
Seedling Growth and Physiological Responses of Sixteen Eucalypt Taxa under Controlled Water Regime

Abstract
:1. Introduction
2. Material and Methods
2.1. Seedling Production and Experimental Site
2.2. Experimental Design
- -
- Irrigated (IRR): plants received daily irrigation of 400 mL to maintain at least 60% of the maximum water retention in the soil (soil moisture > 20%; Ψs < 0.02 bar), eliminating water limitation; and
- -
- Water limited (WL): irrigation was suspended until the occurrence of visual symptoms of stress (leaf wilting), when the plants were irrigated with 500 mL, corresponding to approximately 20% of soil capacity retention, repeating the cycle (soil moisture 3% to 10% v/v; 0.1 < Ψs < 1 bar).
2.3. Physiological and Biometric Measurements
2.4. Statistical Analysis
3. Results
4. Discussion
5. Conclusion
Acknowledgments
Author Contributions
Conflicts of Interests
References
- Gonçalves, J.L.M.; Stape, J.L.; Laclau, J.-P.; Smethurst, P.; Gava, J.L. Silvicultural effects on the productivity and wood quality of eucalypt plantations. For. Ecol. Manag. 2004, 193, 45–61. [Google Scholar] [CrossRef]
- Gonçalves, J.L.M.; Stape, J.L.; Laclau, J.P.; Bouillet, J.P.; Ranger, J. Assessing the effects of early silvicultural management on long-term site productivity of fast-growing eucalypt plantations: The Brazilian experience. South For. J. For. Sci. 2008, 70, 105–118. [Google Scholar] [CrossRef]
- Gonçalves, J.L.M.; Alvares, C.A.; Higa, A.R.; Silva, L.D.; Alfenas, A.C.; Stahl, J.; Ferraz, S.F.; de Paula, W.L.; Brancalion, P.H.S.; Hubner, A.; et al. Integrating genetic and silvicultural strategies to minimize abiotic and biotic constraints in Brazilian eucalypt plantations. For. Ecol. Manag. 2013, 301, 6–27. [Google Scholar] [CrossRef]
- Allen, C.D.; Macalady, A.K.; Chenchouni, H.; Bachelet, D.; McDowell, N.; Vennetier, M.; Kitzberger, T.; Rigling, A.; Breshears, D.D.; Hogg, E.H.; et al. A global overview of drought and heat-induced tree mortality reveals emerging climate change risks for forests. For. Ecol. Manag. 2010, 259, 660–684. [Google Scholar] [CrossRef]
- Florence, R.G. Ecology and Silviculture of Eucalypt Forest; Collingwood: CSIRO, Australia, 2004; p. 413. [Google Scholar]
- Ryan, M.G. Tree responses to drought. Tree Physiol. 2011, 31, 237–239. [Google Scholar] [CrossRef] [PubMed]
- Way, D.A.; Domec, J.-C.; Jackson, R. Elevated growth temperatures alter hydraulic characteristics in trembling aspen (Populus tremuloides) seedlings: implications for tree drought tolerance. Plant Cell Environ. 2012, 36, 103–115. [Google Scholar] [CrossRef] [PubMed]
- Battie-Laclau, P.; Laclau, J.-P.; Domec, J.-C.; Christina, M.; Bouillet, J.-P.; Piccolo, M.C.; Gonçalves, J.L.M.; Moreira, R.M.; Krusche, A.V.; Bouvet, J.-M.; et al. Effects of potassium and sodium supply on drought-adaptive mechanisms in Eucalyptus grandis plantations. New Phytol. 2014, 203, 401–413. [Google Scholar] [CrossRef] [PubMed]
- Granda, V.; Delatorre, C.; Cuesta, C.; Centeno, M.L.; Fernández, B.; Rodríguez, A.; Feito, I. Physiological and biochemical responses to severe drought stress of nine Eucalyptus globulus clones: A multivariate approach. Tree Physiol. 2014, 34, 778–786. [Google Scholar] [CrossRef] [PubMed]
- Breda, N.; Huc, R.; Granier, A.; Dreyer, E. Temperate forest trees and stands under severe drought: A review of ecophysiological responses, adaptation processes and long-term consequences. Ann. For. Sci. 2006, 63, 625–644. [Google Scholar] [CrossRef]
- McDowell, N.; Pockman, W.T.; Allen, C.D.; Breshears, D.D.; Cobb, N.; Kolb, T.; Plaut, J.; Sperry, J.; West, A.; Williams, D.G.; et al. Mechanisms of plant survival and mortality during drought: Why do some plants survive while others succumb to drought? New Phytol. 2008, 178, 719–739. [Google Scholar] [CrossRef] [PubMed]
- Ngugi, M.; Doley, D.; Hunt, M.; Ryan, P.; Dart, P. Physiological responses to water stress in Eucalyptus cloeziana and E. argophloia seedlings. Trees 2004, 18, 381–389. [Google Scholar] [CrossRef]
- Ladiges, P.Y. Variation in drought tolerance in Eucalyptus viminalis Labill. Aust. J. Bot. 1974, 22, 489–500. [Google Scholar] [CrossRef]
- Costa e Silva, F.; Shvaleva, A.; Maroco, J.P.; Almeida, M.H.; Chaves, M.M.; Pereira, J.S. Responses to water stress in two Eucalyptus globulus clones differing in drought tolerance. Tree Physiol. 2004, 24, 1165–1172. [Google Scholar] [CrossRef] [PubMed]
- Claeys, H.; Inzé, D. The agony of choice: How plants balance growth and survival under water-limiting conditions. Plant Physiol. 2013, 162, 1768–1779. [Google Scholar] [CrossRef] [PubMed]
- Harwood, C. New Introductions—Doing It Right In Developing a Eucalypt Resource: Learning from Australia and Elsewhere; Walker, J., Ed.; Wood Technology Research Centre, University of Canterbury: Christchurch, New Zealand, 2011; pp. 125–136. [Google Scholar]
- Potts, B.M.; Dungey, H.S. Hybridisation of Eucalyptus: Key issues for breeders and geneticists. New For. 2004, 27, 115–138. [Google Scholar] [CrossRef]
- Dungey, H.S.; Potts, B.M.; Carnegie, A.J.; Ades, P.K. Mycosphaerella leaf disease: Genetic variation in damage to Eucalyptus nitens, Eucalyptus globulus, and their F1 hybrid. Can J. For. Res. 1997, 27, 750–759. [Google Scholar] [CrossRef]
- Reis, G.G.; Reis, M.D.G.F.; Fontan, I.D.C.I.; Monte, M.A.; Gomes, A.N.; de Oliveira, C.H.R. Crescimento de raízes e da parte aérea de clones de híbridos de Eucalyptus grandis x Eucalyptus urophylla e de Eucalyptus camaldulensis x Eucalyptus spp submetidos a dois regimes de irrigação no campo. R Árvore 2006, 30, 921–931. [Google Scholar] [CrossRef]
- Alvares, C.A.; Stape, J.L.; Sentelhas, P.C.; Gonçalves, J.L.M.; Sparovek, G. Köppen’s climate classification map for Brazil. Meteorol Z 2013, 22, 711–728. [Google Scholar] [CrossRef]
- Sarruge, J.R. Soluções nutritivas. Summa Phytop 1975, 1, 231–233. [Google Scholar]
- Stape, J.; Binkley, D.; Ryan, M. Eucalyptus production and the supply, use and efficiency of use of water, light and nitrogen across a geographic gradient in Brazil. For. Ecol. Manag. 2004, 193, 17–31. [Google Scholar] [CrossRef]
- Stape, J.L.; Binkley, D.; Ryan, M.G.; Fonseca, S.; Loos, R.A.; Takahashi, E.N.; Silva, C.R.; Silva, S.R.; Hakamada, R.E.; Ferreira, J.M.; et al. The Brazil Eucalyptus Potential Productivity Project: Influence of water, nutrients and stand uniformity on wood production. For. Ecol. Manag. 2010, 259, 1684–1694. [Google Scholar] [CrossRef]
- Jaleel, C.A.; Manivannan, P.; Wahid, A.; Farooq, M.; Somasundaram, R.; Panneerselvam, R. Drought stress in plants: A review on morphological characteristics and pigments composition. Int. J. Agric. Biol. 2009, 11, 100–105. [Google Scholar]
- Myers, B.J.; Landsberg, J.J. Water stress and seedling growth of two eucalypt species from contrasting habitats. Tree Physiol. 1989, 5, 207–218. [Google Scholar] [CrossRef] [PubMed]
- Bauerle, W.L.; Wang, G.G.; Bowden, J.D.; Hong, C.M. An analysis of ecophysiological responses to drought in American Chestnut. Ann. For. Sci. 2006, 63, 833–842. [Google Scholar] [CrossRef]
- Christina, M.; Le Maire, G.; Battie-Laclau, P.; Nouvellon, Y.; Bouillet, J.-P.; Jourdan, C.; de Moraes Gonçalves, J.L.; Laclau, J.-P. Measured and modeled interactive effects of potassium deficiency and water deficit on gross primary productivity and light-use efficiency in Eucalyptus grandis plantations. Glob. Chang. Biol. 2015, 21, 2022–2039. [Google Scholar] [CrossRef] [PubMed]
- Chaves, M.M.; Pereira, J.S.; Maroco, J.; Rodrigues, M.L.; Ricardo C, P.P.; Osório, M.L.; Pinheiro, C. How plants cope with water stress in the field? Photosynthesis and growth. Ann. Bot. 2002, 89, 907–916. [Google Scholar] [CrossRef] [PubMed]
- Litton, C.M.; Raich, J.W.; Ryan, M.G. Carbon allocation in forest ecosystems. Glob. Change Biol. 2007, 13, 2089–2109. [Google Scholar] [CrossRef]
- Ryan, M.G.; Stape, J.L.; Binkley, D.; Fonseca, S.; Loos, R.A.; Takahashi, E.N.; Silva, C.R.; Silva, S.R.; Hakamada, R.E.; Ferreira, J.M.; et al. Factors controlling Eucalyptus productivity: How resource availability and stand structure alter production and carbon allocation. For. Ecol. Manag. 2010, 259, 1695–1703. [Google Scholar] [CrossRef]
- Ochieng, J.W.; Shepherd, M.; Baverstock, P.R.; Nikles, G.; Lee, D.J.; Henry, R.J. Two sympatric spotted gum species are molecularly homogeneous. Conserv. Genet 2010, 11, 45–56. [Google Scholar] [CrossRef]
- Wang, D.; Bachelard, E.P.; Banks, J.C.G. Growth and water relations of seedlings of two subspecies of Eucalyptus globulus. Tree Physiol. 1988, 4, 129–138. [Google Scholar] [CrossRef] [PubMed]
- Teskey, R.O.; Bongarten, B.C.; Cregg, B.M.; Dougherty, P.M.; Hennessey, T.C. Physiology and genetics of tree growth response to moisture and temperature stress: an examination of the characteristics of loblolly pine (Pinus taeda L.). Tree Physiol. 1987, 3, 41–61. [Google Scholar] [CrossRef] [PubMed]
- Valentini, R.; Mugnozza, G.S.; Giordano, E.; Kuzminsky, E. Influence of cold hardening on water relations of three Eucalyptus species. Tree Physiol. 1990, 6, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Choat, B.; Medek, D.E.; Stuart, S.A.; Pasquet-Kok, J.; Egerton, J.J.G.; Salari, H.; Sack, L.; Ball, M. Xylem traits mediate a trade-off between resistance to freeze–thaw-induced embolism and photosynthetic capacity in overwintering evergreens. New Phytol. 2011, 191, 996–1005. [Google Scholar] [CrossRef] [PubMed]
- Taiz, L.; Zeiger, E. Plant Physiology, 5th ed.; Sinauer Associates Inc: Sunderland, UK, 2010. [Google Scholar]
- Parra-O, C.; Bayly, M.J.; Drinnan, A.; Udovicic, F.; Ladiges, P. Phylogeny, major clades and infrageneric classification of Corymbia (Myrtaceae), based on nuclear ribosomal DNA and morphology. Aust. Syst. Bot. 2010, 22, 384–399. [Google Scholar] [CrossRef]
- Cernusak, L.A.; Hutley, L.B.; Beringer, J.; Holtum, J.A.; Turner, B.L. Photosynthetic physiology of eucalypts along a sub-continental rainfall gradient in northern Australia. Agric. For. Meteorol. 2011, 151, 1462–1470. [Google Scholar] [CrossRef]
- Merchant, A.; Callister, A.; Arndt, S.; Tausz, M.; Adams, M.A. Contrasting physiological responses of six Eucalyptus species to water deficit. Ann. Bot. 2007, 100, 1507–1515. [Google Scholar] [CrossRef] [PubMed]
- White, D.A.; Beadle, C.L.; Worledge, D.; Honeysett, J.L. Wood production per evapotranspiration was increased by irrigation in plantations of Eucalyptus globulus and E. nitens. New For. 2016, 47, 303–317. [Google Scholar] [CrossRef]
- Silva, P.H.M.; Campoe, O.C.; Vieira, I.G.; Paula, R.C.D. Aplicação foliar de boro em eucalipto sob estresse hídrico. Scientia Forestalis 2015, 43, 395–405. [Google Scholar]
- Lemcoff, J.H.; Guarnaschelli, A.B.; Garau, A.M.; Prystupa, P. Elastic and osmotic adjustments in rooted cuttings of several clones of Eucalyptus camaldulensis Dehnh. from southeastern Australia after a drought. Flora 2002, 197, 134–142. [Google Scholar] [CrossRef]
- Silva, P.H.M.; Paula, R.C.; Miranda, A.C.; Moraes, M.L.T.; Furtado, E.L.; de Pieri, C.; Brawner, J.; Lee, D. Screening of rust and gall wasp in eucalypts species and provenances. Pesq Flor Brasil 2015, 35, 323–328. [Google Scholar] [CrossRef]
- Mendel, Z.; Protasov, A.; Fisher, N.; La Salle, J. Taxonomy and Biology of Leptocybe invasa (Eulophidae: Hymenoptera) an invasive gall inducer on eucalyptus. Aust. J. Entomol. 2004, 43, 101–113. [Google Scholar] [CrossRef]
- Waraich, E.A.; Ahmad, R.; Ashraf, M.Y. Role of mineral nutrition in alleviation of drought stress in plants. Aust. J. Crop. Sci. 2011, 5, 764. [Google Scholar]



{kind=link}
{kind=link}
| Species | Abbreviation | Section | Origin | Seed lot | Provenance-Source | AAT (°C) | Annual Rainfall (mm) | Koppen |
|---|---|---|---|---|---|---|---|---|
| Corymbia. Citriodora (Hook) subsp. variegata F. Muell | CCV 1 | CSIRO | 19,664 | Barakula, AU (WS) | 21.6 | 656 | BSh | |
| Corymbia citriodora subsp. variegata | CCV 2 | CSIRO | 20,787 | Barclays Deniliquin, AU (SO) | 18.7 | 394 | BSk | |
| Corymbia torelliana F. Muell | CT | IPEF | AN0255N01 | Anhembi, BR (SO) | 22.2 | 1305 | Aw/Cwa | |
| Corymbia henryi S.T. Blake | CH | DAFF | 10,250 | Lockyer, AU (WS) | 21.2 | 820 | CFa | |
| Eucalyptus argophloia Blakely | E. argo | Symphyomyrtus Adnataria | DAFF | 12,716 | Dalby, AU (SO) | 20.8 | 676 | BSh |
| Eucalyptus amplifolia Brooker & A. Slee | E.ampl 1 | Symphyomyrtus Exsertaria | CSIRO | 15,281 | Nerong S.F (WS) | 19.2 | 1321 | Cfb |
| Eucalyptus amplifolia | E. ampl 2 | Symphyomyrtus Exsertaria | CSIRO | 18,731 | Clouds CK SF & TSR (WS) | 16.5 | 1434 | Cfb |
| Eucalyptus crebra F. Muell | E. crebra | Symphyomyrtus Adnataria | CSIRO | 15,146 | NW Baradine, AU (WS) | 18.9 | 747 | BSh |
| Eucalyptus moluccana Roxb. | E. molu | Symphyomyrtus Adnataria | CSIRO | 20,010 | Crediton, AU (WS) | 22.0 | 1539 | BSh |
| Eucalyptus brassiana S.T. Blake | E. bras | Symphyomyrtus Exsertaria | IPEF | US002N01 | Urbano Santos, BR (SO) | 27.0 | 1700 | Aw |
| Eucalyptus camaldulensis Dehnh | E. camal | Symphyomyrtus Exsertaria | IPEF | SE007N01 | Selviria, BR (SO) | 23.6 | 1300 | Aw/Cwa |
| Eucalyptus brookeriana A. M. Gray | E. brook | Symphyomyrtus Maidenaria | CSIRO | 18,317 | Otways, AU (WS) | 12.9 | 1539 | Cs |
| Eucalyptus macarthurii Deane & Maiden | E. maca | Symphyomyrtus Maidenaria | CSIRO | 20,897 | Paddys River, AU (WS) | 12.3 | 1248 | Cfb |
| Eucalyptus longirostrata (Blakely) L. Johnson & K. Hill | E. long | Symphyomyrtus Latoangulate | DAFF | 20,464 | Coominglah, AU (WS) | 22.0 | 690 | Cfa |
| Eucalyptus urophylla S.T. Blake | GG100 * | Symphyomyrtus Latoangulate | Gerdau/Vallourec | Clone GG 100 | NA | NA | NA | NA |
| Eucalyptus urophylla x E. camaldulensis | VM1 * | Symp. Lateangulate x Exsertaria | Vallourec | Clone VM 1 | NA | NA | NA | NA |
| Source of Variation | Total Height | Basal Stem Diameter | Leaf Area | Specific Leaf Area | Biomass | E | A | gs | WUEi | |||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Root | Stem | Leaves | Total | |||||||||
| Taxa | <0.0001 | <0.0001 | <0.0001 | <0.0001 | <0.0001 | <0.0001 | <0.0001 | <0.0001 | <0.0001 | <0.0001 | <0.0001 | <0.0001 |
| Water availability | <0.0001 | <0.0001 | <0.0001 | 0.613 | <0.0001 | <0.0001 | 0.0361 | <0.0001 | <0.0001 | <0.0001 | <0.0001 | <0.0001 |
| Taxa x Water availability | <0.0001 | 0.068 | 0.0876 | 0.125 | 0.0161 | 0.0006 | 0.2528 | 0.0216 | 0.0006 | 0.4914 | 0.9114 | 0.968 |
| Taxa | Total Height cm | Basal Stem Diameter cm | Number of Leaves Leaf Tree−1 | Leaf Area cm2·Tree−1 | Specific Leaf Area m2·kg−1 | Shoot Biomass g Tree−1 | Root Biomass g Tree−1 | Total Biomass g Tree−1 | ||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| IRR | WL | IRR | WL | IRR | WL | IRR | WL | IRR | WL | IRR | WL | IRR | WL | IRR | WL | |||||||||||||||||
| CCV 1 | 76.7 | A | 59.7 | A | 7.9 | B | 6.2 | B | 48 | F | 35 | C | 946 | B | 490 | B | 11.9 | A | 8.1 | B | 18.0 | B | 10.6 | B | 7.0 | B | 3.4 | C | 25.0 | B | 14.0 | B |
| CCV 2 | 73.3 | A | 49.9 | B | 8.5 | B | 6.1 | B | 30 | F | 32 | C | 1158 | A | 836 | A | 7.3 | B | 7.2 | B | 24.6 | A | 15.6 | A | 8.0 | A | 5.3 | B | 32.5 | A | 20.9 | A |
| CT | 73.0 | A | 64.3 | A | 9.2 | A | 7.8 | A | 39 | F | 24 | C | 1081 | B | 880 | A | 9.9 | A | 8.9 | A | 21.9 | B | 17.4 | A | 7.8 | A | 5.4 | B | 29.7 | A | 22.9 | A |
| CH | 64.3 | B | 53.3 | B | 7.4 | C | 6.6 | B | 23 | F | 18 | C | 1043 | B | 707 | B | 7.5 | B | 6.5 | B | 21.3 | B | 15.8 | A | 6.9 | B | 5.1 | B | 28.1 | B | 20.8 | A |
| E. argo | 73.9 | A | 61.1 | A | 6.4 | C | 5.9 | B | 662 | A | 344 | A | 925 | B | 588 | B | 6.3 | B | 7.5 | B | 24.6 | A | 14.5 | B | 6.0 | B | 3.8 | C | 30.7 | A | 18.3 | B |
| E.ampl 1 | 66.7 | B | 49.0 | B | 6.6 | C | 5.8 | B | 36 | F | 20 | C | 1189 | A | 684 | B | 5.8 | B | 5.4 | B | 28.0 | A | 16.8 | A | 5.1 | B | 3.7 | C | 33.1 | A | 20.4 | A |
| E. ampl 2 | 69.4 | B | 63.8 | A | 9.2 | A | 8.5 | A | 47 | F | 40 | C | 1132 | A | 872 | A | 9.9 | A | 8.6 | B | 21.1 | B | 16.2 | A | 9.3 | A | 8.2 | A | 30.4 | A | 24.4 | A |
| E. crebra | 67.3 | B | 51.1 | B | 6.2 | C | 5.2 | B | 437 | B | 320 | A | 817 | B | 458 | B | 6.0 | B | 4.9 | B | 21.3 | B | 14.2 | B | 5.9 | B | 3.9 | C | 27.2 | B | 18.1 | B |
| E. molu | 59.5 | B | 57.9 | A | 8.7 | A | 6.5 | B | 110 | E | 99 | C | 1388 | A | 721 | B | 8.2 | B | 12.6 | A | 24.7 | A | 13.0 | B | 8.9 | A | 5.0 | B | 33.7 | A | 18.0 | B |
| E. bras | 84.6 | A | 65.4 | A | 7.9 | B | 6.8 | B | 88 | E | 73 | C | 1081 | B | 682 | B | 10.3 | A | 9.8 | A | 19.7 | B | 12.6 | B | 7.6 | A | 4.6 | B | 27.3 | B | 17.1 | B |
| E. camal | 77.8 | A | 64.0 | A | 9.1 | A | 7.5 | A | 63 | F | 46 | C | 997 | B | 606 | B | 8.2 | B | 7.6 | B | 21.3 | B | 13.8 | B | 9.4 | A | 7.4 | A | 30.7 | A | 21.3 | A |
| E. brook | 62.6 | B | 62.0 | A | 6.3 | C | 6.3 | B | 238 | D | 220 | B | 923 | B | 783 | A | 9.4 | A | 9.5 | A | 17.4 | B | 15.3 | A | 4.3 | B | 2.6 | C | 21.8 | B | 18.0 | B |
| E. maca | 63.4 | B | 57.6 | A | 6.7 | C | 5.8 | B | 317 | C | 255 | B | 1320 | A | 1003 | A | 10.0 | A | 12.9 | A | 21.3 | B | 13.9 | B | 7.4 | A | 4.7 | B | 28.6 | A | 18.6 | B |
| E. long | 77.9 | A | 62.4 | A | 9.2 | A | 8.4 | A | 214 | D | 182 | B | 827 | B | 586 | B | 10.0 | A | 11.2 | A | 17.7 | B | 11.8 | B | 6.6 | B | 4.9 | B | 24.3 | B | 16.6 | B |
| GG100 | 63.5 | B | 58.3 | A | 7.6 | C | 6.3 | B | 83 | E | 69 | C | 1461 | A | 976 | A | 11.2 | A | 10.1 | A | 21.1 | B | 16.1 | A | 7.7 | A | 5.5 | B | 28.8 | A | 21.6 | A |
| VM1 | 68.4 | B | 60.5 | A | 7.1 | C | 5.8 | B | 88 | E | 64 | C | 1273 | A | 678 | B | 11.1 | A | 9.1 | A | 19.9 | B | 13.5 | B | 7.0 | B | 4.0 | C | 26.9 | B | 17.4 | B |
| Average | 70.1 | 58.8 | 7.8 | 6.6 | 158 | 115 | 1098 | 722 | 8.9 | 8.7 | 21.5 | 14.4 | 7.2 | 4.8 | 28.7 | 19.3 | ||||||||||||||||
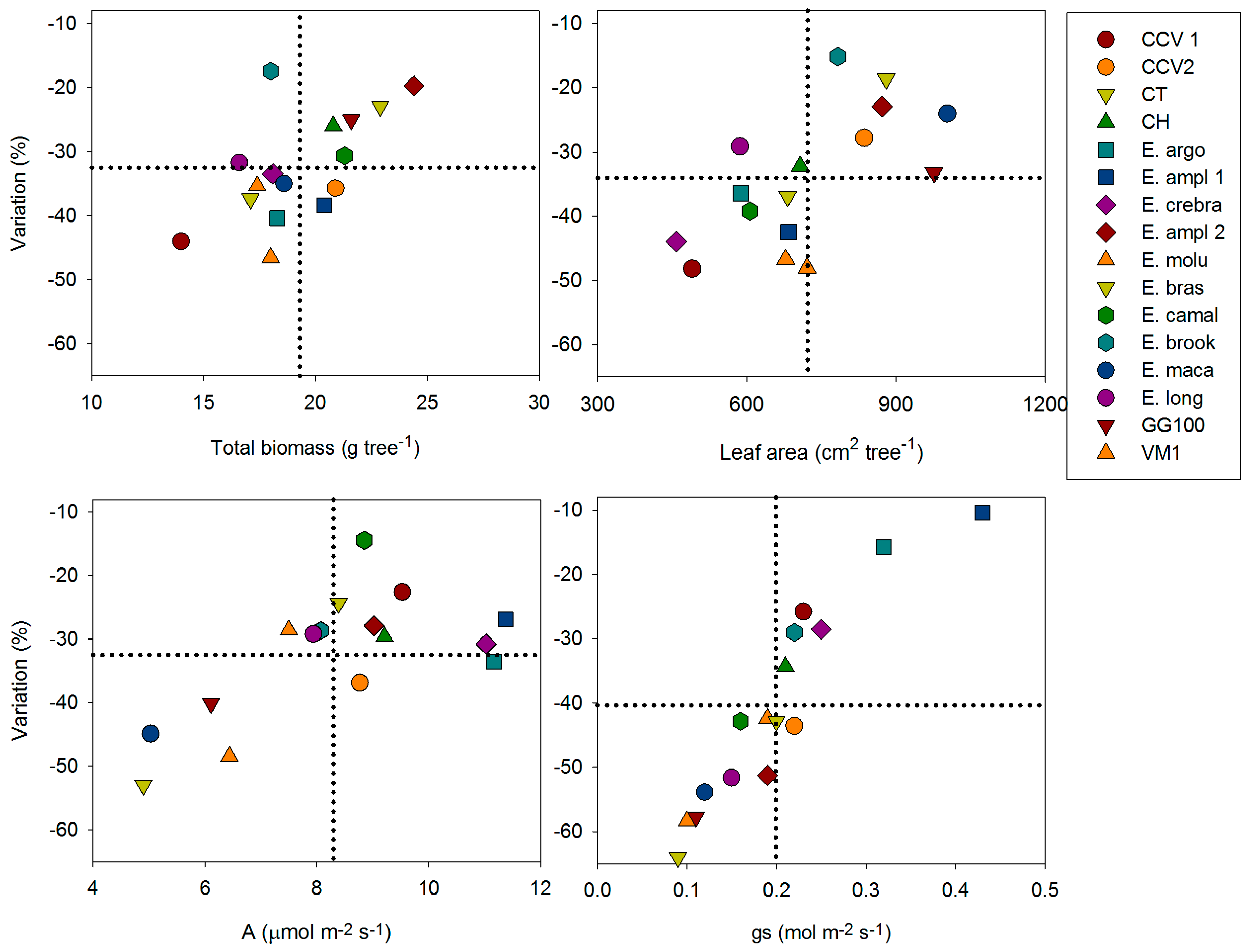
| Variation (%) | −16 | −15 | −27 | −34 | −2 | −33 | −33 | −33 | ||||||||||||||||||||||||
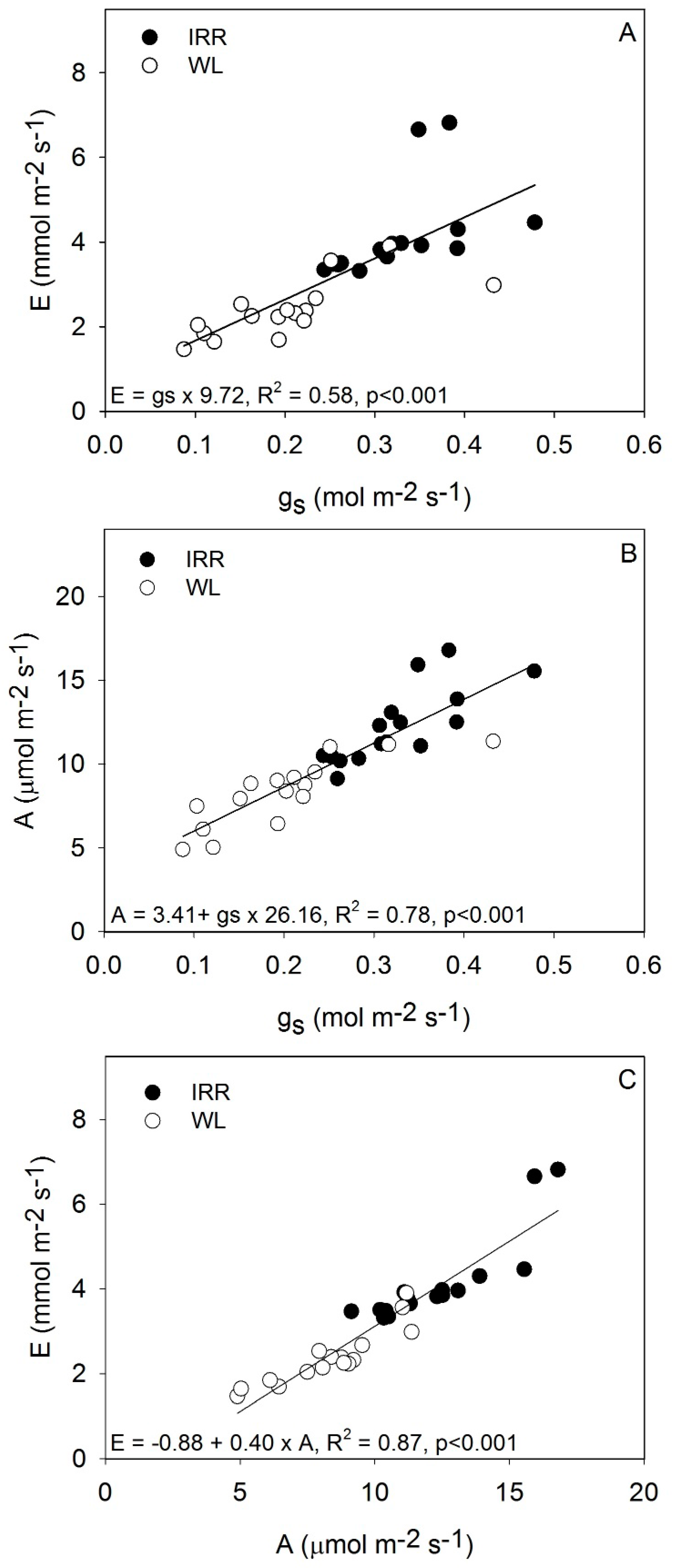
| Taxa | Transpiration mmol m−2·s−1 | Stomatal Conductance mol m−2·s−1 | Photosynthesis µmol m−2·s−1 | WUEi µmol mmol−1 | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| IRR | WL | IRR | WL | IRR | WL | IRR | WL | |||||||||
| CCV 1 | 3.82 | B | 2.67 | B | 0.31 | B | 0.23 | B | 12.31 | B | 9.53 | B | 3.27 | A | 3.62 | A |
| CCV 2 | 4.30 | B | 2.38 | B | 0.39 | A | 0.22 | B | 13.89 | A | 8.77 | B | 3.25 | A | 3.73 | A |
| CT | 3.48 | B | 1.47 | C | 0.25 | B | 0.09 | B | 10.41 | B | 4.90 | C | 3.00 | A | 3.37 | B |
| CH | 3.96 | B | 2.32 | B | 0.32 | B | 0.21 | B | 13.09 | B | 9.21 | B | 3.32 | A | 4.00 | A |
| E. argo | 6.82 | A | 3.91 | A | 0.38 | A | 0.32 | A | 16.80 | A | 11.17 | A | 2.44 | B | 2.82 | B |
| E.ampl 1 | 4.46 | B | 2.98 | A | 0.48 | A | 0.43 | A | 15.55 | A | 11.37 | A | 3.48 | A | 3.92 | A |
| E. ampl 2 | 3.85 | B | 2.23 | B | 0.39 | A | 0.19 | B | 12.51 | B | 9.02 | B | 3.26 | A | 4.06 | A |
| E. crebra | 6.66 | A | 3.56 | A | 0.35 | A | 0.25 | B | 15.93 | A | 11.03 | A | 2.42 | B | 3.12 | B |
| E. molu | 3.97 | B | 1.69 | C | 0.33 | B | 0.19 | B | 12.50 | B | 6.44 | C | 3.16 | A | 3.77 | A |
| E. bras | 3.92 | B | 2.39 | B | 0.35 | A | 0.20 | B | 11.09 | B | 8.39 | B | 2.82 | B | 3.57 | A |
| E. camal | 3.32 | B | 2.25 | B | 0.28 | B | 0.16 | B | 10.34 | B | 8.85 | B | 3.12 | A | 3.99 | A |
| E. brook | 3.66 | B | 2.14 | B | 0.31 | B | 0.22 | B | 11.31 | B | 8.07 | B | 3.11 | A | 3.83 | A |
| E. maca | 3.47 | B | 1.65 | C | 0.26 | B | 0.12 | B | 9.13 | B | 5.03 | C | 2.63 | B | 3.00 | B |
| E. long | 3.78 | B | 2.53 | B | 0.31 | B | 0.15 | B | 11.21 | B | 7.94 | B | 2.96 | A | 3.20 | B |
| GG100 | 3.50 | B | 1.84 | C | 0.26 | B | 0.11 | B | 10.20 | B | 6.11 | C | 2.95 | A | 3.27 | B |
| VM1 | 3.35 | B | 2.04 | B | 0.24 | B | 0.10 | B | 10.50 | B | 7.50 | C | 3.14 | A | 3.70 | A |
| Average | 4.15 | 2.38 | 0.33 | 0.20 | 12.30 | 8.33 | 3.02 | 3.56 | ||||||||
| Variation (%) | −43 | −39 | −32 | +18 | ||||||||||||
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Silva, P.H.M.; Campoe, O.C.; De Paula, R.C.; Lee, D.J. Seedling Growth and Physiological Responses of Sixteen Eucalypt Taxa under Controlled Water Regime. Forests 2016, 7, 110. https://doi.org/10.3390/f7060110
Silva PHM, Campoe OC, De Paula RC, Lee DJ. Seedling Growth and Physiological Responses of Sixteen Eucalypt Taxa under Controlled Water Regime. Forests. 2016; 7(6):110. https://doi.org/10.3390/f7060110
Chicago/Turabian StyleSilva, Paulo H. M., Otavio C. Campoe, Rinaldo C. De Paula, and David J. Lee. 2016. "Seedling Growth and Physiological Responses of Sixteen Eucalypt Taxa under Controlled Water Regime" Forests 7, no. 6: 110. https://doi.org/10.3390/f7060110