1. Introduction
Forests are increasingly recognized as a critical element of the global ecosystem, given their importance in providing multiple environmental services, such as carbon sequestration and storage, biodiversity conservation, climate change mitigation, poverty alleviation, and watershed protection [
1]. Conservation of species diversity in managed forests is an important objective in sustainable forest management [
2,
3]. Plant diversity in forests, a measure of community structural and functional complexity, maintains the operation of an ecosystem on a complex spatial–temporal scale [
4,
5]. High species diversity is the basis of ecosystem stability, and this factor likely facilitates the optimization of ecosystem functions [
6]. As a comprehensive feature of plant community structure and function, stability is not only closely related to community structure and function, but is also correlated with the nature and intensity of external interference [
7,
8,
9]. Most tropical forests, even those in protected areas, are influenced by human activity [
10]. To meet livelihoods, forest harvesting needs can impact forest regeneration, structure, and diversity [
11], but there is scope for considerable variation with location, human activities, and histories [
12]. Different types and intensities of local resource extraction can lead to varying outcomes even within one forest [
13]. So, the diversity in human activities and their impacts call for different interventions.
Southeast China is located in the subtropical zone. The mixed Chinese fir (
Cunninghamia lanceolata (Lamb.) Hook.), pine (
Pinus massoniana Lamb.), and hardwood forest in southeastern China is a major assemblage in natural secondary forests. After conifer species were selected for harvesting, the mixed Chinese fir, pine, and hardwood forest was changed into the mixed evergreen broadleaf forest. Evergreen broadleaf forest is the common zonal vegetation found in the northern Fujian Province of China and is one of the typical types of vegetation in subtropical areas. Evergreen broadleaf forest is characterized by a rich floristic composition and biodiversity, as well as significant ecological functions. However, many primary forests have been or are currently being converted into secondary forests because of historical reasons and human interference [
14]. Except for several natural conservation areas, only a few primitive, evergreen, broadleaf forests are left because of human interference, which is unsustainable at the landscape and timber stand level. Sustainable forest management is being advanced by the forestry sector with the expectation, amongst others, that it will reduce the environmental impacts caused by deforestation [
15]. Conservation and utilization are both integral parts of sustainable forest management. Forest selective harvesting and clear-cut harvesting are the management practices most frequently used for silvicultural rotation and timber harvesting worldwide [
16,
17]. In terms of the ecological perspective of forest harvesting, selective harvesting is common and most reasonable as applied to natural secondary forests based on forest characteristics and geographic locations [
18,
19]. Evergreen broadleaf forest is widely recognized as providing ecological, economic, and social benefits and supporting governmental efforts related to biodiversity conservation and forest protection [
20]. A major challenge of forest management is to maintain the biodiversity and integrity of the forests, while at the same time satisfying human needs through productive activities. While the selective extraction of natural resources has less severe consequences on biodiversity and ecosystem function than the complete removal of vegetation, such consequences need to be evaluated in detail [
21,
22,
23]. Therefore, investigating the long-term impact of harvesting intensity on community species diversity and stability is significant to forest management theory and practice.
Vast expanses of tropical forests worldwide are being impacted by selective harvesting [
24]. To select the appropriate level of harvesting intensity for a certain stand is a basic challenge in forest management planning. The decision depends on economic as well as biological factors [
25]. Forest management employing clear-cut harvesting and artificial reforestation has been the dominant silviculture regime in natural forests in China for 50–60 years. After 1998, the implementation of natural forest protection projects saw increased attention drawn to uneven-aged forests, selective harvesting, and natural reforestation. This increased interest is partly due to expected enhancements for landscape aesthetics and biodiversity, and to expected benefits for reforestation costs, timber quality, and profits. Selective harvesting has certain characteristics: (i) only a part of the standing volume is harvested; (ii) conditions for recruitment are favoured in order to maintain or develop an uneven-aged forest structure; (iii) the decision unit is the individual tree, or small groups of trees; and (iv) the choice of trees to be harvested is based on certain specified attributes (e.g., tree size, timber quality, volume increment, or financial maturity) [
25]. Long-term field experiments with selective harvesting in uneven-aged coniferous forests have shown that inappropriate forest structures and poor conditions for natural reforestation can cause low volume production, and hence, low profitability [
26]. Previous research on plant diversity in forests mainly focused on biodiversity features and differences among various undisturbed forest types and communities [
27]. In terms of human interference, the effect of selective harvesting intensity on species diversity and stability should be described in detail. For instance, the monitoring of changes in species diversity and stability during vegetation restoration after different harvesting intensities is one of the main research goals of sustainable forest management [
28,
29]. Many studies have referred to the impact of harvesting intensity on biodiversity [
30,
31,
32].
For the purpose of timber production, the traditional clear-cut harvesting was a productive forest operation. It only considered economic benefits and seriously damaged the ecological environment. A large number of studies have shown that selective harvesting is the most suitable management of natural secondary forests [
19,
25,
33]. But, for specific natural secondary forest, in order to protect species diversity and maintain community stability, the suitable selective harvesting intensity is not yet clear. To date, only a few studies have reported on the effect of selective harvesting intensity on plant species diversity and community stability of natural, secondary forests in subtropical areas. Most of the studies used space instead of time (temporary plots) to analyze the short-term effects of harvesting on species diversity, but few established long-term, fixed plots for the study [
19,
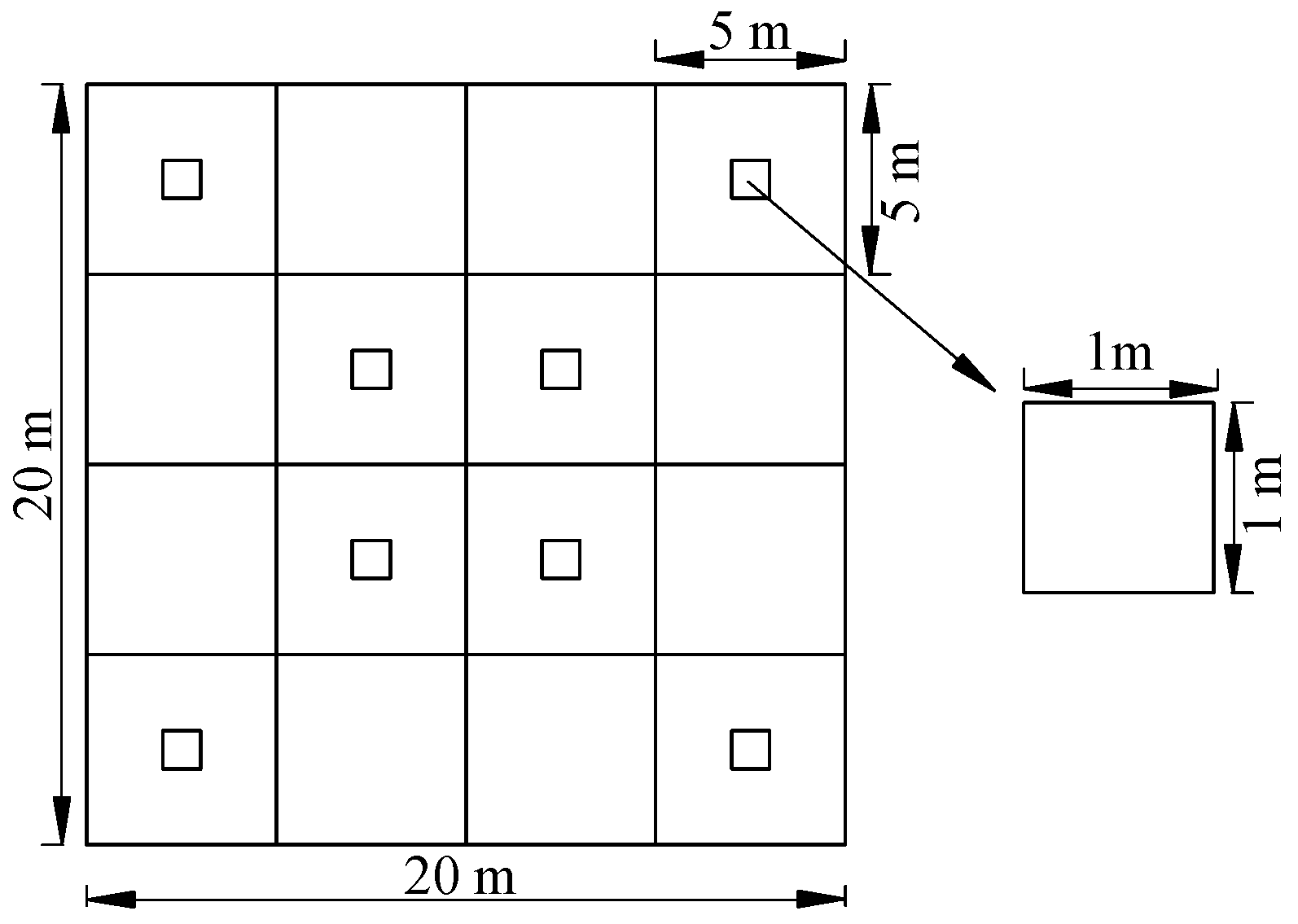
33]. In order to manage the forest scientifically, our study examined the long-term effects of harvesting intensity on the species diversity and community stability of natural secondary forests in subtropical areas 15 years after harvesting. We addressed the following two questions. Was the plant species diversity different in natural secondary forests with different harvesting intensities? Was plant community stability different in natural secondary forests with different harvesting intensities?
4. Discussion
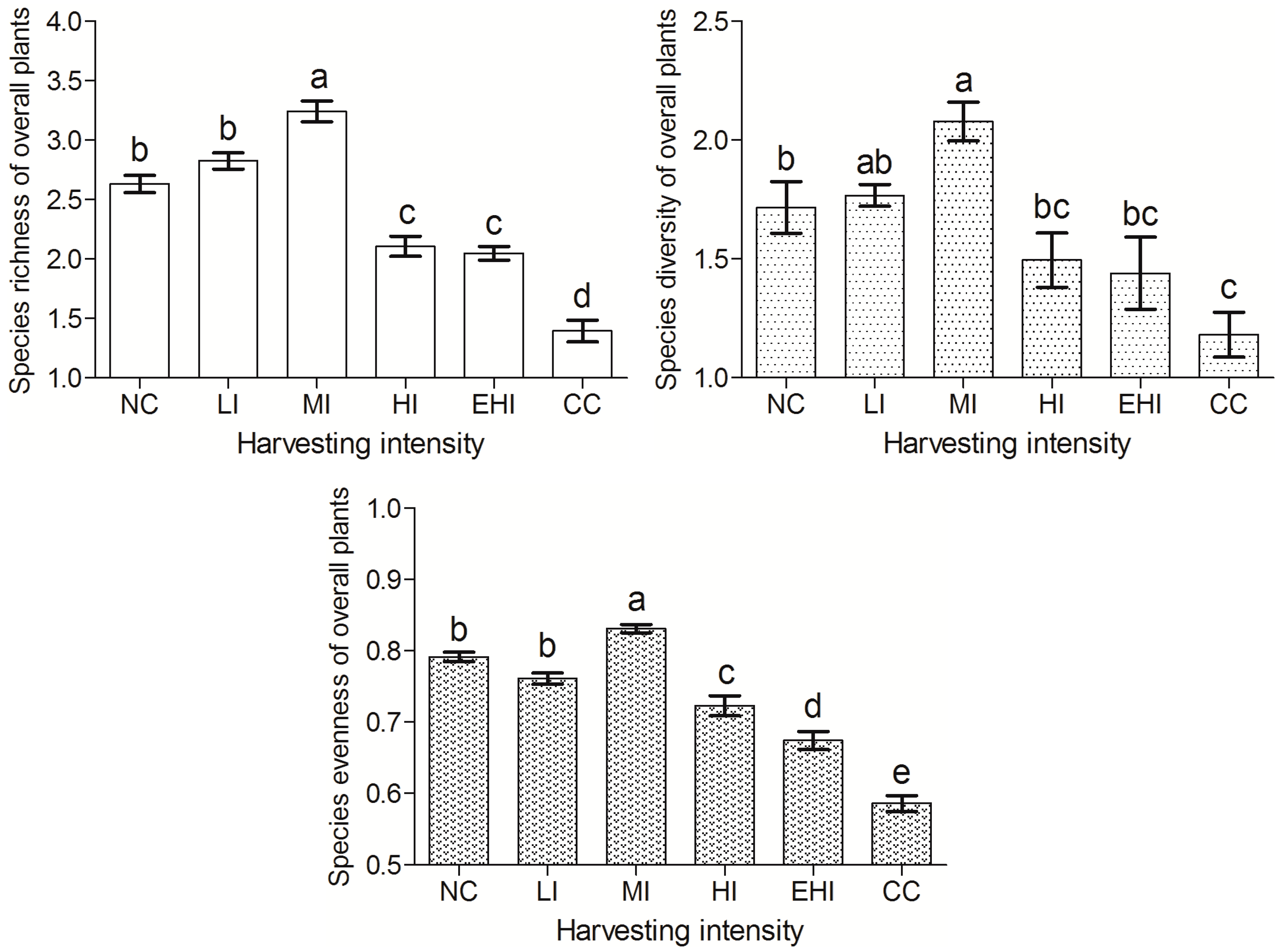
Numerous studies have shown that disturbances such as harvesting and other human activities in some cases cause an immediate decline in plant diversity followed by a recovery [
46,
47]. But, some studies have shown an increase in species richness after harvesting or other disturbance. Sassen (2013) et al. found that human impacts, as well as natural gradients, had major impacts on species richness patterns, and several areas in intermediate states of disturbance showed higher tree species richness than either old-growth forest or more severely degraded areas [
12]. The results of some perious studies were different because these influences had a close relationship with location, histories, harvesting intensity, and recovery time, and so on [
48,
49]. In our study, we found the plant species richness, diversity, and evenness in natural secondary forest increased after selective harvesting at low and medium intensities, and these parameters were the highest with selective harvesting at a medium intensity. This conclusion was similar to some related studies, and the course of succession in subtropical forests seems to support the intermediate disturbance hypothesis [
21,
50,
51]. After selective harvesting at low and medium intensities, a certain amount of forest space was released, sunlight in the forest was enhanced, and the soil environment was improved [
34]. Changes of these factors were conducive to creating good conditions for plant growth, promoting natural reforestation, and increasing species richness, diversity, and evenness [
52,
53].
Non-harvesting might require a long time to facilitate biological features of vegetation communities and restored species diversity [
29]. Some studies have shown that the percent of species lost was significantly higher in reference (non-harvesting) than harvested plots [
35]. In our study, species diversity indices of selective harvesting at a medium intensity were significantly greater than non-harvesting, but these parameters were not significantly different between non-harvesting and selective harvesting of low intensity. This was because the forest was inhibited by a high canopy density (>0.9) due to non-harvesting, which resulted in reduced sunlight exposure and thus, a limited growth of understory shrubs and herbs. Although a natural forests protection project was proposed by the Chinese government after 1998, the natural forests require scientific management.
Some studies assessing the effects of different types of disturbances, such as firewood harvesting [
46,
54], slash-and-burn [
55], or cattle grazing [
56,
57], have found that species diversity was reduced as the intensity and frequency of disturbance increased [
46,
58]. In our study, plant species richness, diversity, and evenness significantly decreased with selective harvesting at high and extra-high intensities, as well as with clear-cut harvesting. This might be due to overly large canopy gaps under over-harvesting [
33], resulting in major changes to the ecological environment, such as the stand structure, soil moisture, porosity, and nutrients [
19,
50,
51]. For instance, the worst-case scenario was from clear-cut harvesting, which resulted in the disappearance of the tree layer and a significant change in stand structure. Moreover, the
Dicranopteris pedata (Houtt.) Nakaike coverage rate was very high (>90%), so plant species of shrub and herb layers were few. The reduction in species diversity, changes in patterns of dominance, and the proliferation of species associated with disturbed sites suggested that current practices of selective harvesting at an exorbitant intensity require adjustments to make this forest management application more consistent with the local conservation of woody plant species diversity and community structure [
23].
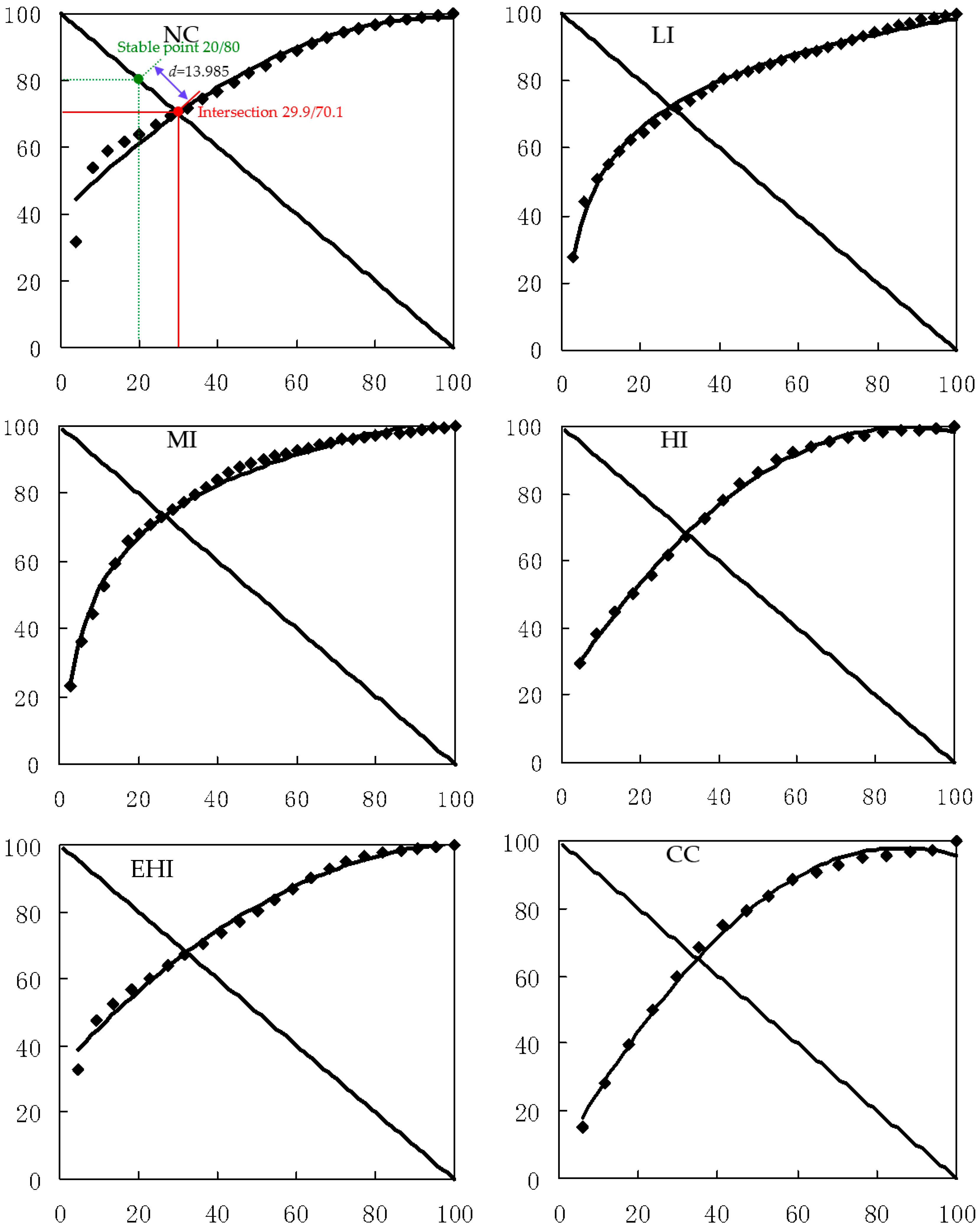
The relationship between community stability and species diversity has always been a complex theoretical issue of ecology. Most scholars have hypothesized that the higher species diversity resulted in higher forest community stability [
59,
60]. We obtained the same results in our study area. In our study, we found that the plant community was closer to stability after selective harvesting at low and medium intensities. But it was far from stable after clear-cut harvesting and selective harvesting at high and extra-high intensities. We tested the forest community stability based on the stability test method of improved Godron M., which made full use of the overall characteristics of the plant community and was a more comprehensive method including tree, shrub, and herb species. However, the forest community stability test could only determine whether the community was stable, but could not reveal community succession direction and trend [
44,
45]. Natural restoration of natural secondary forest after harvesting still exhibited dynamic change. The long-term tracking of permanent sample plots was very necessary to understand the long-term response mechanism of species diversity and stability to harvesting intensity. This measure was also needed to determine the relationship between species diversity and stability, as well as their mutual influencing mechanisms. Our study focuses on the impact of harvesting intensity on species diversity and community stability. Future studies should be conducted to obtain additional time series data. Coordinated multiple-regional studies can help explore the impact of other forcing such as environmental conditions together with timber harvesting intensity.
5. Conclusions
Results of this study showed that the plant species diversity indices of tree, shrub, and herb layers in this forest slightly increased with selective harvesting at low intensity. These parameters significantly increased with selective harvesting at medium intensity, but significantly decreased with selective harvesting at high and extra-high intensities, as well as with clear-cut harvesting. With increasing harvesting intensity, the richness, diversity, and evenness of the overall plants first increased, and then decreased, and these parameters achieved maxima with selective harvesting at medium intensity, but clear-cut harvesting reduced these parameters to their minima. The results of this study also showed that the plant community of this forest was closest to a stable community with selective harvesting at medium intensity. It was far from the stability with selective harvesting at high and extra-high intensities with clear-cut harvesting, and it was farthest from the stability with clear-cut harvesting.
With all the above impacts in mind, if diversity and community stability are prioritized, selective harvesting at low and medium intensities should be chosen in natural secondary forest management. Selective harvesting at medium intensity is the most favorable. However, clear-cut harvesting and selective harvesting at high and extra-high intensities should be avoided in this type of forest in this region.






{kind=link}
{kind=link}
{kind=link}