
The Effects of Disturbance History on Ground-Layer Plant Community Composition in British Columbia

Abstract
:
1. Introduction
2. Materials and Methods
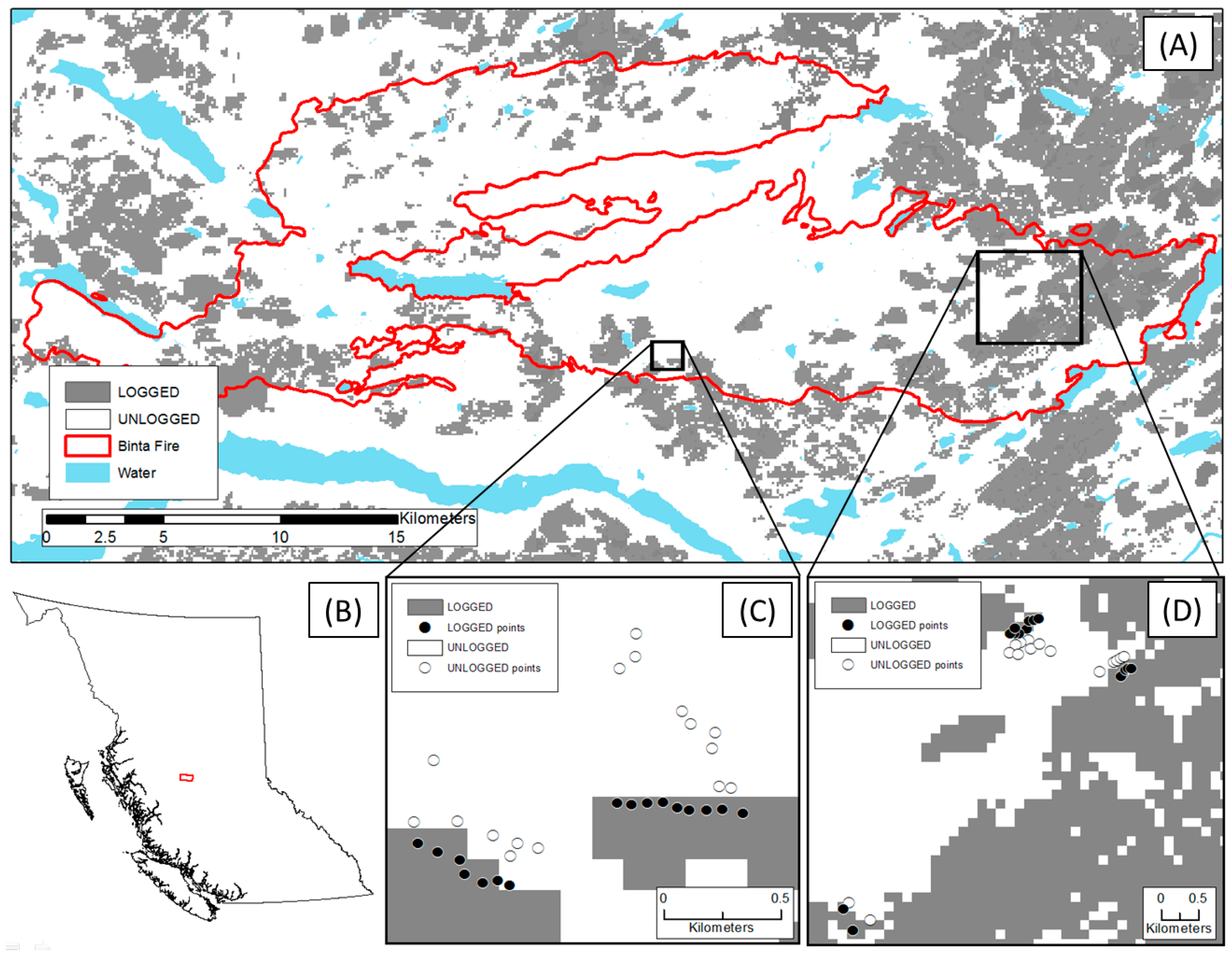
2.1. Study Area
2.2. Data Collection
2.2.1. Site and Sample Point Selection
2.2.2. Plant Community Data
2.2.3. Environmental Variables
2.3. Analysis
2.3.1. Community Richness, Diversity, and Evenness
2.3.2. Differences in Community Composition
2.3.3. Indicator Species Analysis
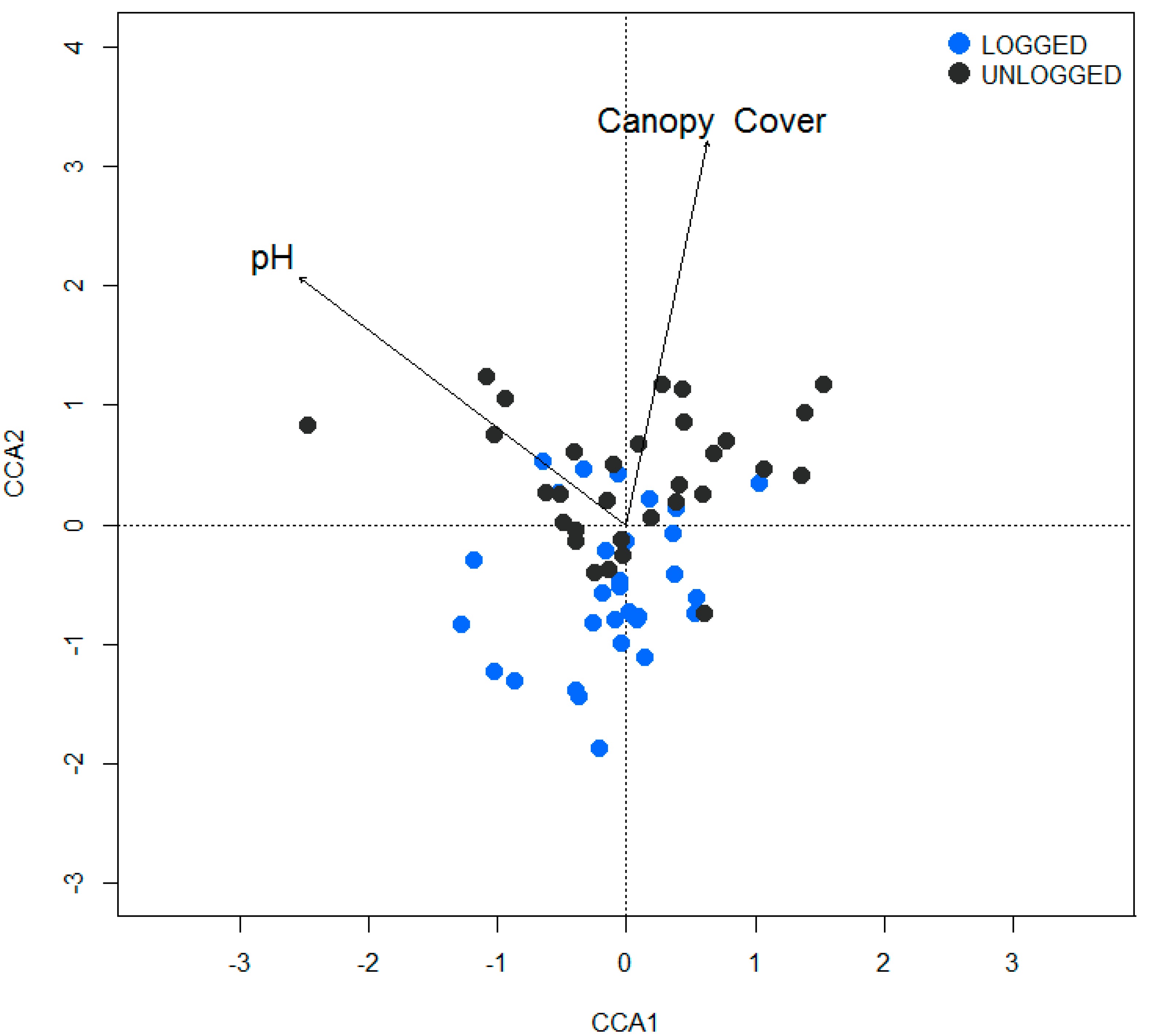
2.3.4. Canonical Correspondence Analysis
2.3.5. Trait Analyses
3. Results
3.1. Community Richness, Diversity, and Evenness
3.2. Community Composition
3.3. Indicator Species Analysis
3.4. Canonical Correspondence Analysis
3.5. Trait Differences
4. Discussion
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
Abbreviations
| ANOSIM | Analysis of similarity |
| ANOVA | Analysis of variance |
| BC | British Columbia |
| CCA | Canonical correspondence analysis |
| dNBR | Difference in normalized burn ratio |
| GIS | Geographic information system |
| MFLNRO | Ministry of Forest, Lands and Natural Resource Operations |
| MRBP | Blocked multi-response permutation procedure |
| MRPP | Multi-response permutation procedure |
Appendix A
| A list of all species included in the paper with authority names. |
| Abies lasiocarpa Hook. Nutt |
| Arnica cordifolia Hook. |
| Aster foliaceus Lindl. ex DC. |
| Ceratodon purpureus Hedw. Brid. |
| Chamerion angustifolium var. angustifolium (L.) Holub |
| Cornus canadensis L. |
| Dendroctonus ponderosae Hopkins |
| Hieracium gracile Hook. |
| Petasites frigidus var. vitifolius Greene Cherne |
| Picea glauca Moench Voss × engelmannii Parry ex Engelm. |
| Pinus contorta var. latifolia Engelm. ex S. Watson |
| Polytrichum juniperinum Hedw. |
| Populus tremuloides Michx. |
| Rosa acicularis Lindl. |
| Rubus idaeus L. |
| Spiraea betulifolia Pall. |
| Taraxacum officinale F.H. Wigg. |
| Vaccinium membranaceum Douglas ex Torr. |
References
- Clark, D.F.; Antos, J.A.; Bradfield, G.E. Succession in sub-boreal forests of West-Central British Columbia. J. Veg. Sci. 2003, 14, 721–732. [Google Scholar] [CrossRef]
- De Grandpre, L.; Gagnon, D.; Bergeron, Y. Changes in the Understory of Canadian Southern Boreal Forest after Fire. J. Veg. Sci. 1993, 4, 803–810. [Google Scholar] [CrossRef]
- Hart, S.; Chen, H. Fire, logging, and overstory affect understory abundance, diversity, and composition in boreal forest. Ecol. Monogr. 2008, 78, 123–140. [Google Scholar] [CrossRef]
- Selmants, P.C.; Knight, D.H. Understory plant species composition 30–50 years after clearcutting in southeastern Wyoming coniferous forests. For. Ecol. Manag. 2003, 185, 275–289. [Google Scholar] [CrossRef]
- Kurulok, S.E.; Macdonald, S.E. Impacts of postfire salvage logging on understory plant communities of the boreal mixedwood forest 2 and 34 years after disturbance. Can. J. For. Res. 2007, 37, 2637–2651. [Google Scholar] [CrossRef]
- Key, C.H.; Benson, N.C. Landscape Assessment (LA) Sampling and Analysis Methods; USDA Forest Service: Odgen, UT, USA, 2006.
- Cuddington, K. Legacy Effects: The Persistent Impact of Ecological Interactions. Biol. Theory 2012, 6, 203–210. [Google Scholar] [CrossRef]
- Macadam, A.M. Effects of broadcast slash burning on fuels and soil chemical properties in the Sub-boreal Spruce Zone of central British Columbia. Can. J. For. Res. 1987, 17, 1577–1584. [Google Scholar] [CrossRef]
- Arocena, J.M.; Opio, C. Prescribed fire-induced changes in properties of sub-boreal forest soils. Geoderma 2003, 113, 1–16. [Google Scholar] [CrossRef]
- Knicker, H. How does fire affect the nature and stability of soil organic nitrogen and carbon? A review. Biogeochemistry 2007, 85, 91–118. [Google Scholar] [CrossRef]
- Huffman, E.L.; MacDonald, L.H.; Stednick, J.D. Strength and persistence of fire-induced soil hydrophobicity under ponderosa and lodgepole pine, Colorado Front Range. Hydrol. Processes 2001, 15, 2877–2892. [Google Scholar] [CrossRef]
- Reich, P.B.; Bakken, P.; Carlson, D.; Frelich, L.E.; Friedman, S.K.; Grigal, D.F. Influence of Logging, Fire, and Forest Type on Biodiversity and Productivity in Southern Boreal Forests. Ecology 2001, 82, 2731–2748. [Google Scholar] [CrossRef]
- Rees, D.C.; Juday, G.P. Plant species diversity on logged versus burned sites in central Alaska. For. Ecol. Manag. 2002, 155, 291–302. [Google Scholar] [CrossRef]
- Mannerkoski, H.; Finér, L.; Piirainen, S.; Starr, M. Effect of clear-cutting and site preparation on the level and quality of groundwater in some headwater catchments in eastern Finland. For. Ecol. Manag. 2005, 220, 107–117. [Google Scholar] [CrossRef]
- Piirainen, S.; Finér, L.; Mannerkoski, H.; Starr, M. Carbon, nitrogen and phosphorus leaching after site preparation at a boreal forest clear-cut area. For. Ecol. Manag. 2007, 243, 10–18. [Google Scholar] [CrossRef]
- Paine, R.; Tegner, M.; Johnson, E. Compounded perturbations yield ecological surprises. Ecosystems 1998, 1, 535–545. [Google Scholar] [CrossRef]
- Buma, B.; Wessman, C.A. Disturbance interactions can impact resilience mechanisms of forests. Ecosphere 2011, 2, 1–13. [Google Scholar] [CrossRef]
- Harvey, B.J.; Donato, D.C.; Romme, W.H.; Turner, M.G. Influence of recent bark beetle outbreak on fire severity and postfire tree regeneration in montane Douglas-fir forests. Ecology 2013, 94, 2475–2486. [Google Scholar] [CrossRef] [PubMed]
- Pidgen, K.; Mallik, A.U. Ecology of Compounding Disturbances: The Effects of Prescribed Burning After Clearcutting. Ecosystems 2013, 16, 170–181. [Google Scholar] [CrossRef]
- Diaz, S.; Cabido, M. Vive la difference: Plant functional diversity matters to ecosystem processes. Trends Ecol. Evol. 2001, 16, 646–655. [Google Scholar] [CrossRef]
- Diaz, S.; Cabido, M.; Casanoves, F. Plant functional traits and environmental filters at a regional scale. J. Veg. Sci. 1998, 9, 113–122. [Google Scholar] [CrossRef]
- Keddy, P.A. Assembly and response rules: Two goals for predictive community ecology. J. Veg. Sci. 1992, 3, 157–164. [Google Scholar] [CrossRef]
- Decocq, G.; Aubert, M.; Dupont, F.; Alard, D.; Saguez, R.; Wattez-Franger, A.; De Foucault, B.; Delelis-Dusollier, A.; Bardat, J. Plant diversity in a managed temperate deciduous forest: Understorey response to two silvicultural systems. J. Appl. Ecol. 2004, 41, 1065–1079. [Google Scholar] [CrossRef]
- Lachance, É.; Pothier, D.; Bouchard, M. Forest Structure and Understory Plant Communities Inside and Outside Tree Retention Groups in Boreal Forests. Ecoscience 2013, 20, 252–263. [Google Scholar] [CrossRef]
- Donato, D.C.; Fontaine, J.B.; Robinson, W.D.; Kauffman, J.B.; Law, B.E. Vegetation response to a short interval between high-severity wildfires in a mixed-evergreen forest. J. Ecol. 2009, 97, 142–154. [Google Scholar] [CrossRef]
- Delitti, W.; Ferran, A.; Trabaud, L.; Vallejo, V.R. Effects of fire recurrence in Quercus coccifera L. shrublands of the Valencia Region (Spain): I. plant composition and productivity. Plant Ecol. 2005, 177, 57–70. [Google Scholar] [CrossRef]
- Pojar, J.; Klinka, K.; Meidinger, D.V. Biogeoclimatic ecosystem classification in British Columbia. For. Ecol. Manag. 1987, 22, 119–154. [Google Scholar] [CrossRef]
- Picketts, I.M.; Werner, A.T.; Murdock, T.Q. Climate Change in Prince George: Summary of Past Trends and Future Projections; University of Northern British Columbia: Prince George, BC, Canada, 2009. [Google Scholar]
- Reynolds, G. Climatic Data Summaries for the Biogeoclimatic Zones of British Columbia; Ministry of Forests Research Branch: Victoria, BC, Canada, 1989; (unpublished report).
- BC Ministry of Forests, Lands and Natural Resource Operations. Summary of Previous Fire Seasons: 2010 Fire Season. Available online: http://bcwildfire.ca/History/SummaryArchive.htm#2010 (accessed on 1 April 2014).
- DeLong, C.; Tanner, D.; Jull, M.J. A Field Guide for Site Identification and Interpretation for the Northern Rockies Portion of the Prince George Forest Region: Land Management Handbook; No. 29; Ministry of Forests: Victoria, BC, Canada, 1993.
- BC Ministry of Forests, Lands and Natural Resource Operations. Mid-Term Timber Supply: Prince George Timber Supply Area; Ministry of Forests, Lands and Natural Resource Operations: Prince George, BC, Canada, 2012.
- Edwards, M.; Krawchuk, M.A.; Burton, P.J. Short-interval disturbance in lodgepole pine forests, British Columbia, Canada: Understory and overstory response to mountain pine beetle and fire. For. Ecol. Manag. 2015, 338, 163–175. [Google Scholar] [CrossRef]
- BC Ministry of Forests, Lands and Natural Resource Operations. Vegetation Resource Inventory; Ministry of Forests, Lands and Natural Resource Operations: Victoria, BC, Canada, 2013.
- Walton, A. Provincial-Level Projection of the Current Mountain Pine Beetle Outbreak: Update of the Infestation Projection Based on the Provincial Aerial Overview Surveys of Forest Health Conducted from 1999 through 2012 and the BCMPB Model (Year 10); BC Forest Service: Powell River, BC, Canada, 2013. [Google Scholar]
- Harris, C.; (BC Ministry of Forests, Lands and Natural Resource Operations, Victoria, BC, Canada). Personal communication, 2015.
- Cornelissen, J.H.C.; Lavorel, S.; Garnier, E.; Díaz, S.; Buchmann, N.; Gurvich, D.E.; Reich, P.B.; ter Steege, H.; Morgan, H.D.; Heijden, M.G.A.; et al. A handbook of protocols for standardised and easy measurement of plant functional traits worldwide. Aust. J. Bot. 2003, 51, 335–380. [Google Scholar] [CrossRef]
- Schneider, C.A.; Rasband, W.S.; Eliceiri, K.W. NIH Image to ImageJ: 25 years of image analysis. Nat. Methods 2012, 9, 671–675. [Google Scholar] [CrossRef] [PubMed]
- Murray, D. Measuring pH in Water or CaCl2 Using a pH Meter; SFU Geography: Burnaby, BC, Canada, 2011. [Google Scholar]
- Vegan: Community Ecology Package, version 2.2-1; 2015; Available online: https://cran.r-project.org/web/packages/vegan/ (accessed on 12 January 2015).
- R: A Language and Environment for Statistical Computing, version 3.1.1; 2014; Available online: https://cran.r-project.org (accessed on 10 July 2014).
- Benjamini, Y.; Hochberg, Y. Controlling the False Discovery Rate—A Practical and Powerful Approach. J. R. Stat. Soc. Ser. B-Methodol. 1995, 57, 289–300. [Google Scholar]
- PC-ORD: Multivariate Analysis of Ecological Data, version 5; MjM Software Design: Glaneden Beach, OR, USA, 2011.
- Dufrene, M.; Legendre, P. Species assemblages and indicator species: The need for a flexible asymmetrical approach. Ecol. Monogr. 1997, 67, 345–366. [Google Scholar] [CrossRef]
- De Cáceres, M.; Legendre, P. Associations between species and groups of sites: Indices and statistical inference. Ecology 2009, 90, 3566–3574. [Google Scholar] [CrossRef] [PubMed]
- De Cáceres, M.; Font, X.; Oliva, F. Assessing species diagnostic value in large data sets: A comparison between phi-coefficient and Ochiai index. J. Veg. Sci. 2008, 19, 779–788. [Google Scholar] [CrossRef]
- McCune, B.; Grace, J.B. Analysis of Ecological Communities, 3rd ed.; MjM Software Design: Glaneden Beach, OR, USA, 2002. [Google Scholar]
- Faith, D.P.; Minchin, P.R.; Belbin, L. Compositional dissimilarity as a robust measure of ecological distance. Vegetatio 1987, 69, 57–68. [Google Scholar] [CrossRef]
- Minchin, P.R. An evlauation of the relative robustness of techniques for ecoloigcal ordination. Vegetatio 1987, 69, 89–107. [Google Scholar] [CrossRef]
- Moss, E. The ecology of Epilobium angustifolium with particular reference to rings of periderm in the wood. Am. J. Bot. 1936, 23, 114–120. [Google Scholar] [CrossRef]
- Thomas, P.A.; Proctort, M.C.F.; Maltby, E. The Ecology of Severe Moorland Fire on the North York Moors: Chemical and Physical Constraints on Moss Establishment from Spores. J. Ecol. 1994, 82, 457–474. [Google Scholar] [CrossRef]
- Cherniawsky, D.M.; Bayer, R.J. Systematics of North American Petasites (Asteraceae: Senecioneae). III. A taxonomic revision. Can. J. Bot. 1998, 76, 2061–2075. [Google Scholar] [CrossRef]
- Favorite, J.; Anderson, M.K. Plant Guide: American Red Raspberrry Rubus idaeus L.; United States Department of Agiculture: Washington, DC, USA, 2003.
- Scott, N.A.; Saggar, S.; McIntosh, P.D. Biogeochemical impact of Hieracium invasion in New Zealand’s grazed tussock grasslands: Sustainability implications. Ecol. Appl. 2001, 11, 1311–1322. [Google Scholar] [CrossRef]
- Cavieres, L.A.; Quiroz, C.L.; Molina-Montenegro, M.A.; Muñoz, A.A.; Pauchard, A. Nurse effect of the native cushion plant Azorella monantha on the invasive non-native Taraxacum officinale in the high-Andes of central Chile. Perspect. Plant Ecol. Evol. Syst. 2005, 7, 217–226. [Google Scholar] [CrossRef]
- Wilson, L.M. Key to Identification of Invasive and Native Hawkweeds (Hieracium spp.) in the Pacific Northwest; Ministry of Forests and Range: Kamloops, BC, Canada, 2007.
- Stevens, M.; Anderson, M.K. Plant Guide: Black Huckleberry Vaccinium Membranaceum; United States Department of Agiculture: Washington, DC, USA, 2000.
- Wherry, E.T. Temperature relations of the bunchberry, Cornus canadensis L. Ecology 1934, 15, 440–443. [Google Scholar] [CrossRef]
- Myerscough, P.J. Epilobium angustifolium L. J. Ecol. 1980, 68, 1047. [Google Scholar] [CrossRef]
- Kao, R.H. Origins and widespread distribution of co-existing polyploids in Arnica cordifolia (Asteraceae). Ann. Bot. 2008, 101, 145–152. [Google Scholar] [CrossRef] [PubMed]
- Young, D.R.; Smith, W.K. Influence of sunlight on photosynthesis, water relations, and leaf structure in the understory species Arnica cordifolia. Ecology 1980, 61, 1380–1390. [Google Scholar] [CrossRef]
- Härdtle, W.; von Oheimb, G.; Friedel, A.; Meyer, H.; Westphal, C. Relationship between pH-values and nutrient availability in forest soils—The consequences for the use of ecograms in forest ecology. Flora 2004, 199, 134–142. [Google Scholar] [CrossRef]
- Pearsall, W.H. The pH of natural soils and its ecological significance. J. Soil Sci. 1952, 3, 41–51. [Google Scholar] [CrossRef]
- Marage, D.; Gégout, J.C. Importance of soil nutrients in the distribution of forest communities on a large geographical scale. Glob. Ecol. Biogeogr. 2009, 18, 88–97. [Google Scholar] [CrossRef]
- Flesch, T.K.; Wilson, J.D. Wind and remnant tree sway in forest cutblocks. Agric. For. Meteorol. 1999, 93, 229–242. [Google Scholar] [CrossRef]
- Purdon, M.; Biais, S.; Bergeron, Y. Initial response of understorey vegetation to fire severity and salvage-logging in the southern boreal forest of Québec. Appl. Veg. Sci. 2004, 7, 49–60. [Google Scholar] [CrossRef]
- Childs, S.W.; Flint, L.E. Effect of shadecards, shelterwoods, and clearcuts on temperature and moisture environments. For. Ecol. Manag. 1987, 18, 205–217. [Google Scholar] [CrossRef]
- Wang, G.G.; Kemball, K.J. Effects of fire severity on early development of understory vegetation. Can. J. For. Res. 2005, 35, 254–262. [Google Scholar] [CrossRef]
- Lanta, V.; Lepš, J. How does surrounding vegetation affect the course of succession: A five-year container experiment. J. Veg. Sci. 2009, 20, 686–694. [Google Scholar] [CrossRef]
- Quinby, P.A. First-year impacts of shelterwood logging on understory vegetation in an old-growth pine stand in central Ontario, Canada. Environ. Conserv. 2000, 27, 229–241. [Google Scholar] [CrossRef]
- Zenner, E.K.; Kabrick, J.M.; Jensen, R.G.; Peck, J.E.; Grabner, J.K. Responses of ground flora to a gradient of harvest intensity in the Missouri Ozarks. For. Ecol. Manag. 2006, 222, 326–334. [Google Scholar] [CrossRef]
- Stone, W.E.; Wolfe, M.L. Response of understory vegetation to variable tree mortality following a mountain pine beetle epidemic in lodgepole pine stands in Northern Utah. Vegetatio 1996, 122, 1–12. [Google Scholar] [CrossRef]
- Kovacic, D.A.; Dyer, M.I.; Cringan, A.T. Understory biomass in ponderosa pine following mountain pine beetle infestation. For. Ecol. Manag. 1985, 13, 53–67. [Google Scholar] [CrossRef]
- Law, R.; Morton, D.R. Alternative Permanent States of Ecological Communities. Ecology 1993, 74, 1347–1361. [Google Scholar] [CrossRef]
- Beisner, B.; Haydon, D.; Cuddington, K. Alternative stable states in ecology. Front. Ecol. Environ. 2003, 1, 376–382. [Google Scholar] [CrossRef]
- Fukami, T.; Nakajima, M. Community assembly: Alternative stable states or alternative transient states? Ecol. Lett. 2011, 14, 973–984. [Google Scholar] [CrossRef] [PubMed]
- Parminter, J. Burning Alternatives Panel: A Review of Fire Ecology, Fire History and Prescribed Burning in Southern British Columbia; Fire Ecologist Protection Branch: Victoria, BC, Canada, 1991. [Google Scholar]
- Kranabetter, J.M.; Macadam, A.M. Ten-Year Results from Operational Broadcast Burning Trials in Northwestern British Columbia; Ministry of Forests: Victoria, BC, Canada, 1998.



{kind=link}
{kind=link}
{kind=link}
| LOGGED Treatment | UNLOGGED Treatment | ||||
|---|---|---|---|---|---|
| Species | Frequency Observed | Abundance (%) | Species | Frequency Observed | Abundance (%) |
| Chamerion angustifolium | 30 | 22.0 | Chamerion angustifolium | 31 | 29.2 |
| Ceratodon purpureus | 29 | 17.6 | Ceratodon purpureus | 29 | 18.4 |
| Cornus canadensis | 23 | 0.5 | Polytrichum juniperinum | 28 | 5.4 |
| Spiraea betulifolia | 22 | 3.8 | Cornus canadensis | 25 | 1.7 |
| Polytrichum juniperinum | 20 | 3.5 | Arnica cordifolia | 24 | 3.0 |
| Petasites frigidus | 16 | 2.0 | Vaccinium membranaceum | 23 | 2.3 |
| Rubus idaeus | 16 | 0.5 | Spiraea betulifolia | 22 | 5.1 |
| Vaccinium membranaceum | 16 | 0.1 | Rosa acicularis | 16 | 1.1 |
| Growth Form | Mean Biomass (g) LOGGED | Mean Biomass (g) UNLOGGED | p-Value | p.adjusted |
|---|---|---|---|---|
| Grass | 1.48 | 0.73 | 0.242 | 0.242 |
| Shrub | 1.05 | 3.50 | 0.000 | 0.001 |
| Forb | 9.07 | 12.56 | 0.099 | 0.148 |
| Site | Treatment | Species | A | B | Test Statistic | p-Value | Pearson’s r | p-Value |
|---|---|---|---|---|---|---|---|---|
| EAST | LOGGED | Aster foliaceus | 1.000 | 0.400 | 0.632 | 0.017 | 0.384 | 0.017 |
| Petasites frigidus | 1.000 | 0.333 | 0.577 | 0.046 | 0.204 | 0.039 | ||
| UNLOGGED | Vaccinium membranaceum | - | - | - | - | 0.487 | 0.002 | |
| Chamerion angustifolium | - | - | - | - | 0.454 | 0.011 | ||
| WEST | LOGGED | Petasites frigidus | 0.815 | 0.688 | 0.749 | 0.030 | - | - |
| Taraxacum officinale | 1.000 | 0.375 | 0.612 | 0.017 | 0.338 | 0.019 | ||
| UNLOGGED | Arnica cordifolia | 0.9099 | 0.8750 | 0.892 | 0.010 | 0.435 | 0.008 | |
| Vaccinium membranaceum | 0.9031 | 0.7500 | 0.823 | 0.006 | - | - | ||
| Global | LOGGED | Rubus idaeus | 0.848 | 0.516 | 0.662 | 0.048 | - | - |
| Petasites frigidus | 0.845 | 0.516 | 0.660 | 0.007 | 0.274 | 0.028 | ||
| Taraxacum officinale | 0.900 | 0.484 | 0.660 | 0.002 | 0.240 | 0.002 | ||
| Hieracium gracile | 1.000 | 0.194 | 0.440 | 0.024 | 0.327 | 0.024 | ||
| UNLOGGED | Vaccinium membranaceum | 0.958 | 0.742 | 0.843 | 0.001 | 0.360 | <0.001 | |
| Cornus canadensis | - | - | - | - | 0.233 | <0.050 |
| Species | LOGGED Mean | UNLOGGED Mean | Adjusted p-Values | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Height (cm) | Length (cm) | Width (cm) | Leaf Area (cm2) | Height (cm) | Length (cm) | Width (cm) | Leaf Area (cm2) | Height | Length | Width | Leaf Area | ||
| A | Chamerion angustifolium | 25.3 | 15.3 | 14.1 | 9.6 | 40.4 | 19.7 | 18.4 | 14.1 | 0.015 | 0.001 | 0.002 | 0.002 |
| Cornus canadensis | 3.7 | 4.3 | 3.6 | 7.4 | 3.7 | 4.5 | 3.8 | 8.4 | 0.505 | 0.719 | 0.719 | 0.451 | |
| Vaccinium membranaceum | 5.1 | 7.2 | 5.4 | 1.7 | 5.8 | 8.9 | 6.8 | 1.8 | 0.505 | 0.317 | 0.236 | 0.719 | |
| Spiraea betulifolia | 15.5 | 16.5 | 13.2 | 4.1 | 24.5 | 22.3 | 17.1 | 6.3 | 0.001 | 0.005 | 0.022 | 0.003 | |
| Arnica cordifolia | 11.7 | 8.3 | 5.8 | 9.3 | 12.1 | 10.1 | 7.6 | 13.4 | 0.976 | 0.133 | 0.095 | 0.095 | |
| B | Aster foliaceus | 21.2 | 13.2 | 10.9 | 11.3 | 22.0 | 15.3 | 13.3 | 11.7 | 0.957 | 0.171 | 0.335 | 0.829 |
| Petasites frigidus | 6.5 | 14.7 | 10.7 | 24.5 | 5.8 | 12.8 | 9.9 | 18.0 | 0.684 | 0.656 | 0.731 | 0.236 | |
| Taraxacum officinale | 12.8 | 19.6 | 15.1 | 38.9 | 17.3 | 14.3 | 7.8 | 23.4 | 0.451 | 0.676 | 0.095 | 0.440 | |
| Rubus idaeus | 8.0 | 12.4 | 9.5 | 9.6 | 8.0 | 12.5 | 9.2 | 9.9 | 0.987 | 0.987 | 0.976 | 0.979 | |
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ton, M.; Krawchuk, M.A. The Effects of Disturbance History on Ground-Layer Plant Community Composition in British Columbia. Forests 2016, 7, 109. https://doi.org/10.3390/f7050109
Ton M, Krawchuk MA. The Effects of Disturbance History on Ground-Layer Plant Community Composition in British Columbia. Forests. 2016; 7(5):109. https://doi.org/10.3390/f7050109
Chicago/Turabian StyleTon, Michael, and Meg A. Krawchuk. 2016. "The Effects of Disturbance History on Ground-Layer Plant Community Composition in British Columbia" Forests 7, no. 5: 109. https://doi.org/10.3390/f7050109