
Predicting Aboveground Biomass in Second Growth Coast Redwood: Comparing Localized with Generic Allometric Models


Abstract
:1. Introduction
2. Materials and Methods
2.1. Description of Coast Redwood
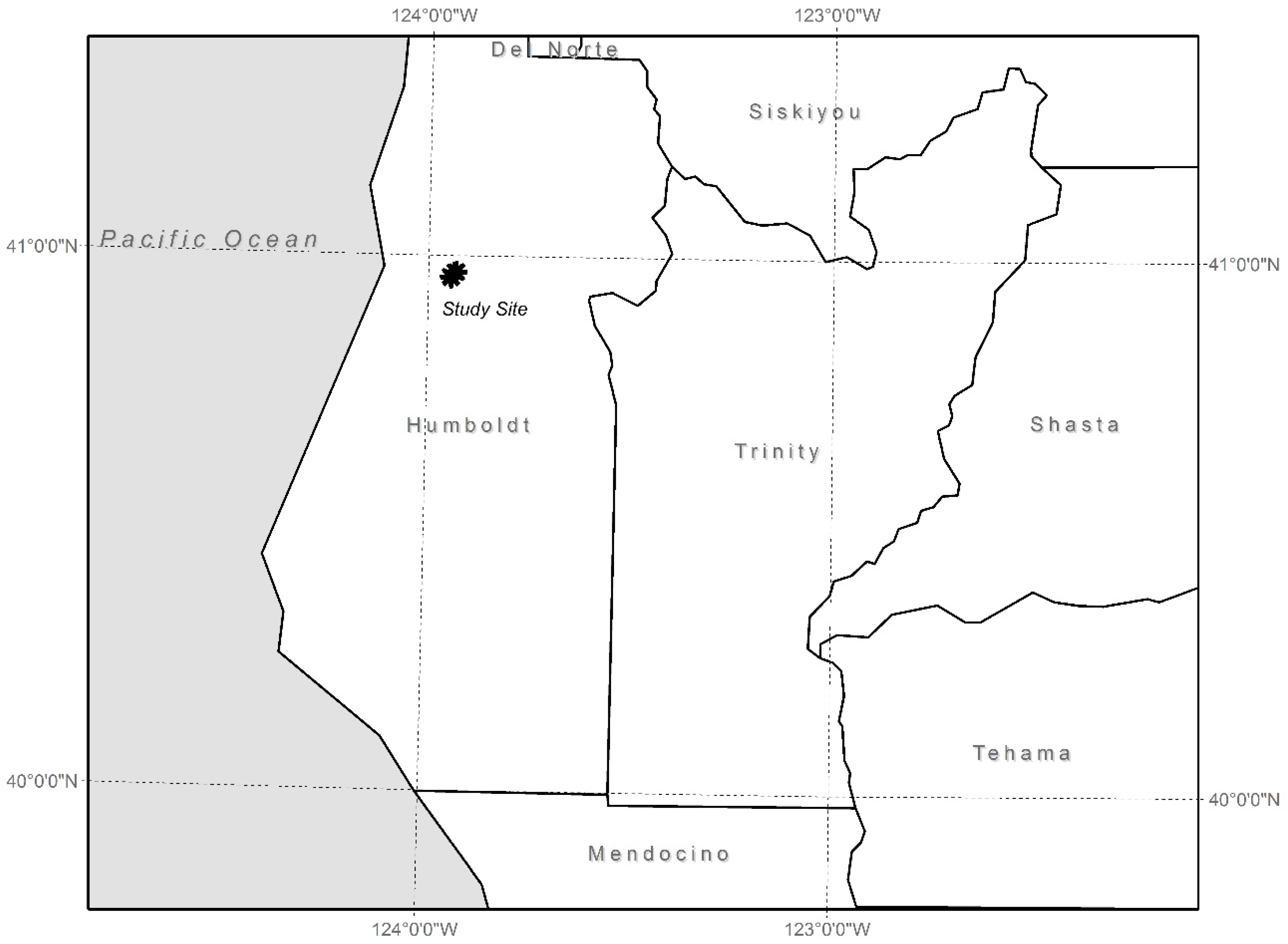
2.2. Study Sites
2.3. Sampling Procedures and Data Collection
2.4. Bole Measurements
2.5. Branch Measurements
2.6. Laboratory Measurements
2.7. Statistical Analysis
3. Results and Discussion
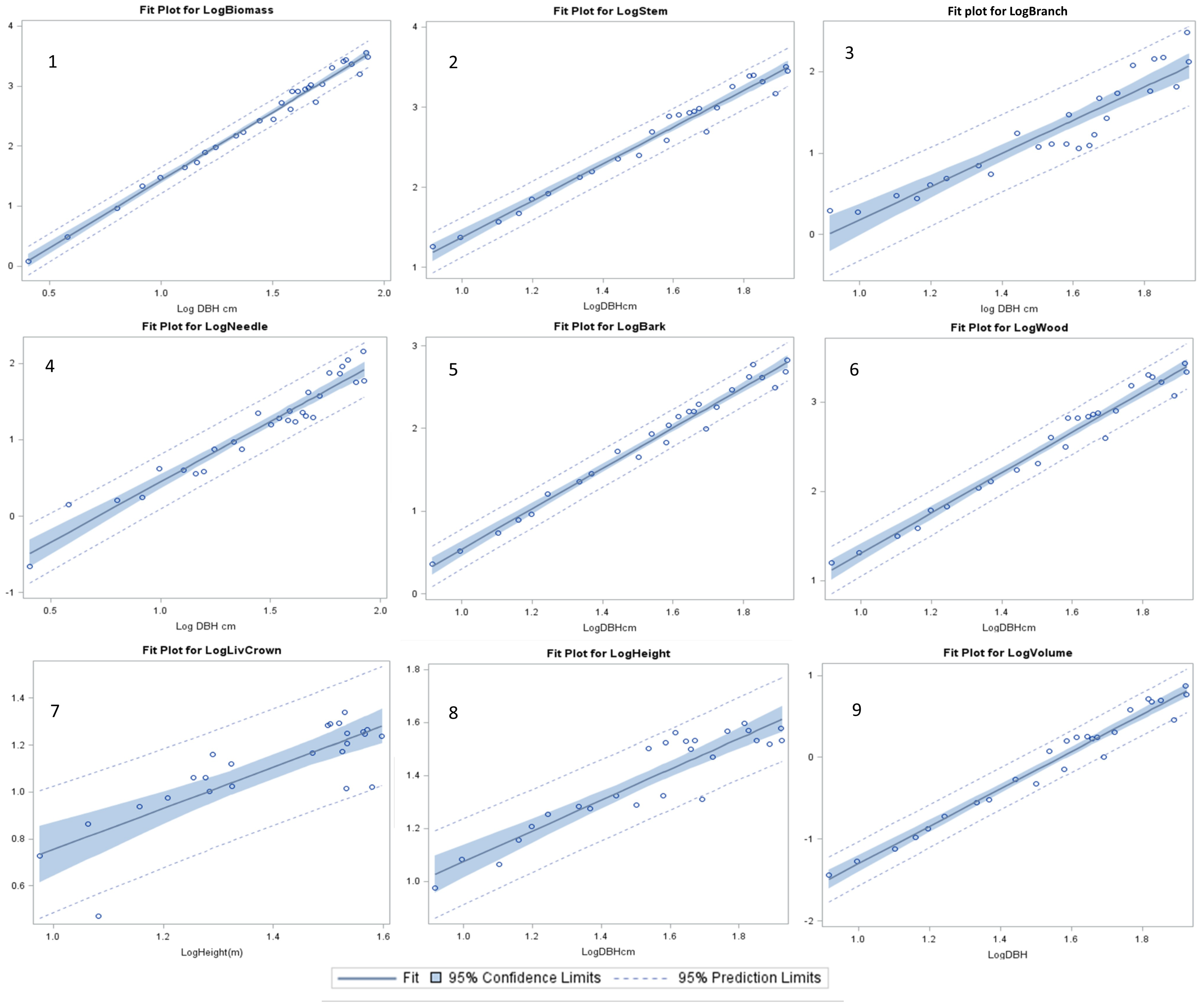
3.1. Model Selection
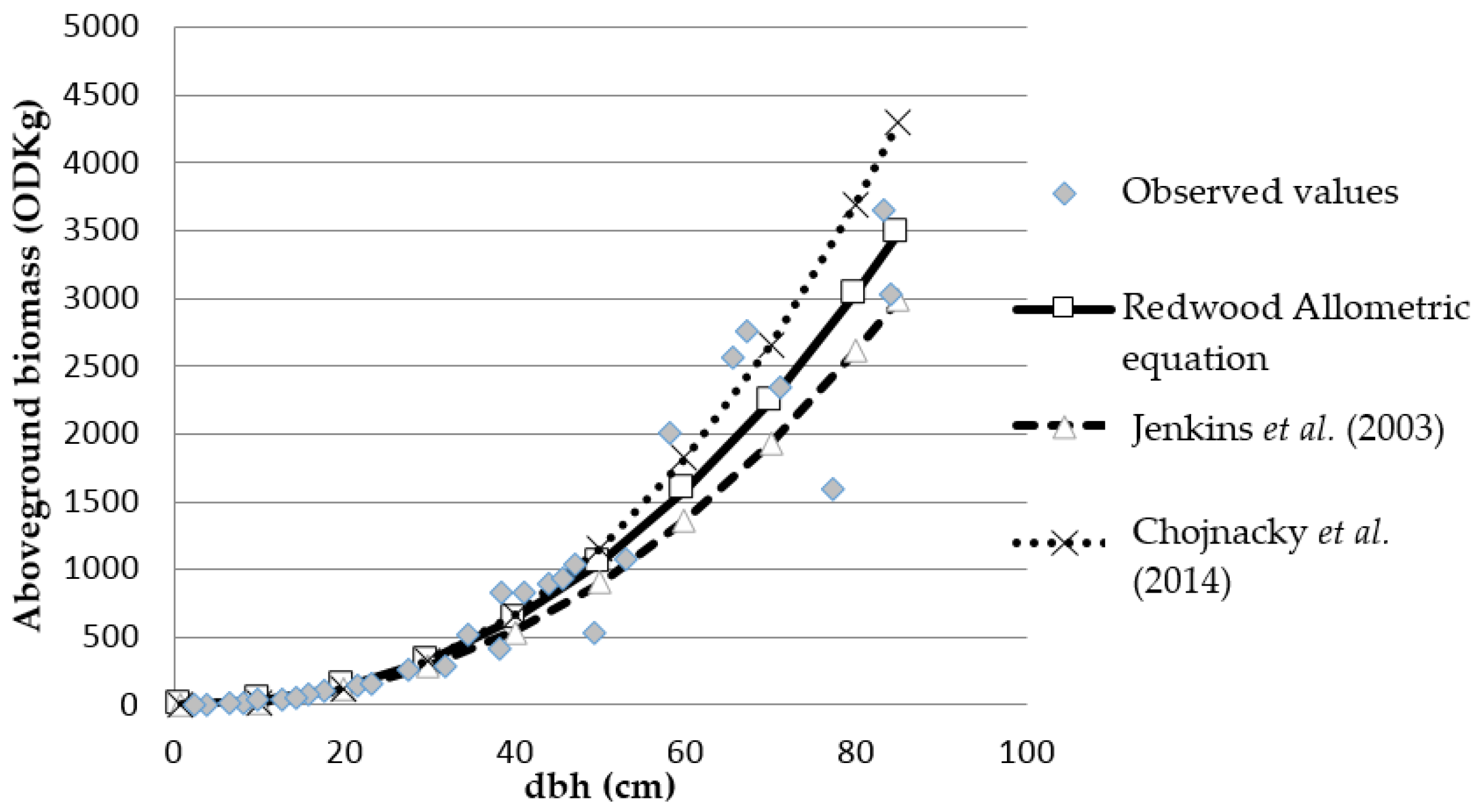
3.2. Comparing with Previous Studies
3.3. Scope and Limitations of Application
4. Conclusions
- Biomass components and tree dimensions, in general, were closely related to the dbh and were able to account for around 90% of the variance.
- The study site can be characterized as a second-growth even-aged stand and the dbh range for candidate trees used to estimate TAGB and foliage was 2.54–84.07 cm, while for all other tree components and dimensions the range was 8.25–84.07 cm.
Acknowledgments
Author Contributions
Conflicts of Interest
Abbreviations
| dbh | diameter at breast height |
| H | Height |
| Ln | natural logarithm |
| log10 | Log to the base of 10 |
| ODKg | Oven dry kilogram |
| TAGB | Total Aboveground Biomass |
References
- Picard, N.; Saint-André, L.; Henry, M. Manual for building tree volume and biomass allometric equations from field measurement to prediction. In Food and Agriculture Organization of the United Nations and Centrede Coopération; Internationale en Recherche Agronomique pour le Développement (CIRAD): Rome, Italy, 2012; p. 215. [Google Scholar]
- Niklas, K.J. Plant Allometry: The Scaling of Form and Process; University of Chicago Press: Chicago, IL, USA, 1994. [Google Scholar]
- Wang, C. Biomass allometric equations for 10 co-occurring tree species in Chinese temperate forests. For. Ecol. Manag. 2006, 222, 19–16. [Google Scholar] [CrossRef]
- Basuki, T.M.; van Laake, P.E.; Skidmore, A.K.; Hussin, Y.A. Allometric equations for estimating the above-ground biomass in tropical lowland Dipterocarp forests. For. Ecol. Manag. 2009, 257, 1684–1694. [Google Scholar] [CrossRef]
- Jenkins, J.C.; Chojnacky, D.C.; Heath, L.S.; Birdsey, R.A. National-scale biomass estimators for United States tree species. For. Sci. 2003, 49, 12–35. [Google Scholar]
- Chojnacky, D.C.; Jenkins, J.C.; Heath, L.S. Updated generalized biomass equations for North American tree species. Forestry 2014, 87, 129–151. [Google Scholar] [CrossRef]
- Koukoulomatis, K.; Mitsopoulos, J.D. Crown fuel weight estimation of Black pine (Pinus nigra) plantations in southern Bulgaria. Silva Balc. 2007, 8, 57–65. [Google Scholar]
- Cobb, R.C.; Chan, M.N.; Meentemeyer, R.K.; Rizzo, D.M. Common factors drive disease and coarse woody debris dynamics in forests impacted by Sudden Oak death. Ecosystems 2012, 15, 242–255. [Google Scholar] [CrossRef]
- Huang, S.; Crabtree, R.L.; Potter, C.; Gross, P. Estimating the quantity and quality of coarse woody debris in Yellowstone post-fire forest ecosystem from fusion of SAR and optical data. Remote Sens. Environ. 2009, 113, 1926–1938. [Google Scholar] [CrossRef]
- Vitorelo, B.D.; Han, H.-S.; Elliot, W. Cost and productivity of two mechanical fire hazard reduction methods: Mastication and thinning. For. Prod. J. 2011, 61, 664–674. [Google Scholar]
- Kizha, A.R.; Han, H.-S. Forest residues recovered from whole-tree timber harvesting operations. Eur. J. For. Eng. 2015, 1, 46–55. [Google Scholar]
- Jenkins, J.C.; Chojnacky, D.C.; Heath, L.S.; Birdsey, R.A. Comprehensive Database of Diameter-Based Biomass Regressions for North American Tree Species; USDA Forest Service General Technical Report NE-319; Northern Research Station: Burlington, VT, USA, 2004; p. 48.
- Woodall, C.W.; Heath, L.S.; Domke, G.M.; Nichols, M.C. Methods and Equations for Estimating Aboveground Volume, Biomass, and Carbon for Trees in the U.S. Forest Inventory 2010; USDA Forest Service General Technical Report NRS-88; Northern Research Station: Newtown Square, PA, USA, 2011; p. 34.
- Woudenberg, S.W.; Conkling, B.L.; O’Connell, B.M.; LaPoint, E.B.; Turner, J.A.; Waddell, K.L. The Forest Inventory and Analysis Database: Database Description and Users Manual Version 4.0 for Phase 2; USDA Forest Service General Technical Report RMRS-GTR-245; Rocky Mountain Research Station: Fort Collins, CO, USA, 2010; p. 344. [Google Scholar]
- Stewart, W. The New Economies of the Redwood Region in the 21st Century; USDA Forest Service General Technical Report PSW-GTR-194; Pacific Southwest Research Station: Albany, CA, USA, 2007; pp. 393–401. [Google Scholar]
- Standiford, R.B. Trends in harvest levels and stumpage prices in coastal California. In Forest Research and Outreach; Cooperative Extension Forestry; University of California: Berkeley, CA, USA, 2012. [Google Scholar]
- Fujimori, T. Ecological and Silvicultural Strategies for Sustainable Forest Management; Elsevier Science: Amsterdam, Netherland, 2001. [Google Scholar]
- Busing, R.T.; Fujimori, T. Biomass, production and woody detritus in an old coast redwood (Sequoia sempervirens) forest. Plant Ecol. 2005, 177, 177–188. [Google Scholar] [CrossRef]
- Fujimori, T. Stem biomass and structure of a mature Sequoia sempervirens stand on the Pacific Coast of northern California. J. Jpn. For. Soc. 1977, 59, 435–441. [Google Scholar]
- Lindquist, J.L.; Palley, M.N. Empiricial Yield Tables for Young-Growth Redwood; California Agricultural Experiment Station; University of California: Berkeley, CA, USA, 1963. [Google Scholar]
- Wensel, L.C.; Krumland, B. Volume and Taper Relationships for Redwood, Douglas Fir Other Conifers in California’s North Coast; Division of Agricultural Sciences; University of California: Berkeley, CA, USA, 1983. [Google Scholar]
- Wensel, L.C.; Olson, C.M. Tree taper models for major commercial California conifers. Hilgardia 1995, 62, 1–18. [Google Scholar]
- Wensel, L.C.; Krumland, B.E.; Meerschaert, W.J. CRYPTOS User's Guide: Cooperative Redwood Yield Project Timber Output Simulator; Bulletin 1924, Agricultural Experiment Station; University of California: Berkeley, CA, USA, 1987; p. 89. [Google Scholar]
- Krumland, B.E.; Wensel, L.C. Preliminary Young Growth Volume Tables for Coastal California Counties; Res. Note No. 1; Cooperative Redwood Yield Research Project; University of California: Berkeley, CA, USA, 1975; p. 18. [Google Scholar]
- Forest Inventory and Analysis (FIA). National Core Field Guide: Field Data Collection Procedures for Phase 2 Plots Version 6.0. Available online: http://www.nrs.fs.fed.us/fia/data-collection/field-guides/p2/NRS_FG_6.0-Apri_2014-Complete_Document_NRSP2plus.pdf (accessed on 22 April 2016).
- California Department of Park and Recreation (CaDPR). About Coast Redwoods. Available online: http://www.parks.ca.gov/?page_id=24200 (accessed on 23 October 2014).
- Griffith, R.S. Sequoia sempervirens. In Fire Effects Information System; USDA Forest Service; Available online: http://www.fs.fed.us/database/feis/ (accessed on 1 September 1992).
- Busing, R.T.; Fujimori, T. NPP Temperate Forest: Humboldt Redwoods State Park, California, U.S.A., 1972–2001. Available online: http://daac.ornl.gov/cgi-bin/dsviewer.pl?ds_id=803 (accessed on 22 April 2016).
- Munz, P.A. A Flora of Southern California; University of California Press: Berkeley, CA, USA, 1974. [Google Scholar]
- Olson, D.F.; Roy, D.F.; Walters, G.A. Sequoia Sempiervirens (D. Don) Endl. In Silvics of North America. Conifers; Burns, R.M., Honkala, B.H., Eds.; USDA Forest Service: Washington, DC, USA, 1990; pp. 541–552. [Google Scholar]
- Preston, R.J. North American Trees; the Iowa State College Press: Ames, IA, USA, 1948. [Google Scholar]
- Western Regional Climate Center (WRCC). Willow Creek 1 NW, California-Climate Summary. Available online: http://www.wrcc.dri.edu/cgi-bin/cliMAIN.pl?cawilc+nca (accessed on 29 May 2015).
- WWF Global. Pacific Temperate Rainforests. Available online: http://wwf.panda.org/about_our_earth/ecoregions/pacific_temperate_rainforests.cfm (accessed on 20 October 2014).
- Stuart, J.D. Fire history of an old-growth forest of Sequoia sempervirens (Taxodiaceae) forest in Humboldt Redwoods State Park, California. Madrono 1987, 4, 128–141. [Google Scholar]
- Stone, E.C.; Vasey, R.B. Preservation of coast redwoods on alluvial flats. Science 1968, 15, 157–161. [Google Scholar] [CrossRef] [PubMed]
- Veirs, S.D. Coast redwood forest: Stand dynamics, successional status, and the role of fire. In Forest Succession and Stand Development Research in the Pacific Northwest; Means, J.E., Ed.; Oregon State University Forest Research Lab: Corvallis, OR, USA, 1982; pp. 119–141. [Google Scholar]
- Monserud, R.A.; Marshall, J.D. Allometric crown relations in three northern Idaho conifer species. Can. J. For. Res. 1999, 29, 521–535. [Google Scholar] [CrossRef]
- Snell, J.A.K.; Little, S.N. Predicting Crown Weight and Bole Volume of Five Western Hardwoods; USDA Forest Service General Technical Report PNW-151; Pacific Northwest forest and range experiment station: Portland, OR, USA, 1983; p. 37. [Google Scholar]
- European Committee for Standardization. Solid Biofuels-Determination of Moisture Content-Oven Dry Method-Part 1: Total Moisture-Reference Method; FprEN 14774-1; Management Centre: Brussels, Belgium, 2009; p. 8. [Google Scholar]
- Parresol, B.R. Assessing tree and stand biomass: A review with examples and critical comparisons. For. Sci. 1999, 45, 573–593. [Google Scholar]
- Sokal, R.R.; Rohlf, F.J. Biometry: The Principles and Practice of Statistics in Biological Research; W.H. Freeman and Co.: New York, NY, USA, 1995. [Google Scholar]
- Spruge, D.G. Correcting for bais in Log transformed allometric equations. Ecology 1983, 64, 209–210. [Google Scholar] [CrossRef]
- SAS Institute Inc. SAS 9.1.3 Help and Documentation; SAS Institute Inc.: Cary, NC, USA, 2004. [Google Scholar]
- Baskerville, G.L. Use of Logarithmic regression in the estimation of plant biomass. Can. J. For. Res. 1972, 2, 49–53. [Google Scholar] [CrossRef]
- Cole, T.G.; Ewel, J.J. Allometric equations for four valuable tropical tree species. For. Ecol. Manag. 2006, 229, 351–360. [Google Scholar] [CrossRef]
- Enquist, B.J. Universal scaling in tree and vascular plant allometry: Toward a general quantitative theory linking plant form and function from cells to ecosystems. Tree Physiol. 2002, 22, 1045–1064. [Google Scholar] [CrossRef] [PubMed]
- Gould, S.J. Allometry and size in ontogeny and phylogeny. Biol. Rev. 1966, 41, 587–640. [Google Scholar] [CrossRef] [PubMed]
- Sileshi, G.W. A critical review of forest biomass estimation models, common mistakes and corrective measures. For. Ecol. Manag. 2014, 329, 237–254. [Google Scholar] [CrossRef]
- Fehrmann, L.; Kleinn, C. General considerations about the use of allometric equations for biomass estimation on the example of Norway spruce in central Europe. For. Ecol. Manag. 2006, 236, 412–421. [Google Scholar] [CrossRef]
- Coons, K.L. Douglas–fir (Psuedotsuga menziesii) Biomass and Nutrient Removal under Varying Harvest Scenarios Involving Co-Production of Timber and Feedstock for Liquid Biofuels; Master of Science; Oregon State University: Corvallis, OR, USA, 19 December 2014. [Google Scholar]
- Fried, J.S.; Zhou, X. Forest Inventory-Based Estimation of Carbon Stocks and Flux in California Forests in 1990; USDA Forest Service General Technical Report PNW-GTR-750; Pacific northwest forest and range experiment station: Portland, OR, USA, 2008; p. 32. [Google Scholar]
- Case, B.S.; Hall, R.J. Assessing prediction errors of generalized tree biomass and volume equation for the boreal forest region of west-central Canada. Can. J. For. Res. 2008, 38, 878–889. [Google Scholar] [CrossRef]
- Berrill, J.-P.; Jeffress, J.L.; Engle, J.M. Coast redwood live crown and sapwood dynamics. In Proceedings of Coast Redwood Forests in a Changing California: A Symposium for Scientists and Managers; Standiford, R.B., Weller, T.J., Piirto, D.D., Stuart, J.D., Eds.; USDA Forest Service General Technical Report PSW-GTR-238; Pacific Southwest Research Station: Albany, CA, USA, 2012; pp. 473–484. [Google Scholar]
- Segura, M.; Kanninen, M. Allometric models for tree volume and total aboveground biomass in a tropical humid forest in Costa Rica. Biotropica 2005, 37, 2–8. [Google Scholar] [CrossRef]
- Litton, C.M.; Kauffman, J.B. Allometric models for predicting aboveground biomass in two widespread woody plants in Hawaii. Biotropica 2008, 4, 313–320. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Tree/Biomass Components | Description |
|---|---|
| dbh | Diameter at breast height; measured in centimeters (cm). |
| Height (H) | Total height of the tree from the ground to tip of the crown; measured in meters (m). |
| Stump | Remaining portion (0.30 m above ground) of the stem after the tree is felled with roots still in the ground. For calculating TAGB, the biomass in stump was included in the bole. |
| Bole (stem) | Biomass in stump, bole and bark up to 2.54 cm top; measured in oven dry kilograms (ODkg). |
| Live branch | Biomass in living branches; measured in ODkg. |
| Dead branch | Biomass in dead branches; measured in ODkg. |
| Foliage | Biomass in entire foliage (both live and dead); measured in ODkg. |
| Wood weight | Biomass in bole (main stem) above the stump including bark; measured in ODkg. |
| Bark | Biomass in bark; measured in ODkg. |
| Total aboveground biomass (TAGB) | Biomass in all tree components including bole, branch (dead and live), and foliage; measured in ODkg. |
| Height to live crown base | Height from an imaginary horizontal line drawn across the stem from the bottom of the lowest live foliage of the live crown for trees and from the lowest live foliage of the lowest twig for saplings [25]; measured in m. |
| Volume | Volume of bole (including bark) above the stump up to the top 2.54 cm; measured in m3. |
| Stands | Tree Types | A | B |
|---|---|---|---|
| Ground slope range (%) | 3–37 | 0–50 | |
| Average slope (%) | 22 | 31 | |
| Basal area (m2·ha−1) | C | 9 | 9 |
| H | 4 | 2 | |
| D | 0.4 | 1 | |
| SD | 0.5 | 2 | |
| Trees density (Number of trees ha−1) | C | 75 | 84 |
| H | 45 | 42 | |
| D | 17 | 23 | |
| SD | 126 | 194 | |
| Average dbh (cm) | C | 35 | 37 |
| H | 31 | 25 | |
| D | 14 | 14 | |
| SD | 9 | 10 |
| Diameter at Breast Height (cm) | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| 1–10 | 11–20 | 21–30 | 31–40 | 41–50 | 51–60 | 61–70 | 71–80 | 81–90 | Total | ||
| Height (m) | 1–5 | 3 | 3 | ||||||||
| 5–10 | 1 | 1 | |||||||||
| 11–15 | 1 | 2 | 3 | ||||||||
| 16–20 | 2 | 2 | 1 | 1 | 6 | ||||||
| 21–25 | 1 | 1 | 2 | ||||||||
| 26–30 | 1 | 1 | |||||||||
| 31–35 | 2 | 3 | 2 | 1 | 8 | ||||||
| 36–40 | 1 | 1 | 2 | 1 | 5 | ||||||
| Total | 5 | 4 | 3 | 4 | 5 | 2 | 2 | 2 | 2 | 29 | |
| Intercept | dbh a | H b | R2 | Adjusted R2 | RMSE c | CF d | |
|---|---|---|---|---|---|---|---|
| TAGB | −0.8252 | 2.2607 | 0.9878 | 0.9874 | 0.1036 | 0.0054 | |
| Biomass Components | |||||||
| Bole (stem) e | −0.9180 | 2.2931 | 0.9740 | 0.9729 | 0.1118 | 0.0062 | |
| Live branch | −1.8562 | 2.0382 | 0.8804 | 0.8754 | 0.2242 | 0.0251 | |
| Dead branch | −3.5952 | 2.3257 | 0.5766 | 0.5589 | 0.5948 | 0.1769 | |
| Foliage | −1.3094 | 1.5819 | 0.9386 | 0.9364 | 0.1669 | 0.0139 | |
| Bark | −1.9063 | 2.4469 | 0.9787 | 0.9779 | 0.1076 | 0.0058 | |
| Tree dimensions | |||||||
| Wood f | −0.9553 | 2.2613 | 0.9710 | 0.9698 | 0.1167 | 0.0068 | |
| Height | 0.4943 | 0.5808 | 0.8511 | 0.8449 | 0.0725 | 0.0026 | |
| Volume of bole g | −3.5879 | 2.2859 | 0.9690 | 0.9677 | 0.1220 | 0.0074 | |
| Height to live crown | −0.1241 | 0.8791 | 0.6715 | 0.6572 | 0.1176 | 0.0069 | |
| Height to live crown | 0.3275 | 0.5034 | 0.5529 | 0.5335 | 0.1372 | 0.0094 | |
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kizha, A.R.; Han, H.-S. Predicting Aboveground Biomass in Second Growth Coast Redwood: Comparing Localized with Generic Allometric Models. Forests 2016, 7, 96. https://doi.org/10.3390/f7050096
Kizha AR, Han H-S. Predicting Aboveground Biomass in Second Growth Coast Redwood: Comparing Localized with Generic Allometric Models. Forests. 2016; 7(5):96. https://doi.org/10.3390/f7050096
Chicago/Turabian StyleKizha, Anil Raj, and Han-Sup Han. 2016. "Predicting Aboveground Biomass in Second Growth Coast Redwood: Comparing Localized with Generic Allometric Models" Forests 7, no. 5: 96. https://doi.org/10.3390/f7050096