy, J.A.; Hartley, J.W.; Rowe, W.P.; Huebner, R.J. Studies of FBJ osteosarcoma virus in tissue culture. I. Biologic characteristics of the “C”-type viruses. J. Natl. Cancer Inst. 1973, 51, 525–539. [Google Scholar]
- Sklar, M.D.; White, B.J.; Rowe, W.P. Initiation of oncogenic transformation of mouse lymphocytes in vitro by Abelson leukemia virus. Proc. Natl. Acad. Sci. USA 1974, 71, 4077–4081. [Google Scholar] [CrossRef]
- Levy, J.A.; Hartley, J.W.; Rowe, W.P.; Huebner, R.J. Studies of FBJ osteosarcoma virus in tissue culture. II. Autoinhibition of focus formation. J. Natl. Cancer Inst. 1975, 54, 615–619. [Google Scholar] [PubMed]
- Poiesz, B.J.; Ruscetti, F.W.; Reitz, M.S.; Kalyanaraman, V.S.; Gallo, R.C. Isolation of a new type C retrovirus (HTLV) in primary uncultured cells of a patient with Sezary T-cell leukaemia. Nature 1981, 294, 268–271. [Google Scholar] [CrossRef] [PubMed]
- Vogt, P.K. Retroviral oncogenes: A historical primer. Nat. Rev. Cancer 2012, 12, 639–648. [Google Scholar] [CrossRef] [PubMed]
- Bryan, W.R.; Moloney, J.B.; Calnan, D. Stable standard preparations of the Rous sarcoma virus preserved by freezing and storage at low temperatures. J. Natl. Cancer Inst. 1954, 15, 315–329. [Google Scholar] [PubMed]
- Bryan, W.R. Enhancement of virus yield of Rous sarcomas. Acta Unio Int. Contra Cancrum 1959, 15, 764–767. [Google Scholar] [PubMed]
- Rubin, H.; Temin, H.M. A radiological study of cell-virus interaction in the Rous sarcoma. Virology 1959, 7, 75–91. [Google Scholar] [CrossRef]
- Temin, H.M. Further evidence for a converted, non-virus-producing state of Rous sarcoma virus-infected cells. Virology 1963, 20, 235–245. [Google Scholar] [CrossRef]
- Hanafusa, H.; Hanafusa, T.; Rubin, H. The defectiveness of Rous sarcoma virus. Proc. Natl. Acad. Sci. USA 1963, 49, 572–580. [Google Scholar] [CrossRef]
- Dougherty, R.M.; Di Stefano, H.S. Virus particles associated with “nonproducer” Rous sarcoma cells. Virology 1965, 27, 351–359. [Google Scholar] [CrossRef]
- Weiss, R. Spontaneous virus production from “non-virus producing” Rous sarcoma cells. Virology 1967, 32, 719–723. [Google Scholar] [CrossRef]
- Klucking, S.; Adkins, H.B.; Young, J.A. Resistance to infection by subgroups B, D, and E avian sarcoma and leukosis viruses is explained by a premature stop codon within a resistance allele of the tvb receptor gene. J. Virol. 2002, 76, 7918–7921. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.M.; Bacon, L.D.; Cheng, H.H.; Hunt, H.D. Development and validation of a PCR-RFLP assay to evaluate TVB haplotypes coding receptors for subgroup B and subgroup E avian leukosis viruses in White Leghorns. Avian Pathol. 2005, 34, 324–331. [Google Scholar] [CrossRef] [Green Version]
- Zhang, H.M.; Bacon, L.D.; Heidari, M.; Muir, W.M.; Groenen, M.A.; Zhang, Y.; Wong, G.K.; Fulton, J.E.; O’Sullivan, N.P.; Albers, G.A.; et al. Genetic variation at the tumour virus B locus in commercial and laboratory chicken populations assessed by a medium-throughput or a high-throughput assay. Avian Pathol. 2007, 36, 283–291. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vogt, P.K. A virus released by “nonproducing” Rous sarcoma cells. Proc. Natl. Acad. Sci. USA 1967, 58, 801–808. [Google Scholar] [CrossRef] [PubMed]
- Scheele, C.M.; Hanafusa, H. Proteins of helper-dependent RSV. Virology 1971, 45, 401–410. [Google Scholar] [CrossRef]
- Rychlikova, M.; Svoboda, J. Effects of anti-Rous sarcoma virus sera on a hemorrhagic disease of rats. Virology 1960, 10, 545–547. [Google Scholar] [CrossRef]
- Schmidt-Ruppin, K.H. Heterotransplantation of Rous Sarcoma and Rous Sarcoma Virus to Mammals. Oncologia 1964, 17, 247–272. [Google Scholar] [CrossRef] [PubMed]
- Zilber, L.A. Some data on the interaction of Rous sarcoma virus with mammalian cells. In Proceedings of the International Conference on Avian Tumor Viruses, Duke University, Durham, NC, USA, 31 March–3 April 1964; pp. 261–275. [Google Scholar]
- Thurzo, V.; Slabeyciusova, M.; Klimek, M.; Kovarova, V. New filtrable fowl tumor. Cesk. Onkol. 1954, 1, 230–234. [Google Scholar]
- Svoboda, J. Presence of chicken tumour virus in the sarcoma of the adult rat inoculated after birth with Rous sarcoma tissue. Nature 1960, 186, 980–981. [Google Scholar] [CrossRef]
- Simkovic, D.; Smida, J.; Thurzo, V. Release of tumour virus by cells of the B77 fowl virus tumour in vitro. Neoplasma 1961, 8, 495–500. [Google Scholar]
- Thurzo, V.; Smida, J.; Smidova-Kovarova, V.; Simkovic, D. Some properties of the fowl virus tumour B77. Acta Unio Int. Contra Cancrum 1963, 19, 304–305. [Google Scholar] [PubMed]
- Thurzo, V.; Simkovicova, M.; Lizonova, A. Group-Specific Antigens of Ascites Rat Tumors Induced by B77 Virus. Neoplasma 1971, 18, 551–554. [Google Scholar] [PubMed]
- Svec, J.; Hilubinova, K.; Lizorova, V.; Thurzo, V. Mammalian tropism of B77(RBI) virus. Expression of virus genome in the hamster sarcoma cell clones. Neoplasma 1975, 22, 133–145. [Google Scholar] [PubMed]
- Svoboda, J.; Chyle, P.; Simkovic, D.; Hilgert, I. Demonstration of the absence of infectious Rous virus in rat tumour XC, whose structurally intact cells produce Rous sarcoma when transferred to chicks. Folia Biol. 1963, 9, 77–81. [Google Scholar]
- Svoboda, J.; Hejnar, J.; Geryk, J.; Elleder, D.; Vernerova, Z. Retroviruses in foreign species and the problem of provirus silencing. Gene 2000, 261, 181–188. [Google Scholar] [CrossRef]
- Lounkova, A.; Draberova, E.; Senigl, F.; Trejbalova, K.; Geryk, J.; Hejnar, J.; Svoboda, J. Molecular events accompanying rous sarcoma virus rescue from rodent cells and the role of viral gene complementation. J. Virol. 2014, 88, 3505–3515. [Google Scholar] [CrossRef] [PubMed]
- Golde, A.; Vigier, P. Non-defectivity of the Rous virus of the Prague strain. C. R. Acad. Sci. Hebd. Seances Acad. Sci. D 1966, 262, 2793–2796. [Google Scholar] [PubMed]
- Lai, M.M.; Duesberg, P.H.; Horst, J.; Vogt, P.K. Avian tumor virus RNA: A comparison of three sarcoma viruses and their transformation-defective derivatives by oligonucleotide fingerprinting and DNA-RNA hybridization. Proc. Natl. Acad. Sci. USA 1973, 70, 2266–2270. [Google Scholar] [CrossRef] [PubMed]
- Lai, M.M.; Hu, S.S.; Vogt, P.K. Occurrence of partial deletion and substitution of the src gene in the RNA genome of avian sarcoma virus. Proc. Natl. Acad. Sci. USA 1977, 74, 4781–4785. [Google Scholar] [CrossRef] [Green Version]
- Toyoshima, K.; Vogt, P.K. Temperature sensitive mutants of an avian sarcoma virus. Virology 1969, 39, 930–931. [Google Scholar] [CrossRef]
- Friis, R.R.; Toyoshima, K.; Vogt, P.K. Conditional lethal mutants of avian sarcoma viruses. I. Physiology of ts 75 and ts 149. Virology 1971, 43, 375–389. [Google Scholar] [CrossRef]
- Martin, G.S. Rous sarcoma virus: A function required for the maintenance of the transformed state. Nature 1970, 227, 1021–1023. [Google Scholar] [CrossRef] [PubMed]
- Vogt, P.K. Spontaneous segregation of nontransforming viruses from cloned sarcoma viruses. Virology 1971, 46, 939–946. [Google Scholar] [CrossRef]
- Duesberg, P.H.; Vogt, P.K. Differences between the ribonucleic acids of transforming and nontransforming avian tumor viruses. Proc. Natl. Acad. Sci. USA 1970, 67, 1673–1680. [Google Scholar] [CrossRef] [PubMed]
- Bernstein, A.; MacCormick, R.; Martin, G.S. Transformation-defective mutants of avian sarcoma viruses: The genetic relationship between conditional and nonconditional mutants. Virology 1976, 70, 206–209. [Google Scholar] [CrossRef]
- Stehelin, D.; Varmus, H.E.; Bishop, J.M.; Vogt, P.K. DNA related to the transforming gene(s) of avian sarcoma viruses is present in normal avian DNA. Nature 1976, 260, 170–173. [Google Scholar] [CrossRef] [PubMed]
- Vennstrom, B.; Sheiness, D.; Zabielski, J.; Bishop, J.M. Isolation and characterization of c-myc, a cellular homolog of the oncogene (v-myc) of avian myelocytomatosis virus strain 29. J. Virol. 1982, 42, 773–779. [Google Scholar]
- Coffin, J.M.; Hughes, S.H.; Varmus, H.E. (Eds.) Retroviruses; Cold Spring Harbor Laboratory Press: Cold Spring Harbor, NY, USA, 1997. [Google Scholar]
- Robert C. Bast, C.M.C.; Hait, W.N.; Hong, W.K.; Kufe, D.W.; Piccart-Gebhart, M.; Pollock, R.E.; Weichselbaum, R.R.; Wang, H.; Holland, J.F. (Eds.) Holland-Frei Cancer Medicine, 9th ed.; Wiley Blackwell: Hoboken, NJ, USA, 2017. [Google Scholar]
- John Mendelsohn, J.W.G.; Howley, P.M.; Israel, M.A.; Thompson, C.B. (Eds.) The Molecular Basis of Cancer, 4th ed.; Elsevier Saunders: Philadelphia, PA, USA, 2014. [Google Scholar]
- Hanafusa, H.; Baltimore, D.; Smoler, D.; Watson, K.F.; Yaniv, A.; Spiegelman, S. Absence of polymerase protein in virions of alpha-type rous sarcoma virus. Science 1972, 177, 1188–1191. [Google Scholar] [CrossRef]
- Baltimore, D.; Verma, I.M.; Drost, S.; Mason, W.S. Temperature-sensitive DNA polymerase from Rous sarcoma virus mutants. Cancer 1974, 34, 1395–1397. [Google Scholar] [CrossRef]
- Friis, R.R.; Mason, W.S.; Chen, Y.C.; Halpern, M.S. A replication defective mutant of Rous sarcoma virus which fails to make a functional reverse transcriptase. Virology 1975, 64, 49–62. [Google Scholar] [CrossRef]
- Verma, I.M. Studies on reverse transcriptase of RNA tumor viruses. I. Localization of thermolabile DNA polymerase and RNase H activities on one polypeptide. J. Virol. 1975, 15, 121–126. [Google Scholar] [PubMed]
- Mason, W.S.; Yeater, C.; Bosch, J.V.; Wyke, J.A.; Friis, R.R. Fourteen temperature-sensitive replication mutants of Rous sarcoma virus. Virology 1979, 99, 226–240. [Google Scholar] [CrossRef]
- Duesberg, P.H.; Wang, L.-H.; Mellon, P.; Mason, W.S.; Vogt, P.K. Towards a complete genetic map of Rous sarcoma virus. In Animal Virology—Proceedings of the Fifth ICN-UCLA Symposium on Molecular Biology, Squaw Valley, 1976; Baltimore, D., Huang, A.S., Fox, C.F., Eds.; Academic Press: New York, NY, USA, 1976; pp. 107–125. [Google Scholar]
- Duesberg, P.H.; Wang, L.H.; Beemon, K.; Kawai, S.; Hanafusa, H. Sequences and functions of Rous sarcoma virus RNA. Hamatol. Bluttransfus 1976, 19, 327–340. [Google Scholar] [PubMed]
- Hayward, W.S.; Neel, B.G.; Astrin, S.M. Activation of a cellular onc gene by promoter insertion in ALV-induced lymphoid leukosis. Nature 1981, 290, 475–480. [Google Scholar] [CrossRef] [PubMed]
- Roelink, H.; Wagenaar, E.; Lopes da Silva, S.; Nusse, R. Wnt-3, a gene activated by proviral insertion in mouse mammary tumors, is homologous to int-1/Wnt-1 and is normally expressed in mouse embryos and adult brain. Proc. Natl. Acad. Sci. USA 1990, 87, 4519–4523. [Google Scholar] [CrossRef] [PubMed]
- Mladenov, Z.; Heine, U.; Beard, D.; Beard, J.W. Strain MC29 avian leukosis virus. Myelocytoma, endothelioma, and renal growths: Pathomorphological and ultrastructural aspects. J. Natl. Cancer Inst. 1967, 38, 251–285. [Google Scholar] [PubMed]
- Duesberg, P.H.; Bister, K.; Vogt, P.K. The RNA of avian acute leukemia virus MC29. Proc. Natl. Acad. Sci. USA 1977, 74, 4320–4324. [Google Scholar] [CrossRef] [PubMed]
- Maki, Y.; Bos, T.J.; Davis, C.; Starbuck, M.; Vogt, P.K. Avian sarcoma virus 17 carries the jun oncogene. Proc. Natl. Acad. Sci. USA 1987, 84, 2848–2852. [Google Scholar] [CrossRef]
- Vogt, P.K. Fortuitous convergences: The beginnings of JUN. Nat. Rev. Cancer 2002, 2, 465–469. [Google Scholar] [CrossRef] [PubMed]
- Chang, H.W.; Aoki, M.; Fruman, D.; Auger, K.R.; Bellacosa, A.; Tsichlis, P.N.; Cantley, L.C.; Roberts, T.M.; Vogt, P.K. Transformation of chicken cells by the gene encoding the catalytic subunit of PI 3-kinase. Science 1997, 276, 1848–1850. [Google Scholar] [CrossRef] [PubMed]
- Samuels, Y.; Wang, Z.; Bardelli, A.; Silliman, N.; Ptak, J.; Szabo, S.; Yan, H.; Gazdar, A.; Powell, S.M.; Riggins, G.J.; et al. High frequency of mutations of the PIK3CA gene in human cancers. Science 2004, 304, 554. [Google Scholar] [CrossRef] [PubMed]
- Payne, L.N.; Brown, S.R.; Bumstead, N.; Howes, K.; Frazier, J.A.; Thouless, M.E. A novel subgroup of exogenous avian leukosis virus in chickens. J. Gen. Virol. 1991, 72 Pt 4, 801–807. [Google Scholar] [CrossRef] [Green Version]
- Payne, L.N.; Howes, K.; Gillespie, A.M.; Smith, L.M. Host range of Rous sarcoma virus pseudotype RSV(HPRS-103) in 12 avian species: Support for a new avian retrovirus envelope subgroup, designated J. J. Gen. Virol. 1992, 73 Pt 11, 2995–2997. [Google Scholar] [CrossRef] [PubMed]
- Venugopal, K. Avian leukosis virus subgroup J: A rapidly evolving group of oncogenic retroviruses. Res. Vet. Sci. 1999, 67, 113–119. [Google Scholar] [CrossRef] [PubMed]
- Xu, B.; Dong, W.; Yu, C.; He, Z.; Lv, Y.; Sun, Y.; Feng, X.; Li, N.; Lee, L.F.; Li, M. Occurrence of avian leukosis virus subgroup J in commercial layer flocks in China. Avian Pathol. 2004, 33, 13–17. [Google Scholar] [CrossRef] [PubMed]
- Meng, F.; Li, Q.; Zhang, Y.; Cui, Z.; Chang, S.; Zhao, P. Isolation and characterization of subgroup J Avian Leukosis virus associated with hemangioma in commercial Hy-Line chickens. Poult. Sci. 2018, 97, 2667–2674. [Google Scholar] [CrossRef] [PubMed]
- Bagust, T.J.; Fenton, S.P.; Reddy, M.R. Detection of subgroup J avian leukosis virus infection in Australian meat-type chickens. Aust. Vet. J. 2004, 82, 701–706. [Google Scholar] [CrossRef]
- Malkinson, M.; Banet-Noach, C.; Davidson, I.; Fadly, A.M.; Witter, R.L. Comparison of serological and virological findings from subgroup J avian leukosis virus-infected neoplastic and non-neoplastic flocks in Israel. Avian Pathol. 2004, 33, 281–287. [Google Scholar] [CrossRef] [Green Version]
- Furukawa, S.; Tsukamoto, K.; Maeda, M. Multicentric histiocytosis related to avian leukosis virus subgroup J (ALV-J)-infection in meat-type local chickens. J. Vet. Med. Sci. 2014, 76, 89–92. [Google Scholar] [CrossRef]
- Zhao, Z.; Rao, M.; Liao, M.; Cao, W. Phylogenetic Analysis and Pathogenicity Assessment of the Emerging Recombinant Subgroup K of Avian Leukosis Virus in South China. Viruses 2018, 10, 194. [Google Scholar] [CrossRef]
- Sorge, J.; Hughes, S.H. Splicing of intervening sequences introduced into an infectious retroviral vector. J. Mol. Appl. Genet. 1982, 1, 547–559. [Google Scholar] [PubMed]
- Sorge, J.; Ricci, W.; Hughes, S.H. cis-Acting RNA packaging locus in the 115-nucleotide direct repeat of Rous sarcoma virus. J. Virol. 1983, 48, 667–675. [Google Scholar] [PubMed]
- Hughes, S.H.; Greenhouse, J.J.; Petropoulos, C.J.; Sutrave, P. Adaptor plasmids simplify the insertion of foreign DNA into helper-independent retroviral vectors. J. Virol. 1987, 61, 3004–3012. [Google Scholar] [PubMed]
- Hughes, S.H. The RCAS vector system. Folia Biol. 2004, 50, 107–119. [Google Scholar]
- Hughes, S. The RCAS System. Available online: Home.ncifcrf.gov/hivdrp/rcas/index.html (accessed on 12 December 2018).
- Rubin, H. The early history of tumor virology: Rous, RIF, and RAV. Proc. Natl. Acad. Sci. USA 2011, 108, 14389–14396. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kawai, S.; Hanafusa, H. Isolation of defective mutant of avian sarcoma virus. Proc. Natl. Acad. Sci. USA 1973, 70, 3493–3497. [Google Scholar] [CrossRef] [PubMed]
- Dutta, A.; Wang, L.H.; Hanafusa, T.; Hanafusa, H. Partial nucleotide sequence of Rous sarcoma virus-29 provides evidence that the original Rous sarcoma virus was replication defective. J. Virol. 1985, 55, 728–735. [Google Scholar]
- Wang, L.H.; Beckson, M.; Anderson, S.M.; Hanafusa, H. Identification of the viral sequence required for the generation of recovered avian sarcoma viruses and characterization of a series of replication-defective recovered avian sarcoma viruses. J. Virol. 1984, 49, 881–891. [Google Scholar]
- Svoboda, J.; Geryk, J.; Karakoz, I.; Rejthar, A. Isolation of two transforming viruses from sarcomas obtained in chickens inoculated intraembryonally with a transformation defective mutant of Prague strain Rous sarcoma virus. Folia Biol. 1985, 31, 135–151. [Google Scholar]
- Svoboda, J.; Kandala, J.C.; Geryk, J.; Pichrtova, J.; Guntaka, R.V. A transformation-competent recombinant between v-src and Rous-associated virus RAV-1. J. Virol. 1990, 64, 1873–1877. [Google Scholar]
- Svoboda, J. Conditions for tumorigenic action of Rous virus in heterologous relationships. Acta Unio Int. Contra Cancrum 1962, 18, 154–158. [Google Scholar] [PubMed]
- Svoboda, J.; Simkovic, D. The relationships between virus and cell in rat sarcomas induced by Rous virus. Acta Unio Int. Contra Cancrum 1963, 19, 302–303. [Google Scholar] [PubMed]
- Svoboda, J.; Vesely, P.; Vrba, M.; Jiranek, L. Biological properties of rat cells transformed in vitro by Rous sarcoma virus. Folia Biol. 1965, 11, 251–257. [Google Scholar]
- Svoboda, J.; Hlozanek, I.; Mach, O. Detection of chicken sarcoma virus after transfection of chicken fibroblasts with DNA isolated from mammalian cells transformed with Rous Virus. Folia Biol. 1972, 18, 149–153. [Google Scholar]
- Svoboda, J. Rous sarcoma virus. Intervirology 1986, 26, 1–60. [Google Scholar] [CrossRef] [PubMed]
- Svoboda, J. On board a raft or boat in the retrovirus sea. Proc. Natl. Acad. Sci. USA 2016, 113, 3927–3931. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
The figure summarizes the two possible genome versions of the original viral isolate of Peyton Rous and their subsequent evolution.
Figure 1.
The figure summarizes the two possible genome versions of the original viral isolate of Peyton Rous and their subsequent evolution.

© 2019 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Vogt, P.K. The Importance of Being Non-Defective: A Mini Review Dedicated to the Memory of Jan Svoboda. Viruses 2019, 11, 80. https://doi.org/10.3390/v11010080
AMA Style
Vogt PK. The Importance of Being Non-Defective: A Mini Review Dedicated to the Memory of Jan Svoboda. Viruses. 2019; 11(1):80. https://doi.org/10.3390/v11010080
Chicago/Turabian StyleVogt, Peter K. 2019. "The Importance of Being Non-Defective: A Mini Review Dedicated to the Memory of Jan Svoboda" Viruses 11, no. 1: 80. https://doi.org/10.3390/v11010080
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.