
The Speed of Invasion: Rates of Spread for Thirteen Exotic Forest Insects and Diseases


Abstract
:1. Introduction
1.1. Insects
1.2. Diseases
2. Methods
2.1. Geospatial Databases
2.2. Analytical Methods
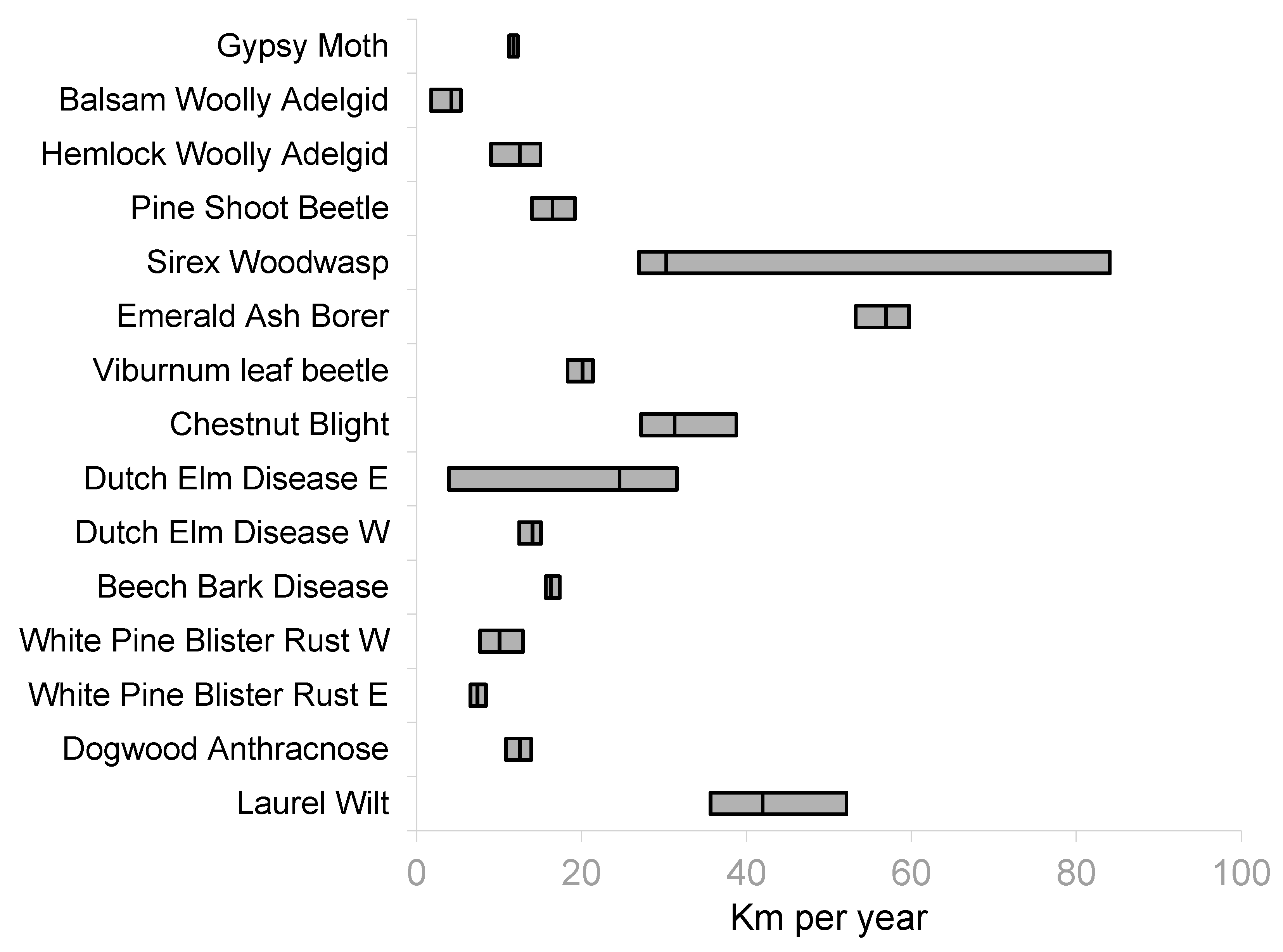
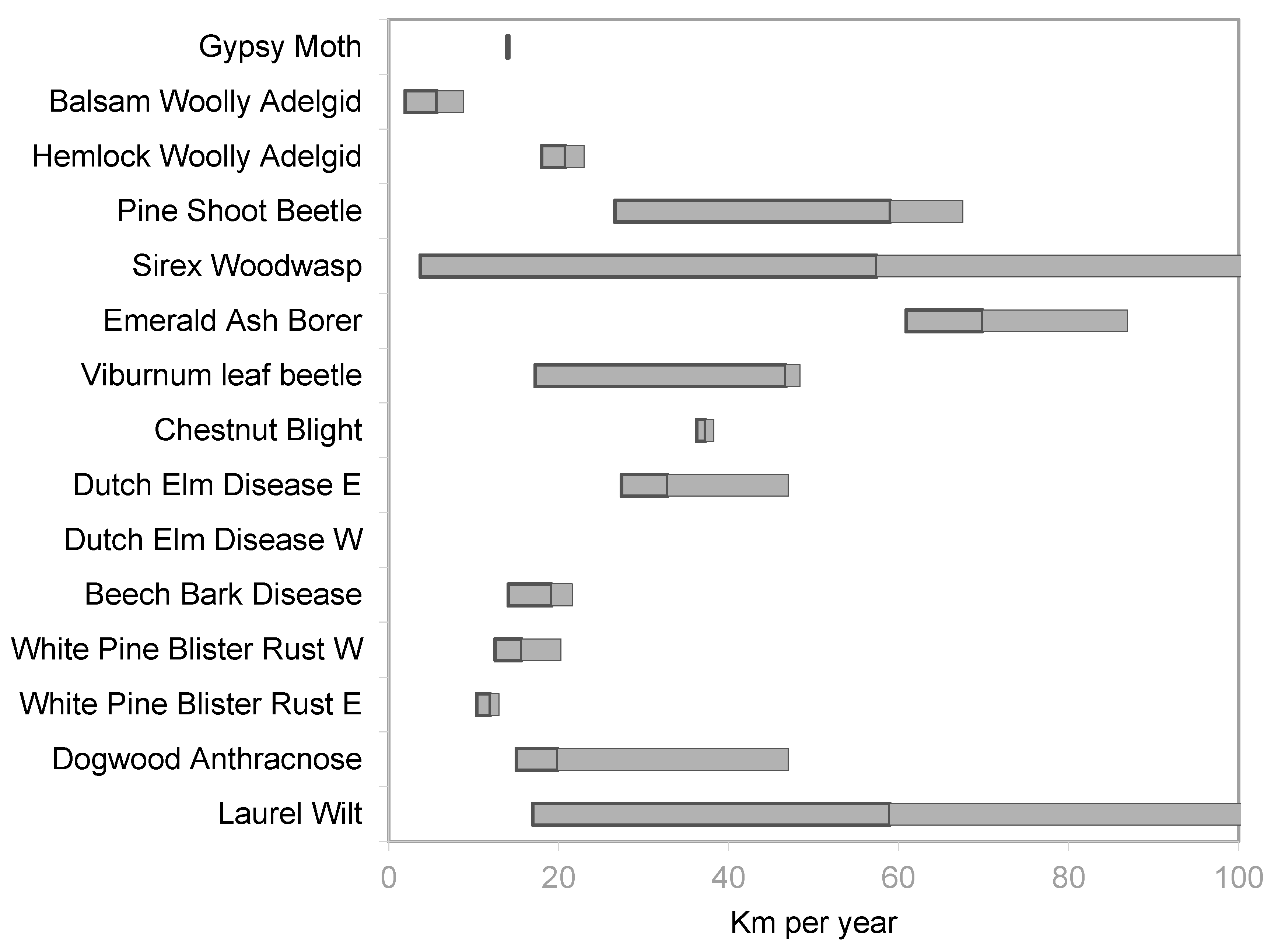
3. Results
4. Discussion
5. Conclusions
Acknowledgments
Conflicts of Interest
References
- Peltzer, D.A.; Allen, R.B.; Lovett, G.M.; Whitehead, D.; Wardle, D.A. Effects of biological invasions on forest carbon sequestration. Glob. Chang. Biol. 2010, 16, 732–746. [Google Scholar] [CrossRef]
- Albani, M.; Moorcroft, P.R.; Ellison, A.M.; Orwig, D.A.; Foster, D.R. Predicting the impact of hemlock woolly adelgid on carbon dynamics of eastern united states forests. Can. J. For. Res. 2010, 40, 119–133. [Google Scholar] [CrossRef]
- Bradley, B.; Wilcove, D.; Oppenheimer, M. Climate change increases risk of plant invasion in the Eastern United States. Biol. Invasions 2010, 12, 1855–1872. [Google Scholar] [CrossRef]
- Diez, J.M.; D’Antonio, C.M.; Dukes, J.S.; Grosholz, E.D.; Olden, J.D.; Sorte, C.J.B.; Blumenthal, D.M.; Bradley, B.A.; Early, R.; Ibáñez, I.; et al. Will extreme climatic events facilitate biological invasions? Front. Ecol. Environ. 2012, 10, 249–257. [Google Scholar] [CrossRef] [Green Version]
- Kasson, M.T.; Livingston, W.H. Relationships among beech bark disease, climate, radial growth response and mortality of American beech in northern Maine, USA. For. Pathol. 2012, 42, 199–212. [Google Scholar] [CrossRef]
- Murphy, S.D. Concurrent management of an exotic species and initial restoration efforts in forests. Restor. Ecol. 2005, 13, 584–593. [Google Scholar] [CrossRef]
- Holmes, T.P.; Aukema, J.E.; Holle, B.V.; Liebhold, A.; Sills, E. Economic impacts of invasive species in forests. Ann. N. Y. Acad. Sci. 2009, 1162, 18–38. [Google Scholar] [CrossRef] [PubMed]
- Aukema, J.E.; Leung, B.; Kovacs, K.; Chivers, C.; Britton, K.O.; Englin, J.; Frankel, S.J.; Haight, R.G.; Holmes, T.P.; Liebhold, A.M.; et al. Economic impacts of non-native forest insects in the continental united states. PLoS ONE 2011, 6, e24587. [Google Scholar] [CrossRef] [PubMed]
- Marchetto, K.M.; Shea, K.; Kelly, D.; Groenteman, R.; Sezen, Z.; Jongejans, E. Unrecognized impact of a biocontrol agent on the spread rate of an invasive thistle. Ecol. Appl. 2014, 24, 1178–1187. [Google Scholar] [CrossRef] [PubMed]
- Yemshanov, D.; Koch, F.H.; Ben-Haim, Y.; Smith, W.D. Robustness of risk maps and survey networks to knowledge gaps about a new invasive pest. Risk Anal. 2010, 30, 261–276. [Google Scholar] [CrossRef] [PubMed]
- Ayres, M.P.; Lombardero, M.J. Assessing the consequences of global change for forest disturbance from herbivores and pathogens. Sci. Total Environ. 2000, 262, 263–286. [Google Scholar] [CrossRef]
- Gray, D.R. The gypsy moth life stage model: Landscape-wide estimates of gypsy moth establishment using a multi-generational phenology model. Ecol. Model. 2004, 176, 155–171. [Google Scholar] [CrossRef]
- Evans, A.M.; Finkral, A.J. A new look at spread rates of exotic diseases in north american forests. For. Sci. 2010, 56, 453–459. [Google Scholar]
- Evans, A.M.; Gregoire, T.G. A geographically variable model of hemlock woolly adelgid spread. Biol. Invasions 2007, 9, 369–382. [Google Scholar] [CrossRef]
- Liebhold, A.M.; Halverson, J.; Elmes, G. Quantitative analysis of the invasion of gypsy moth in North America. J. Biogeogr. 1992, 19, 513–520. [Google Scholar] [CrossRef]
- Balch, R.E. Spread of Balsam Woolly Aphid (Adelges piceae) in relation to climate of Newfoundland. In Bi-Monthly Progress Report; Department of Agriculture, Science Service Forest Biology Division: Ottawa, ON, Canada, 1954; Volume 10, p. 5. [Google Scholar]
- Havill, N.P.; Montgomery, M.E. The role of arboreta in studying the evolution of host resistance to the hemlock woolly adelgid. Arnoldia 2008, 65, 1–9. [Google Scholar]
- Barak, A.V.; McGervy, D.; Tokaya, G. Dispersal and re-capture of marked, overwintering Tomicus piniperda (coleoptera: Scolytidae) from scotch pine bolts. Gt. Lakes Entomol. 2000, 33, 81–105. [Google Scholar]
- Carnegie, A.J.; Matsuki, M.; Haugen, D.A.; Hurley, B.P.; Ahumada, R.; Klasmer, P.; Sun, J.; Iede, E.T. Predicting the potential distribution of sirex noctilio (hymenoptera: Siricidae), a significant exotic pest of pinus plantations. Ann. For. Sci. 2006, 63, 119–128. [Google Scholar] [CrossRef]
- Bray, A.; Bauer, L.; Poland, T.; Haack, R.; Cognato, A.; Smith, J. Genetic analysis of emerald ash borer (Agrilus planipennis fairmaire) populations in Asia and North America. Biol. Invasions 2011, 13, 2869–2887. [Google Scholar] [CrossRef]
- Weston, P.A.; Hoebeke, E.R. Viburnum leaf beetle, Pyrrhalta viburni (paykull) (coleoptera: Chrysomelidae): Dispersal pattern of a palearctic landscape pest in New York and its distribution status in the northeastern U.S. and eastern Canada. Proc. Entomol. Soc. Wash. 2003, 105, 4105–4107. [Google Scholar]
- Gravatt, G.F. Chestnut blight in North America. Unasylva 1949, 3, 2–7. [Google Scholar]
- Gibbs, J.N. Intercontinental epidemiology of Dutch elm disease. Annu. Rev. Phytopathol. 1978, 16, 287–307. [Google Scholar] [CrossRef]
- Houston, D.R. Major new tree disease epidemics: Beech bark disease. Annu. Rev. Phytopathol. 1994, 32, 75–87. [Google Scholar] [CrossRef]
- Gibbs, J.N.; Wainhouse, D. Spread of forest pests and pathogens in the northern hemisphere. Forestry 1986, 59, 141–153. [Google Scholar] [CrossRef]
- Maloy, O.C. White pine blister rust control in North America: A case history. Annu. Rev. Phytopathol. 1997, 35, 87–109. [Google Scholar] [CrossRef] [PubMed]
- Miller, D.R.; Kimmey, J.W.; Fowler, M.E. White Pine Blister Rust; Forest Pest Leaflet 36; USDA Forest Service: Washington, DC, USA, 1959; p. 8.
- Hibben, C.; Daughtrey, M. Dogwood anthracnose in Northeastern United States. Plant Dis. 1988, 72, 199–203. [Google Scholar] [CrossRef]
- Harrington, T.; Fraedrich, S.; Aghayeva, D. Raffaelea lauricola, a new ambrosia beetle symbiont and pathogen on the lauraceae. Mycotaxon 2008, 104, 399–404. [Google Scholar]
- Tobin, P.C.; Liebhold, A.M.; Roberts, E.A. Comparison of methods for estimating the spread of a non-indigenous species. J. Biogeogr. 2007, 34, 305–312. [Google Scholar] [CrossRef]
- ESRI. Arcgis, version 9.3; Environmental Systems Research Institute (ESRI): Redlands, CA, USA, 2009. [Google Scholar]
- Sas Onlinedoc, version 9; SAS Institute Inc.: Cary, NC, USA, 2002.
- Gannoun, A.; Saracco, J.; Yuan, A.; Bonney, G. Non-parametric quantile regression with censored data. Scand. J. Stat. 2005, 32, 527–550. [Google Scholar] [CrossRef]
- Cade, B.; Noon, B. A gentle introduction to quantile regression for ecologists. Front. Ecol. Environ. 2003, 1, 412–420. [Google Scholar] [CrossRef]
- Cade, B.S.; Terrell, J.W.; Schroeder, R.L. Estimating effects of limiting factors with regression quantiles. Ecology 1999, 80, 311–323. [Google Scholar] [CrossRef]
- Liebhold, A.M. An Atlas of Historical Gypsy Moth Defoliation & Quarantined Areas in the US. Available online: http://www.fs.fed.us/ne/morgantown/4557/gmoth/atlas/#spread (accessed on 8 June 2013).
- Pest Tracker. Available online: http://pest.ceris.purdue.edu/ (accessed on 8 June 2013).
- Greenbank, D.O. Climate and the ecology of the balsam woolly aphid. Can. Entomol. 1970, 102, 546–578. [Google Scholar] [CrossRef]
- USDA Forest Service. Pine Shoot Beetle, 1992–2001. Available online: http://www.ncrs.fs.fed.us/4153/deltawest/plantanimal/pineshoot.asp (accessed on 8 June 2013).
- Plant Health Surveillance Unit. Plant Protection Survey Report. Available online: http://epe.lac-bac.gc.ca/100/206/301/cfia-acia/2011-09-21/www.inspection.gc.ca/english/plaveg/pestrava/surv/sit2008e.shtml (accessed on 8 June 2013).
- USDA Forest Service. Alien Forest Pest Explorer. Available online: http://www.fs.fed.us/ne/morgantown/4557/AFPE/index.html (accessed on 16 June 2009).
- Eastern Ontario Model Forest. Ecosystem Condition and Productivity. Available online: http://sof.eomf.on.ca/Ecosystem_Condition_and_Productivity/Biotic/Indicators/Disease/Area/i_forest_area_affected_by_disease_e.htm (accessed on 16 June 2009).
- U.S. Forest Service. Forest Insect and Disease Conditions in the United Utates 1999; U.S. Forest Service, Forest Health Protection: Washington, DC, USA, 2007; p. 176.
- Cooperative Emerald Ash Project. Initial County EAB Detections. Available online: http://www.aphis.usda.gov/plant_health/plant_pest_info/emerald_ash_b/downloads/multistateeab.pdf (accessed on 8 June 2013).
- Forest Health Protection. Distribution of Counties with Laurel Wilt Disease by Year of Initial Detection. Available online: http://www.fs.fed.us/r8/foresthealth/laurelwilt/dist_map.shtml (accessed on 8 June 2013).
- De Groot, P.; Nystrom, K.; Scarr, T. Discovery of sirex noctilio (hymenoptera: Siricidae) in Ontario, Canada. Gt. Lakes Entomol. 2006, 39, 49–53. [Google Scholar]
- Weston, P.A.; Desurmont, G.; Hoebeke, E.R. Viburnum leaf beetle. Am. Entomol. 2007, 53, 96–101. [Google Scholar] [CrossRef]
- Plant Health Division. Viburnum Leaf Beetle. Available online: http://www.agri.ohio.gov/divs/plant/caps/vlb.aspx (accessed on 8 June 2013).
- Taylor, R.A.; Bauer, L.S.; Miller, D.L.; Haack, R.A. Emerald ash borer flight potential. In Proceedings of the Emerald Ash Borer Research and Technology Development Meeting, Romulus, MI, USA, 5–6 October 2004; Mastro, V., Reardon, R., Eds.; U.S. Forest Service, Forest Health Technology Enterprise Team: Morgantown, WV, USA; pp. 15–16.
- McClure, M.S. Role of winds, birds, deer, and humans in the dispersal of hemlock woolly adelgid (homoptera: Adelgidae). Environ. Entomol. 1990, 19, 36–43. [Google Scholar] [CrossRef]
- Wainhouse, D. Dispersal of first instar larvae of the felted beech scale, Cryptococcus fagisuga. J. Appl. Ecol. 1980, 17, 523–532. [Google Scholar] [CrossRef]
- Morin, R.S.; Liebhold, A.M.; Tobin, P.C.; Gottschalk, K.W.; Luzader, E. Spread of beech bark disease in the eastern United States and its relationship to regional forest composition. Can. J. For. Res. 2007, 37, 726–736. [Google Scholar] [CrossRef]
- Prasad, A.; Iverson, L.; Peters, M.; Bossenbroek, J.; Matthews, S.; Davis Sydnor, T.; Schwartz, M. Modeling the invasive emerald ash borer risk of spread using a spatially explicit cellular model. Landsc. Ecol. 2010, 25, 353–369. [Google Scholar] [CrossRef]
- Koch, F.H.; Smith, W.D. Spatio-temporal analysis of Xyleborus glabratus (coleoptera: Circulionidae: Scolytinae) invasion in eastern U.S. Forests. Environ. Entomol. 2009, 37, 442–452. [Google Scholar] [CrossRef]
- Sharov, A.A.; Pijanowski, B.C.; Liebhold, A.M.; Gage, S.H. What affects the rate of gypsy moth (lepidoptera: Lymantriidae) spread: Winter temperature or forest susceptibility? Agric. For. Entomol. 1999, 1, 37–45. [Google Scholar] [CrossRef]
- Morin, R.; Liebhold, A.; Gottschalk, K. Anisotropic spread of hemlock woolly adelgid in the eastern United States. Biol. Invasions 2009, 11, 2341–2350. [Google Scholar] [CrossRef]
- Sharov, A.A.; Liebhold, A.M.; Roberts, E.A. Methods for monitoring the spread of gypsy moth (lepidoptera: Lymantriidae) populations in the appalachian mountains. J. Econ. Entomol. 1997, 90, 1259–1266. [Google Scholar] [CrossRef]
- Tobin, P.; Turcotte, R.; Snider, D. When one is not necessarily a lonely number: Initial colonization dynamics of Adelges tsugae on eastern hemlock, Tsuga canadensis. Biol. Invasions 2013, 15, 1925–1932. [Google Scholar] [CrossRef]
- Mercader, R.J.; Siegert, N.W.; Liebhold, A.M.; McCullough, D.G. Dispersal of the emerald ash borer, Agrilus planipennis, in newly-colonized sites. Agric. For. Entomol. 2009, 11, 421–424. [Google Scholar] [CrossRef]
- Siegert, N.W.; McCullough, D.G.; Williams, D.W.; Fraser, I.; Poland, T.M.; Pierce, S.J. Dispersal of Agrilus planipennis (coleoptera: Buprestidae) from discrete epicenters in two outlier sites. Environ. Entomol. 2010, 39, 253–265. [Google Scholar] [CrossRef] [PubMed]
- Fitzpatrick, M.C.; Preisser, E.L.; Porter, A.; Elkinton, J.; Ellison, A.M. Modeling range dynamics in heterogeneous landscapes: Invasion of the hemlock woolly adelgid in eastern North America. Ecol. Appl. 2011, 22, 472–486. [Google Scholar] [CrossRef]
- Meentemeyer, R.K.; Cunniffe, N.J.; Cook, A.R.; Filipe, J.A.N.; Hunter, R.D.; Rizzo, D.M.; Gilligan, C.A. Epidemiological modeling of invasion in heterogeneous landscapes: Spread of sudden oak death in California (1990–2030). Ecosphere 2011, 2, 1–24. [Google Scholar] [CrossRef]
- Bigsby, K.; Tobin, P.; Sills, E. Anthropogenic drivers of gypsy moth spread. Biol. Invasions 2011, 13, 2077–2090. [Google Scholar] [CrossRef]
- Koch, F.H.; Yemshanov, D.; Magarey, R.D.; Smith, W.D. Dispersal of invasive forest insects via recreational firewood: A quantitative analysis. J. Econ. Entomol. 2012, 105, 438–450. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Species | Date of Introduction | Period of Analysis | Number of Records | References |
|---|---|---|---|---|
| Balsam Woolly Adelgid (BWA) | 1908 | 1934 to 1960 | 30 | [38] |
| Beech Bark Disease (BBD) | 1890 | 1929 to 2008 | 157 | [13,24,41,42] |
| Chestnut Blight (CB) | 1904 | 1910 to 1940 | 40 | [13,22] |
| Dogwood Anthracnose (DA) | 1970s | 1989 to 2006 | 380 | [43] |
| Dutch Elm Disease E (DED E) | 1930 | 1944 to 1962 | 190 | [13] |
| Dutch Elm Disease W (DED W) | 1967 | 1967 to 1990 | 22 | [13] |
| Emerald Ash Borer (EAB) | 2002 | 2003 to 2008 | 308 | [37,44] |
| Gypsy Moth (GM) | 1868 | 1905 to 2007 | 542 | [36,37] |
| Hemlock Woolly Adelgid (HWA) | <1951 | 1991 to 2004 | 201 | [14] |
| Laurel Wilt (LW) | 2002 | 2004 to 2008 | 103 | [45] |
| Pine Shoot Beetle (PSB) | 1992 | 1993 to 2006 | 418 | [37,39,40] |
| Sirex Woodwasp (SWW) | 2005 | 2004 to 2012 | 66 | [37,46] |
| Viburnum leaf beetle (VLB) | 1978 | 1998 to 2012 | 110 | [47,48] |
| White Pine Blister Rust E (WPBR E) | 1889 | 1942 to 2008 | 317 | [13] |
| White Pine Blister Rust W (WPBR W) | 1910 | 1923 to 2006 | 124 | [13] |
© 2016 by the author; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Evans, A.M. The Speed of Invasion: Rates of Spread for Thirteen Exotic Forest Insects and Diseases. Forests 2016, 7, 99. https://doi.org/10.3390/f7050099
Evans AM. The Speed of Invasion: Rates of Spread for Thirteen Exotic Forest Insects and Diseases. Forests. 2016; 7(5):99. https://doi.org/10.3390/f7050099
Chicago/Turabian StyleEvans, Alexander M. 2016. "The Speed of Invasion: Rates of Spread for Thirteen Exotic Forest Insects and Diseases" Forests 7, no. 5: 99. https://doi.org/10.3390/f7050099