Variation in the Climate Sensitivity Dependent on Neighbourhood Composition in a Secondary Mixed Forest


 and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Site Description
2.2. Study Species
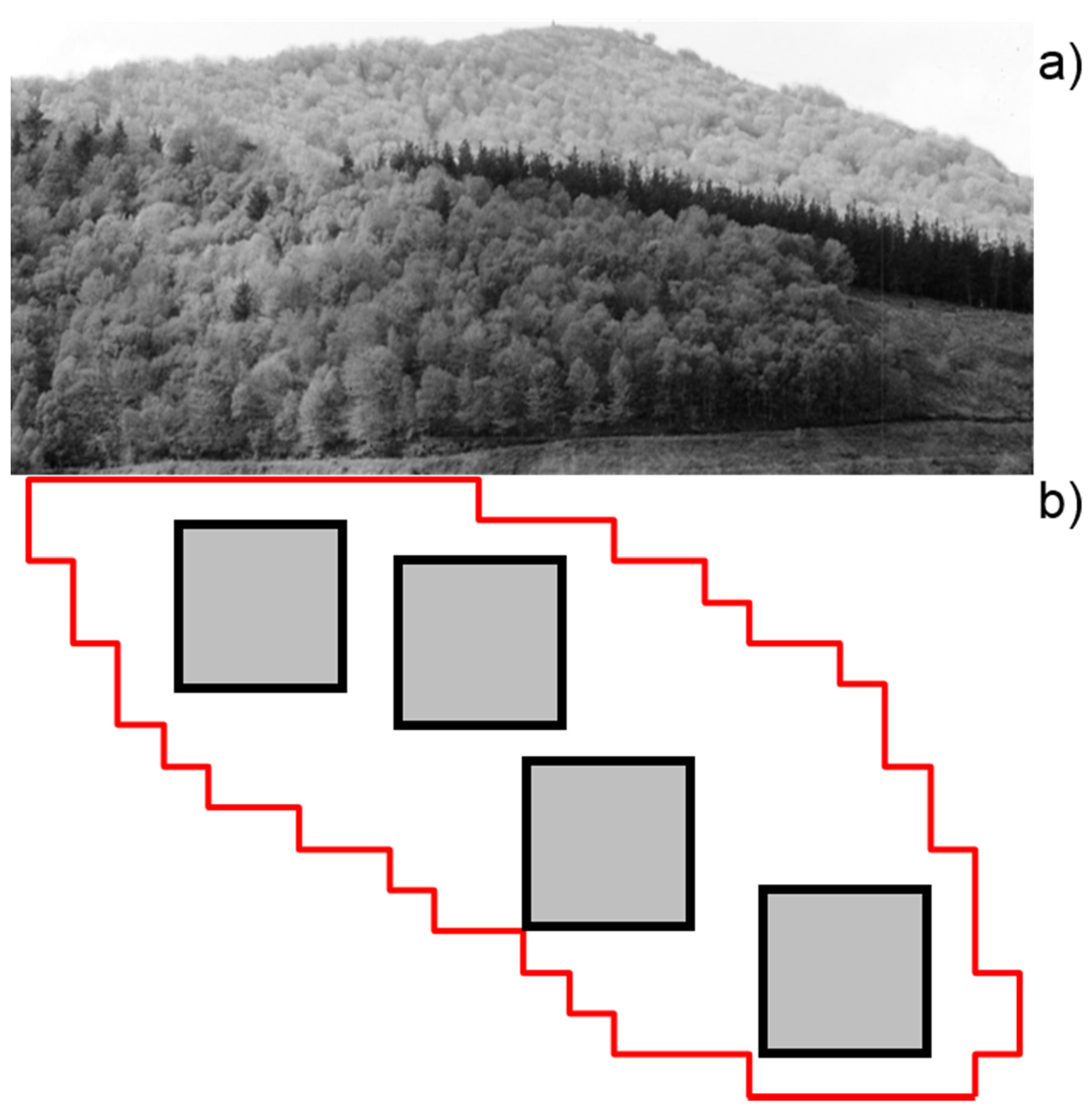
2.3. Experimental Design and Field Sampling
2.4. Core Preparation and Chronology Building
2.5. Data Analyses
2.5.1. Assessing the Growth Response to Climate at the Species Level
2.5.2. Assessing Individual Variation of Tree-Ring Growth Response to Climate
2.5.3. Calculating Individual and Neighbourhood Variables
2.5.4. Tree and Neighbourhood Effects on Growth
2.5.5. Assessing the Influence of Tree Level Factors on Climate–Growth Response
3. Results
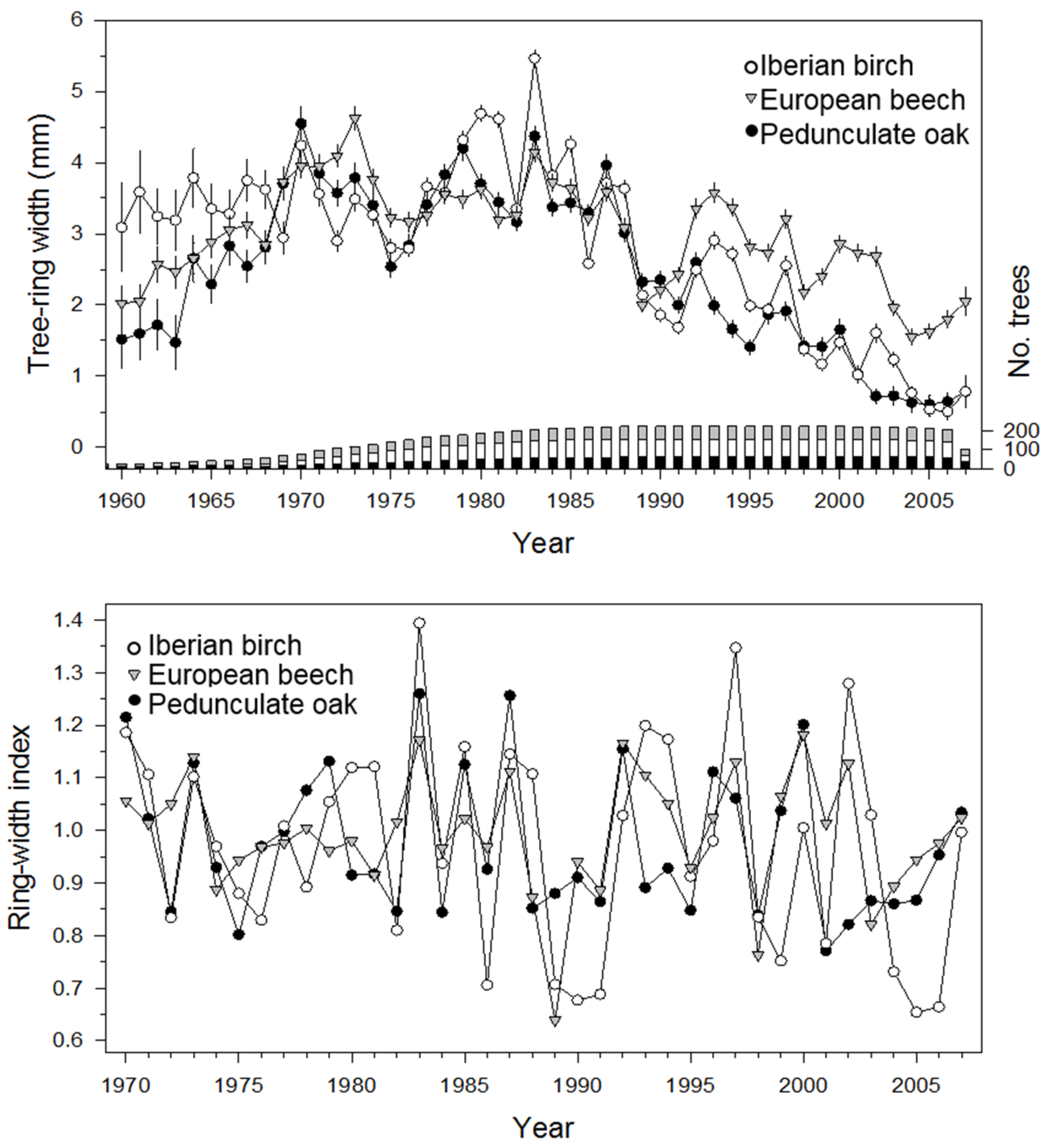
3.1. Sampled Tree Characteristics and Tree-Ring Chronologies
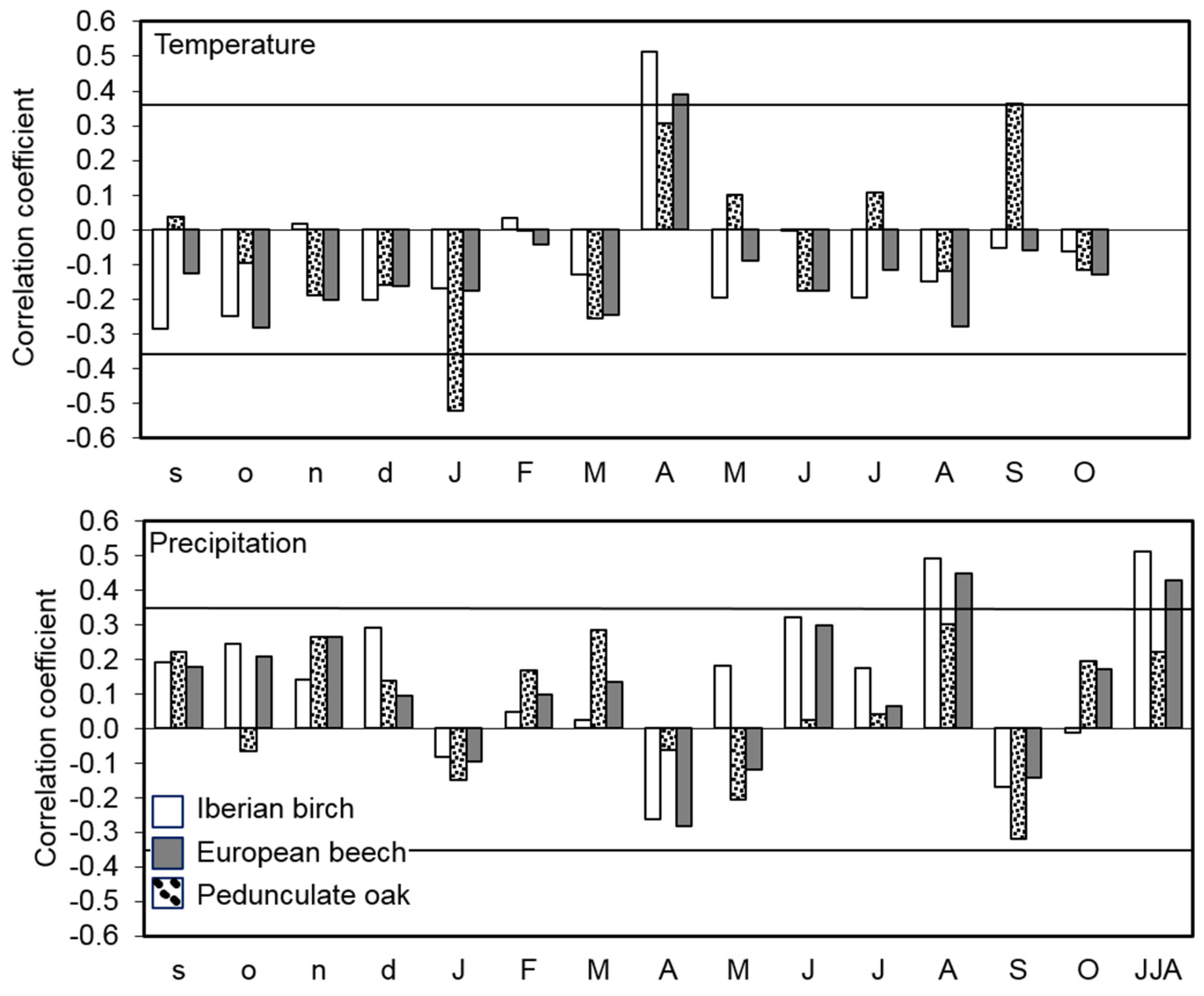
3.2. Tree-Ring Growth Response to Climate
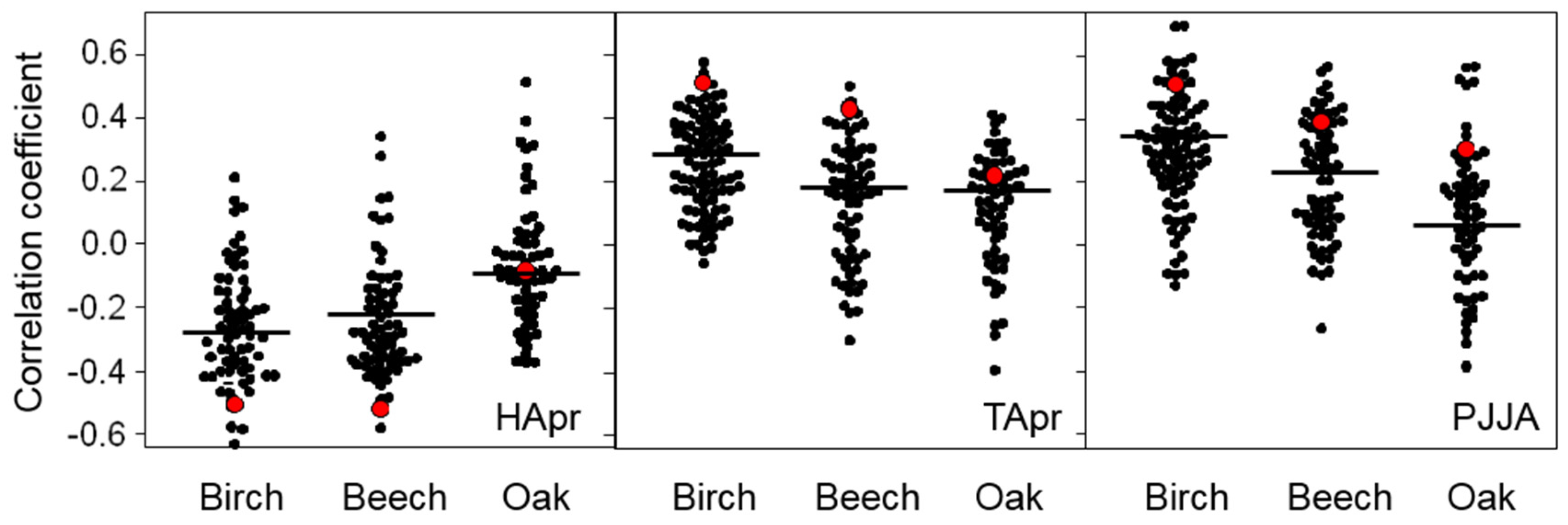
3.3. Individual Growth Response
3.4. Effect of Environmental Factors on the Climate–Growth Response
4. Discussion
4.1. Species-Dependent Growth Response to Climate
4.2. Individual Tree Responses to Climate
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Cook, E.R. The decomposition of tree-ring series for environmental studies. Tree-Ring Bull. 1987, 47, 37–59. [Google Scholar]
- Swetnam, T.W.; Allen, C.D.; Betancourt, J.L. Applied historical ecology: Using the past to manage for the future. Ecol. Appl. 1999, 9, 1189–1206. [Google Scholar] [CrossRef]
- Yang, F.; Wang, N.; Shi, F.; Ljungqvist, F.C.; Wang, S.; Fan, Z.; Lu, J. Multi-proxy temperature reconstruction from the West Qinling Mountains, China, for the past 500 years. PLoS ONE. 2013, 8, e57638. [Google Scholar] [CrossRef] [PubMed]
- Batllori, E.; Gutiérrez, E. Regional tree line dynamics in response to global change in the Pyrenees. J. Ecol. 2008, 96, 1275–1288. [Google Scholar] [CrossRef]
- Olano, J.M.; Arzac, A.; García-Cervigón, A.I.; Arx, G.; Rozas, V. New star on the stage: Amount of ray parenchyma in tree rings shows a link to climate. New Phytol. 2013, 198, 486–495. [Google Scholar] [CrossRef] [PubMed]
- Pieper, H.; Heinrich, I.; Heußner, K.U.; Helle, G. The influence of volcanic eruptions on growth of central European lowland trees in NE-Germany during the last millennium. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2014, 411, 155–166. [Google Scholar] [CrossRef]
- Fritts, H.C. Tree Rings and Climate; Academic Press: London, UK, 1976. [Google Scholar]
- Carrer, M. Individualistic and time-varying tree-ring growth to climate sensitivity. PLoS ONE 2011, 6, e22813. [Google Scholar] [CrossRef] [PubMed]
- Rozas, V. Individual-based approach as a useful tool to disentangle the relative importance of tree age, size and inter-tree competition in dendroclimatic studies. iForest 2014, 8, 187–194. [Google Scholar] [CrossRef]
- Rozas, V.; Olano, J.M. Environmental heterogeneity and neighbourhood interference modulate the individual response of Juniperus thurifera tree-ring growth to climate. Dendrochronologia 2013, 31, 105–113. [Google Scholar] [CrossRef]
- Jump, A.S.; Hunt, J.M.; Martínez-Izquierdo, J.A.; Peñuelas, J. Natural selection and climate change: Temperature-linked spatial and temporal trends in gene frequency in Fagus sylvatica. Mol. Ecol. 2006, 15, 3469–3480. [Google Scholar] [CrossRef] [PubMed]
- Carrer, M.; Urbinati, C. Age-dependent tree-ring growth responses to climate in Larix decidua and Pinus cembra. Ecology 2004, 85, 730–740. [Google Scholar] [CrossRef]
- Mérian, P.; Lebourgeois, F. Size-mediated climate–growth relationships in temperate forests: A multi-species analysis. For. Ecol. Manag. 2011, 261, 1382–1391. [Google Scholar] [CrossRef]
- Rozas, V.; DeSoto, L.; Olano, J.M. Sex specific, age dependent sensitivity of tree-ring growth to climate in the dioecious tree Juniperus thurifera. New Phytol. 2009, 182, 687–697. [Google Scholar] [CrossRef] [PubMed]
- Galván, J.D.; Camarero, J.J.; Gutiérrez, E. Seeing the trees for the forest: Drivers of individual growth responses to climate in Pinus uncinata mountain forests. J. Ecol. 2014, 102, 1244–1257. [Google Scholar] [CrossRef]
- Martin-Benito, D.; Kint, V.; Del Rio, M.; Muys, B.; Cañellas, I. Growth responses of West-Mediterranean Pinus nigra to climate change are modulated by competition and productivity: Past trends and future perspectives. For. Ecol. Manag. 2011, 262, 1030–1040. [Google Scholar] [CrossRef]
- Magruder, M.; Chhin, S.; Monks, A.; O’Brien, J. Effects of initial stand density and climate on red pine productivity within Huron National Forest, Michigan, USA. Forests 2012, 3, 1086–1103. [Google Scholar] [CrossRef]
- Castagneri, D.; Motta, R. A research gap in the interactive effects of climate and competition on trees growth. In Tree Growth: Influences, Layers and Types; Karam, W.P., Ed.; Nova Science Publisher: New York, NY, USA, 2010; pp. 93–102. [Google Scholar]
- Sánchez-Gómez, D.; Zavala, M.A.; Van Schalkwijk, D.B.; Urbieta, I.R.; Valladares, F. Rank reversals in tree growth along tree size, competition and climatic gradients for four forest canopy dominant species in Central Spain. Ann. For. Sci. 2008, 65, 1. [Google Scholar] [CrossRef]
- Linares, J.C.; Camarero, J.J.; Carreira, J.A. Competition modulates the adaptation capacity of forests to climatic stress: Insights from recent growth decline and death in relict stands of the Mediterranean fir Abies pinsapo. J. Ecol. 2010, 98, 592–603. [Google Scholar] [CrossRef]
- Tenzin, J.; Tenzin, K.; Hasenauer, H. Individual tree basal area increment models for broadleaved forests in Bhutan. Forestry 2017, 90, 367–380. [Google Scholar] [CrossRef]
- Gómez-Aparicio, L.; García-Valdés, R.; Ruíz-Benito, P.; Zavala, M.A. Disentangling the relative importance of climate, size and competition on tree growth in Iberian forests: Implications for forest management under global change. Glob. Chang. Biol. 2011, 17, 2400–2414. [Google Scholar] [CrossRef] [Green Version]
- Sánchez-Salguero, R.; Linares, J.C.; Camarero, J.J.; Madrigal-González, J.; Hevia, A.; Sánchez-Miranda, Á.; Ballesteros-Cánovas, J.A.; Alfaro-Sánchez, R.; García-Cervigón, A.I.; Bigler, C.; et al. Disentangling the effects of competition and climate on individual tree growth: A retrospective and dynamic approach in Scots pine. For. Ecol. Manag. 2015, 358, 12–25. [Google Scholar] [CrossRef]
- Zhang, J.; Huang, S.; He, F. Half-century evidence from western Canada shows forest dynamics are primarily driven by competition followed by climate. Proc. Natl. Acad. Sci. USA 2015, 112, 4009–4014. [Google Scholar] [CrossRef] [PubMed]
- Zambrano, J.; Marchand, P.; Swenson, N.G. Local neighbourhood and regional climatic contexts interact to explain tree performance. Proc. R. Soc. B 2017, 284. [Google Scholar] [CrossRef] [PubMed]
- Lebourgeois, F.; Eberlé, P.; Mérian, P.; Seynave, I. Social status-mediated tree-ring responses to climate of Abies alba and Fagus sylvatica shift in importance with increasing stand basal area. For. Ecol. Manag. 2014, 328, 209–218. [Google Scholar] [CrossRef]
- Gleason, K.E.; Bradford, J.B.; Bottero, A.; D’Amato, A.W.; Fraver, S.; Palik, B.J.; Battaglia, M.A.; Iverson, L.; Kenefic, L.; Kern, C.C. Competition amplifies drought stress in forests across broad climatic and compositional gradients. Ecosphere 2017, 8, e01849. [Google Scholar] [CrossRef]
- González de Andrés, E.; Camarero, J.J.; Blanco, J.A.; Imbert, J.B.; Lo, Y.-H.; Sangüesa-Barreda, G.; Castillo, F.J. Tree-to-tree competition in mixed European beech–Scots pine forests has different impacts on growth and water-use efficiency depending on site conditions. J. Ecol. 2017, 106, 59–75. [Google Scholar] [CrossRef]
- Lebourgeois, F.; Gomez, N.; Pinto, P.; Mérian, P. Mixed stands reduce Abies alba tree-ring sensitivity to summer drought in the Vosges mountains, western Europe. For. Ecol. Manag. 2013, 303, 61–71. [Google Scholar] [CrossRef]
- Mölder, I.; Leuschner, C. European beech grows better and is less drought sensitive in mixed than in pure stands: Tree neighbourhood effects on radial increment. Trees 2014, 28, 777–792. [Google Scholar] [CrossRef]
- Valladares, F.; Niinemets, Ü. Shade tolerance, a key plant feature of complex nature and consequences. Annu. Rev. Ecol. Evol. Syst. 2008, 39, 237–257. [Google Scholar] [CrossRef]
- Bolte, A.; Eisenhauer, D.R.; Ehrhart, H.P.; Groß, J.; Hanewinkel, M.; Kölling, C.; Profft, I.; Rohde, M.; Röhe, P.; Amereller, K. Klimawandel und Forstwirtschaft—Übereinstimmungen und Unterschiede bei der Einschätzung der AnpassungsnotwendigkeitenundAnpassungsstrategien der Bundesländer. Landbauforsch. vTI Agric. For. Res. 2009, 59, 269–278. [Google Scholar]
- Vanoni, M.; Bugmann, H.; Nötzli, M.; Bigler, C. Quantifying the effects of drought on abrupt growth decreases of major tree species in Switzerland. Ecol. Evol. 2016, 6, 3555–3570. [Google Scholar] [CrossRef] [PubMed]
- Weemstra, M.; Eilmann, B.; Sass-Klaassen, U.G.; Sterck, F.J. Summer droughts limit tree growth across 10 temperate species on a productive forest site. For. Ecol. Manag. 2013, 306, 142–149. [Google Scholar] [CrossRef]
- Walentowski, H.; Falk, W.; Tobias, M.; Kunz, J.; Bräuning, A.; Meinardus, C.; Zang, C.; Sutcliffe, L.; Leuschner, C. Assessing future suitability of tree species under climate change by multiple methods: A case study in southern Germany. Ann. For. Sci. 2017, 60, 101. [Google Scholar] [CrossRef]
- Pretzsch, H.; Schutze, G. Effect of tree species mixing on the size structure, density, and yield of forest stands. Eur. J. For. Res. 2016, 135, 1–22. [Google Scholar] [CrossRef]
- Pretzsch, H.; Schütze, G.; Uhl, E. Resistance of European tree species to drought stress in mixed versus pure forests: Evidence of stress release by inter specific facilitation. Plant Biol. 2013, 15, 483–495. [Google Scholar] [CrossRef] [PubMed]
- Chhin, S.; Wang, G. Climatic Sensitivity of a Mixed Forest Association of White Spruce and Trembling Aspen at Their Southern Range Limit. Forests 2016, 7, 235. [Google Scholar] [CrossRef]
- Hartmann, H.; Messier, C. Interannual variation in competitive interactions from natural and anthropogenic disturbances in a temperate forest tree species: Implications for ecological interpretation. For. Ecol. Manag. 2011, 261, 1936–1944. [Google Scholar] [CrossRef] [Green Version]
- Olano, J.M.; Laskurain, N.A.; Escudero, A.; De La Cruz, M. Why and where do adult trees die in a young secondary temperate forest? The role of neighbourhood. Ann. For. Sci. 2009, 66, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Hohl, R.; Schweingruber, F.H.; Schiesser, H.H. Reconstruction of severe hailstorm occurrence with tree rings: A case study in central Switzerland. Tree-Ring Res. 2002, 58, 11–22. [Google Scholar]
- Herrera, J.; Laskurain, N.A.; Loidi, J.; Escudero, A.; Olano, J.M. Sucesión secundaria en un abedular-hayedo en el Parque Natural de Urquiola. Vizcaya. Lazaroa 2001, 22, 59–66. [Google Scholar]
- Grissino-Mayer, H.D. Evaluating crossdating accuracy: A manual and tutorial for the computer program COFECHA. Tree-Ring Res. 2001, 57, 205–221. [Google Scholar]
- Cook, E.R.; Holmes, R.L. Guide for computer program ARSTAN. In The International Tree-Ring Data Bank Program Library, Version 2.0.; University of Arizona: Tucson, AZ, USA, 1996; Volume 2, pp. 75–87. [Google Scholar]
- Helama, S.; Lindholm, M.; Timonen, M.; Eronen, M. Detection of climate signal in dendrochronological data analysis: A comparison of tree-ring standardization methods. Theor. Appl. Climatol. 2004, 79, 239–254. [Google Scholar] [CrossRef]
- De Castro, M.D.; Martín-Vide, J.; Alonso, S. El clima de España: Pasado, Presente y Escenarios de Clima Para el Siglo XXI; Impactos del Cambio Climatico en España; Ministerio Medio Ambiente: Madrid, Spain, 2005. [Google Scholar]
- Woods, K.D. Dynamics in late-successional hemlock-hardwood forests over three decades. Ecology 2000, 81, 110–126. [Google Scholar]
- Calcagno, V. Glmulti: Model Selection and Multimodel Inference Made Easy; R Package Version, 1.7; R Package Vignette: Madison, WI, USA, 2013. [Google Scholar]
- Bunn, A.G.; Goetz, S.J.; Fiske, G.J. Observed and predicted responses of plant growth to climate across Canada. Geophys. Res. Lett. 2005, 32, L16710. [Google Scholar] [CrossRef]
- Chmielewski, F.M.; Rötzer, T. Response of tree phenology to climate change across Europe. Agric. For. Meteorol. 2001, 108, 101–112. [Google Scholar] [CrossRef]
- Čufar, K.; Prislan, P.; de Luis, M.; Gričar, J. Tree-ring variation, wood formation and phenology of beech (Fagus sylvatica) from a representative site in Slovenia, SE Central Europe. Trees 2008, 22, 749–758. [Google Scholar] [CrossRef]
- Lupi, C.; Morin, H.; Deslauriers, A.; Rossi, S. Xylem phenology and wood production: Resolving the chicken or egg dilemma. Plant Cell Environ. 2010, 33, 1721–1730. [Google Scholar] [CrossRef] [PubMed]
- Piovesan, G.; Biondi, F.; Filippo, A.D.; Alessandrini, A.; Maugeri, M. Drought driven growth reduction in old beech (Fagus sylvatica L.) forests of the central Apennines, Italy. Glob. Chang. Biol. 2008, 14, 1265–1281. [Google Scholar] [CrossRef]
- Rozas, V.; Camarero, J.J.; Sangüesa-Barreda, G.; Souto, M.; García-González, I. Summer drought and ENSO-related cloudiness distinctly drive Fagus sylvatica growth near the species rear-edge in northern Spain. Agric. For. Meteorol. 2015, 201, 153–164. [Google Scholar] [CrossRef]
- Scharnweber, T.; Manthey, M.; Wilmking, M. Differential radial growth patterns between beech (Fagus sylvatica L.) and oak (Quercus robur L.) on periodically waterlogged soils. Tree Physiol. 2013, 33, 425–437. [Google Scholar] [CrossRef] [PubMed]
- Essiamah, S.; Eschrich, W. Water uptake in deciduous trees during winter and the role of conducting tissues in spring reactivation. IAWA J. 1986, 7, 31–38. [Google Scholar] [CrossRef]
- Hacke, U.; Sauter, J.J. Drought-induced xylem dysfunction in petioles, branches, and roots of Populus balsamifera L. and Alnus glutinosa. L. Gaertn. Plant Physiol. 1996, 111, 413–417. [Google Scholar] [CrossRef] [PubMed]
- Alla, A.Q.; Camarero, J.J. Contrasting responses of radial growth and wood anatomy to climate in a Mediterranean ring-porous oak: Implications for its future persistence or why the variance matters more than the mean. Eur. J. For. Res. 2012, 131, 1537–1550. [Google Scholar] [CrossRef]
- González, I.G.; Eckstein, D. Climatic signal of earlywood vessels of oak on a maritime site. Tree Physiol. 2003, 23, 497–504. [Google Scholar] [CrossRef] [PubMed]
- Barbaroux, C.; Bréda, N.; Dufrêne, E. Distribution of above-ground and below-ground carbohydrate reserves in adult trees of two contrasting broad-leaved species. Quercus petraea and Fagus sylvatica. New Phytol. 2003, 157, 605–615. [Google Scholar] [CrossRef]
- Rozas, V.; Lamas, S.; García-González, I. Differential tree-growth responses to local and large-scale climatic variation in two Pinus and two Quercus species in northwest Spain. Ecoscience 2009, 16, 299–310. [Google Scholar] [CrossRef]
- Saa Requejo, A.; García Moreno, R.; Diaz Alvarez, M.C.; Tarquis, A.M.; Burgaz, F. Analysis of hail damages and temperature series for peninsular Spain. Nat. Hazards Earth Syst. Sci. 2011, 11, 3415–3422. [Google Scholar] [CrossRef] [Green Version]
- Intergovernmental Panel on Climate Change (IPCC). 2013 Climate Change: The Physical Science Basis; Contribution of Working Group I to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change; Stocker, T.F., Qin, D., Plattner, G.K., Tignor, M., Allen, S.K., Boschung, J., Nauels, A., Xia, Y., Bex, V., Midgley, P.M., Eds.; IPCC: Cambridge, UK; New York, NY, USA, 2013. [Google Scholar]
- Augspurger, C.K. Spring 2007 warmth and frost: Phenology, damage and refoliation in a temperate deciduous forest. Funct. Ecol. 2009, 23, 1031–1039. [Google Scholar] [CrossRef]
- Fu, Y.H.; Campioli, M.; Deckmyn, G.; Janssens, I.A. Sensitivity of leaf unfolding to experimental warming in three temperate tree species. Agr. For. Meteorol. 2013, 181, 125–132. [Google Scholar] [CrossRef]
- Grossiord, C.; Gessler, A.; Granier, A.; Berger, S.; Bréchet, C.; Hentschel, R.; Hommel, R.; Scherer-Lorenzen, M.; Bonal, D. Impact of interspecific interactions on the soil water uptake depth in a young temperate mixed species plantation. J. Hydrol. 2014, 519, 3511–3519. [Google Scholar] [CrossRef]
- Leuschner, C.; Coners, H.; Icke, R. In situ measurement of water absorption by fine roots of three temperate trees: Species differences and differential activity of superficial and deep roots. Tree Physiol. 2004, 24, 1359–1367. [Google Scholar] [CrossRef] [PubMed]
- Mauer, O.; Palátová, E. The role of root system in silver birch (Betula pendula Roth) dieback in the air-polluted area of Krušné hory Mts. J. For. Sci. 2003, 49, 191–199. [Google Scholar]
- Piutti, E.; Cescatti, A. A quantitative analysis of the interactions between climatic response and intraspecific competition in European beech. Can. J. For. Res. 1997, 27, 277–284. [Google Scholar] [CrossRef]
- Zapater, M.; Hossann, C.; Bréda, N.; Bréchet, C.; Bonal, D.; Granier, A. Evidence of hydraulic lift in a young beech and oak mixed forest using 18O soil water labelling. Trees 2011, 25, 885–894. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | No. Trees | DBH (cm) | Height (m) | Age at 1.3 m (Years) |
|---|---|---|---|---|
| Iberian birch | 95 | 20.1 ± 5.8 (11.1–44.4) | 15.0 ± 4.7 (6.5–31.8) | 33 ± 6 |
| European beech | 70 | 23.0 ± 14.5 (6.0–59.7) | 14.7 ± 5.4 (5.3–38.2) | 38 ± 12 |
| Pedunculate oak | 63 | 20.0 ± 10.3 (6.6–47.5) | 13.5 ± 5.0 (5.9–32.3) | 35 ± 10 |
| Tree species | Raw Data | Residual Chronology | |||
|---|---|---|---|---|---|
| Tree-ring width, mean ± SD (mm) | AC1 | MSx | Rbar | EPS | |
| Iberian birch | 2.78 ± 1.75 | 0.62 | 0.21 | 0.29 | 0.97 |
| European beech | 3.03 ± 1.26 | 0.63 | 0.19 | 0.23 | 0.91 |
| Pedunculate oak | 2.46 ± 1.59 | 0.74 | 0.13 | 0.16 | 0.94 |
| Species | IfB | IfF | IfQ | Cp | IfT |
|---|---|---|---|---|---|
| Iberian birch | 38.2 ± 14.9 | 15.5 ± 12.5 a | 11.6 ± 8.8 | 8.2 ± 4.7 a | 68.2 ± 16.2 |
| European beech | 32.7 ± 16.0 | 20.8 ± 12.2 b | 13.5 ± 9.7 | 10.9 ± 9.0 b | 70.0 ± 20.6 |
| Pedunculate oak | 36.3 ± 15.0 | 17.9 ± 13.2 a,b | 12.4 ± 10.3 | 11.4 ± 7.3 b | 69.8 ± 20.2 |
| Species Selected | x | y | DBH | H | IfB | IfF | Cp | Adjusted r2 |
|---|---|---|---|---|---|---|---|---|
| Iberian birch | − | − | − | 0.107 | ||||
| European beech | + | + | + | − | − | 0.543 | ||
| Pedunculate oak | + | + | 0.395 |
| Climatic Variable | Species | x | y | DBH | H | IfB | IfF | IfQ | Adjusted r2 |
|---|---|---|---|---|---|---|---|---|---|
| April mean temperature | Iberian birch | + | − | 0.087 | |||||
| European beech | + | − | 0.036 | ||||||
| Pedunculate oak | + | + | + | + | + | 0.177 | |||
| April hail | Iberian birch | − | + | − | 0.163 | ||||
| European beech | + | − | 0.088 | ||||||
| Pedunculate oak | − | − | − | 0.156 | |||||
| June–August precipitation | Iberian birch | + | − | 0.126 | |||||
| European beech | + | + | 0.017 | ||||||
| Pedunculate oak | + | + | + | + | + | 0.198 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Laskurain, N.A.; Aldezabal, A.; Odriozola, I.; Camarero, J.J.; Olano, J.M. Variation in the Climate Sensitivity Dependent on Neighbourhood Composition in a Secondary Mixed Forest. Forests 2018, 9, 43. https://doi.org/10.3390/f9010043
Laskurain NA, Aldezabal A, Odriozola I, Camarero JJ, Olano JM. Variation in the Climate Sensitivity Dependent on Neighbourhood Composition in a Secondary Mixed Forest. Forests. 2018; 9(1):43. https://doi.org/10.3390/f9010043
Chicago/Turabian StyleLaskurain, Nere Amaia, Arantza Aldezabal, Iñaki Odriozola, Jesús Julio Camarero, and José Miguel Olano. 2018. "Variation in the Climate Sensitivity Dependent on Neighbourhood Composition in a Secondary Mixed Forest" Forests 9, no. 1: 43. https://doi.org/10.3390/f9010043