
Evidence on the Adaptive Recruitment of Chinese Cork Oak (Quercus variabilis Bl.): Influence on Repeated Germination and Constraint Germination by Food-Hoarding Animals

Abstract
:1. Introduction
2. Methods and Materials
2.1. Study Area
2.2. Seed Collection and Preservation
2.3. Animal Trapping
2.4. Germinated Acorns Cultivation
2.5. Radicle-Excision Behavior of A. Peninsulae
2.6. Seedling Establishment from Radicle-Excised Acorns
2.7. Data Analysis
3. Results
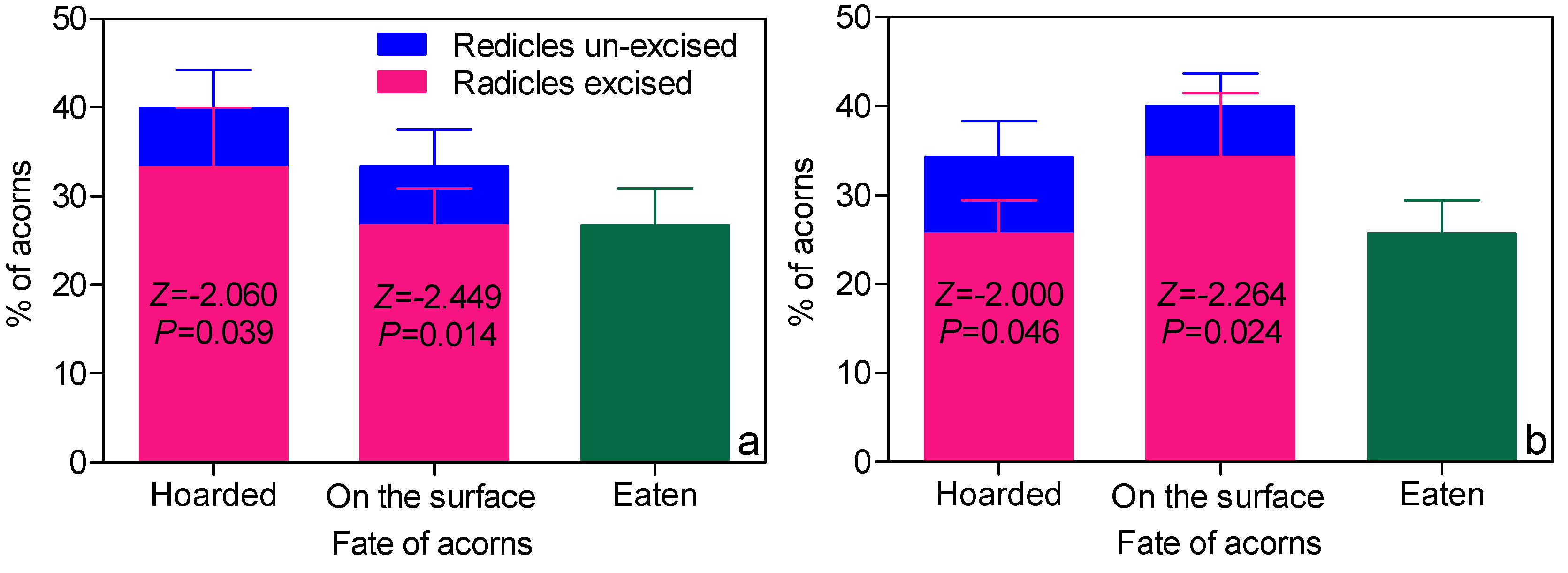
3.1. Radicle-Excision Behavior of A. Peninsulae
3.2. Germination Rates in Sound and Radicle-Excised Acorns
3.3. Germination Dynamics in Sound and Radicle-Excised Acorns
3.4. Comparison of Parameters of Seedlings Derived from Sound and Radicle-Excised Acorns
3.4.1. Seedling Stem
3.4.2. Seedling Biomass
3.4.3. Development of Seedling Root
4. Discussions
4.1. Radicle-Excision Behavior of Rodents
4.2. Renewable Germination and Seedling Establishment of Radicle-Excised Acorns
4.3. Adaptive Recruitment of White Oak in Temperate Area
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Pimm, S.L.; Raven, P. Biodiversity: Extinction by numbers. Nature 2000, 403, 843–845. [Google Scholar] [CrossRef] [PubMed]
- Gardner, T.A.; Barlow, J.; Chazdon, R.; Ewers, R.M.; Harvey, C.A.; Peres, C.A.; Sodhi, N.S. Prospects for tropical forest biodiversity in a human-modified world. Ecol. Lett. 2009, 12, 561–582. [Google Scholar] [CrossRef] [PubMed]
- Guarino, E.D.S.G.; Scariot, A. Direct seeding of dry forest tree species in abandoned pastures: Effects of grass canopy and seed burial on germination. Ecol. Res. 2014, 29, 473–482. [Google Scholar] [CrossRef]
- Holl, K.D.; Lulow, M.E. Effects of species, habitat, and distance from edge on post-dispersal seed predation in a tropical rainforest. Biotropica 1997, 29, 459–468. [Google Scholar] [CrossRef]
- Holl, K.D. Factors limiting tropical rain forest regeneration in abandoned pasture: Seed rain, seed germination, microclimate, and soil. Biotropica 1999, 31, 229–242. [Google Scholar] [CrossRef]
- Desantis, R.D.; Hallgren, S.W.; Stahle, D.W. Drought and fire suppression lead to rapid forest composition change in a forest-prairie ecotone. For. Ecol. Manag. 2011, 261, 1833–1840. [Google Scholar] [CrossRef]
- Daws, M.I.; Garwood, N.C.; Pritchard, H.W. Traits of recalcitrant seeds in a semi-deciduous tropical forest in Panamá: Some ecological implications. Funct. Ecol. 2005, 19, 874–885. [Google Scholar] [CrossRef]
- Sautu, A.; Baskin, J.M.; Baskin, C.C.; Condit, R.; Sautu, A.; Baskin, J.M.; Baskin, C.C.; Condit, R. Studies on the seed biology of 100 native species of trees in a seasonal moist tropical forest, Panama, Central America. For. Ecol. Manag. 2006, 234, 245–263. [Google Scholar] [CrossRef]
- Cole, R.J.; Holl, K.D.; Keene, C.L.; Zahawi, R.A. Direct seeding of late-successional trees to restore tropical montane forest. For. Ecol. Manag. 2011, 261, 1590–1597. [Google Scholar] [CrossRef]
- Stella, J.C.; Battles, J.J. How do riparian woody seedlings survive seasonal drought? Oecologia 2010, 164, 579–590. [Google Scholar] [CrossRef] [PubMed]
- Camargo, J.L.C.; Ferraz, I.D.K.; Imakawa, A.M. Rehabilitation of degraded areas of central Amazonia using direct sowing of forest tree seeds. Restor. Ecol. 2002, 10, 636–644. [Google Scholar] [CrossRef]
- Garcia Orth, X.; Martínez Ramos, M. Seed dynamics of early and late successional tree species in tropical abandoned pastures: Seed burial as a way of evading predation. Restor. Ecol. 2008, 16, 435–443. [Google Scholar] [CrossRef]
- Guo, C.R.; Lu, J.Q.; Yang, D.Z.; Zhao, L.P. Impacts of burial and insect infection on germination and seedling growth of acorns of Quercus variabilis. For. Ecol. Manag. 2009, 258, 1497–1502. [Google Scholar] [CrossRef]
- Bao, C. Synoptic Meteorology in China; China Ocean Press: Beijing, China, 1987. [Google Scholar]
- Qian, W.; Kang, H.S.; Lee, D.K. Distribution of seasonal rainfall in the East Asian monsoon region. Theor. Appl. Climatol. 2002, 73, 151–168. [Google Scholar] [CrossRef]
- Hogg, E.T.E.D.; Wein, R.W. Impacts of drought on forest growth and regeneration following fire in southwestern Yukon, Canada. Can. J. For. Res. 2005, 35, 2141–2150. [Google Scholar] [CrossRef]
- Klos, R.J.; Wang, G.G.; Bauerle, W.L.; Rieck, J.R. Drought impact on forest growth and mortality in the southeast USA: An analysis using Forest Health and Monitoring data. Ecol. Appl. 2009, 19, 699–708. [Google Scholar] [CrossRef] [PubMed]
- Zhao, X.F.; Lu, J.; Qiao, W.T.; Tang, F.Y. Dispersal and hoarding on acorns of Quercus variabilis by rodents in different habitats. Acta Theriol. Sin. 2009, 29, 160–166. [Google Scholar]
- Ma, Q.L.; Zhao, X.F.; Sun, M.Y.; Lu, J.Q.; Kong, M.C. Seasonal variations of wild apricot seed dispersal and hoarding by rodents in rehabilitated land. Chin. J. Appl. Ecol. 2010, 21, 1238–1243. [Google Scholar]
- Price, M.W.; Jenkins, S.H. Rodents as seeds consumers and dispersers. In Seed Dispersal; Murray, D.R., Ed.; Academic Press: Sydney, Australia, 1986; pp. 191–235. [Google Scholar]
- Vander Wall, S.B. Food Hoarding in Animals; University of Chicago Press: Chicago, IL, USA, 1990. [Google Scholar]
- Chang, G.; Xiao, Z.S.; Zhang, Z.B. Hoarding decisions by Edward’s long-tailed rats (Leopoldamys edwardsi) and South China field mice (Apodemus draco): The responses to seed size and germination schedule in acorns. Behav. Process. 2009, 82, 7–11. [Google Scholar] [CrossRef] [PubMed]
- Fox, J.F. Adaptation of gray squirrel behavior to autumn germination by white oak acorns. Evolution 1982, 36, 800–809. [Google Scholar] [CrossRef]
- Jansen, P.A.; Bongers, F.; Prins, H.H.T. Tropical rodents change rapidly germinating seeds into long-term food supplies. Oikos 2006, 113, 449–458. [Google Scholar] [CrossRef]
- Forget, P. Seed-dispersal of Vouacapoua americana (Caesalpiniaceae) by caviomorph rodents in French Guiana. J. Trop. Ecol. 1990, 6, 459–468. [Google Scholar] [CrossRef]
- Andersson, C.; Frost, I. Growth of Quercus robur seedlings after experimental grazing and cotyledon removal. Acta Bot. Neerl. 1996, 45, 85–94. [Google Scholar] [CrossRef]
- Steele, M.A.; Manierre, S.; Genna, T.; Contreras, T.A.; Smallwood, P.D.; Pereira, M.E. The innate basis of food-hoarding decisions in grey squirrels: Evidence for behavioural adaptations to the oaks. Anim. Behav. 2006, 71, 155–160. [Google Scholar] [CrossRef]
- Fleck, D.C.; Layne, J.N. Variation in tannin activity of acorns of seven species of central Florida oaks. J. Chem. Ecol. 1990, 16, 2925–2934. [Google Scholar] [CrossRef] [PubMed]
- Chung-MacCoubrey, A.L.; Hagerman, A.E.; Kirkpatrick, R.L. Effects of tannins on digestion and detoxification activity in gray squirrels (Sciurus carolinensis). Physiol. Zool. 1997, 70, 270–277. [Google Scholar] [CrossRef] [PubMed]
- Yi, X.F.; Curtis, R.; Bartlow, A.W.; Agosta, S.J.; Steele, M.A. Ability of chestnut oak to tolerate acorn pruning by rodents. Naturwissenschaften 2013, 100, 81–90. [Google Scholar] [CrossRef] [PubMed]
- Cao, L.; Xiao, Z.S.; Wang, Z.Y.; Guo, C.; Chen, J.; Zhang, Z.B. High regeneration capacity helps tropical seeds to counter rodent predation. Oecologia 2011, 166, 997–1007. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.Q.; Yi, X.F.; Yu, F. Repeated radicle pruning of Quercus mongolica acorns as a cache management tactic of Siberian chipmunks. Acta Ethol. 2012, 15, 9–14. [Google Scholar] [CrossRef]
- Yi, X.F.; Yang, Y.Q.; Curtis, R.; Bartlow, A.W.; Agosta, S.J.; Steele, M.A. Alternative strategies of seed predator escape by early-germinating oaks in Asia and North America. Ecol. Evol. 2012, 2, 487–492. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Zhang, Y.F.; Wang, Z.L.; Qiao, W.T.; Lu, J.Q. Seed dispersal and hoarding of three species by rodents in the Mt. Taihangshan Area, Jiyuan, China. Acta Theriol. Sin. 2013, 33, 150–156. [Google Scholar]
- Seiwa, K.; Watanabe, A.; Saitoh, T.; Kannu, H.; Akasaka, S. Effects of burying depth and seed size on seedling establishment of Japanese chestnuts, Castanea crenata. For. Ecol. Manag. 2002, 164, 149–156. [Google Scholar] [CrossRef]
- Perea, R.; San Miguel, A.; Gil, L. Leftovers in seed dispersal: Ecological implications of partial seed consumption for oak regeneration. J. Ecol. 2011, 99, 194–201. [Google Scholar] [CrossRef] [Green Version]
- Lu, J.Q.; Zhang, Z.B. Differentiation in seed hoarding among three sympatric rodent species in a warm temperate forest. Integr. Zool. 2008, 3, 134–142. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.B.; Wang, F.S. Effect of burial on acorn survival and seedling recruitment of Liaodong oak (Quercus liaotungensis) under rodent predation. Acta Theriol. Sin. 2001, 21, 35–43. [Google Scholar]
- Xiao, Z.; Jansen, P.A.; Zhang, Z. Using seed-tagging methods for assessing post-dispersal seed fate in rodent-dispersed trees. For. Ecol. Manag. 2006, 223, 18–23. [Google Scholar] [CrossRef]
- Xiao, Z.S.; Zhang, Z.B.; Wang, Y.S. Dispersal and germination of big and small nuts of Quercus serrata in a subtropical broad-leaved evergreen forest. For. Ecol. Manag. 2004, 195, 141–150. [Google Scholar] [CrossRef]
- Zhang, H.; Chen, Y.; Zhang, Z. Differences of dispersal fitness of large and small acorns of Liaodong oak (Quercus liaotungensis) before and after seed caching by small rodents in a warm temperature forest, China. For. Ecol. Manag. 2008, 255, 1243–1250. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Seedling Types | Lateral Roots (Number/Percentage) | |||
|---|---|---|---|---|
| Zero | One | Two | Three | |
| Sound acorns | 24/58.54% | 11/26.83% | 5/12.20% | 1/2.44% |
| Radicle-excised acorns | 10/31.25% | 14/43.75% | 2/6.25% | 6/18.75% |
| p-Value | p = 0.003 | p = 0.044 | p = 0.157 | p < 0.001 |
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons by Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, Y.; Shi, Y.; Sichilima, A.M.; Zhu, M.; Lu, J. Evidence on the Adaptive Recruitment of Chinese Cork Oak (Quercus variabilis Bl.): Influence on Repeated Germination and Constraint Germination by Food-Hoarding Animals. Forests 2016, 7, 47. https://doi.org/10.3390/f7020047
Zhang Y, Shi Y, Sichilima AM, Zhu M, Lu J. Evidence on the Adaptive Recruitment of Chinese Cork Oak (Quercus variabilis Bl.): Influence on Repeated Germination and Constraint Germination by Food-Hoarding Animals. Forests. 2016; 7(2):47. https://doi.org/10.3390/f7020047
Chicago/Turabian StyleZhang, Yifeng, Yuhua Shi, Alfred M Sichilima, Meilin Zhu, and Jiqi Lu. 2016. "Evidence on the Adaptive Recruitment of Chinese Cork Oak (Quercus variabilis Bl.): Influence on Repeated Germination and Constraint Germination by Food-Hoarding Animals" Forests 7, no. 2: 47. https://doi.org/10.3390/f7020047