
Radial Growth Response of Larix gmelinii to Climate along a Latitudinal Gradient in the Greater Khingan Mountains, Northeastern China

Abstract
:1. Introduction
2. Materials and Methods
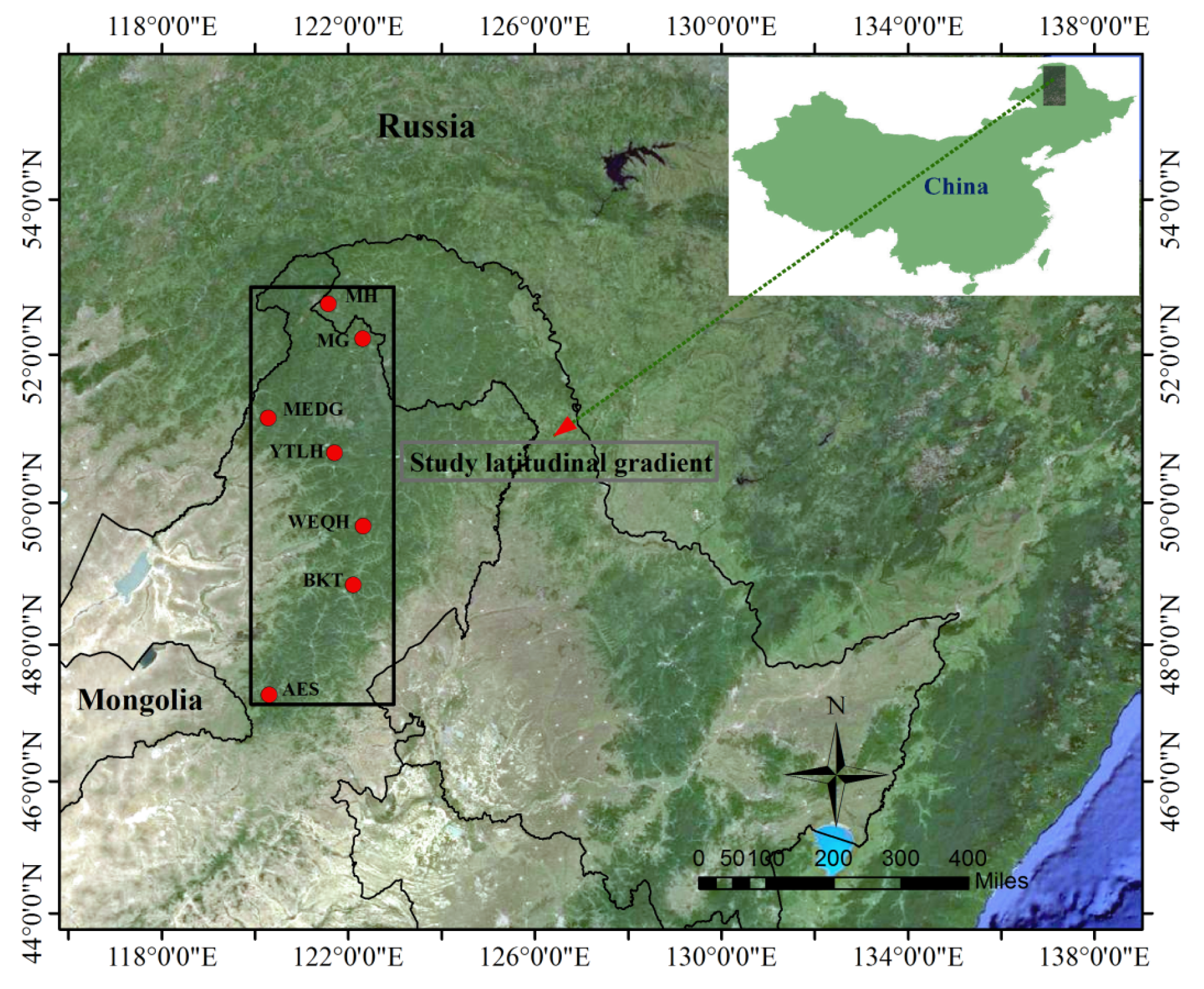
2.1. Study Area
2.2. Tree-Ring Sampling and Chronology Development
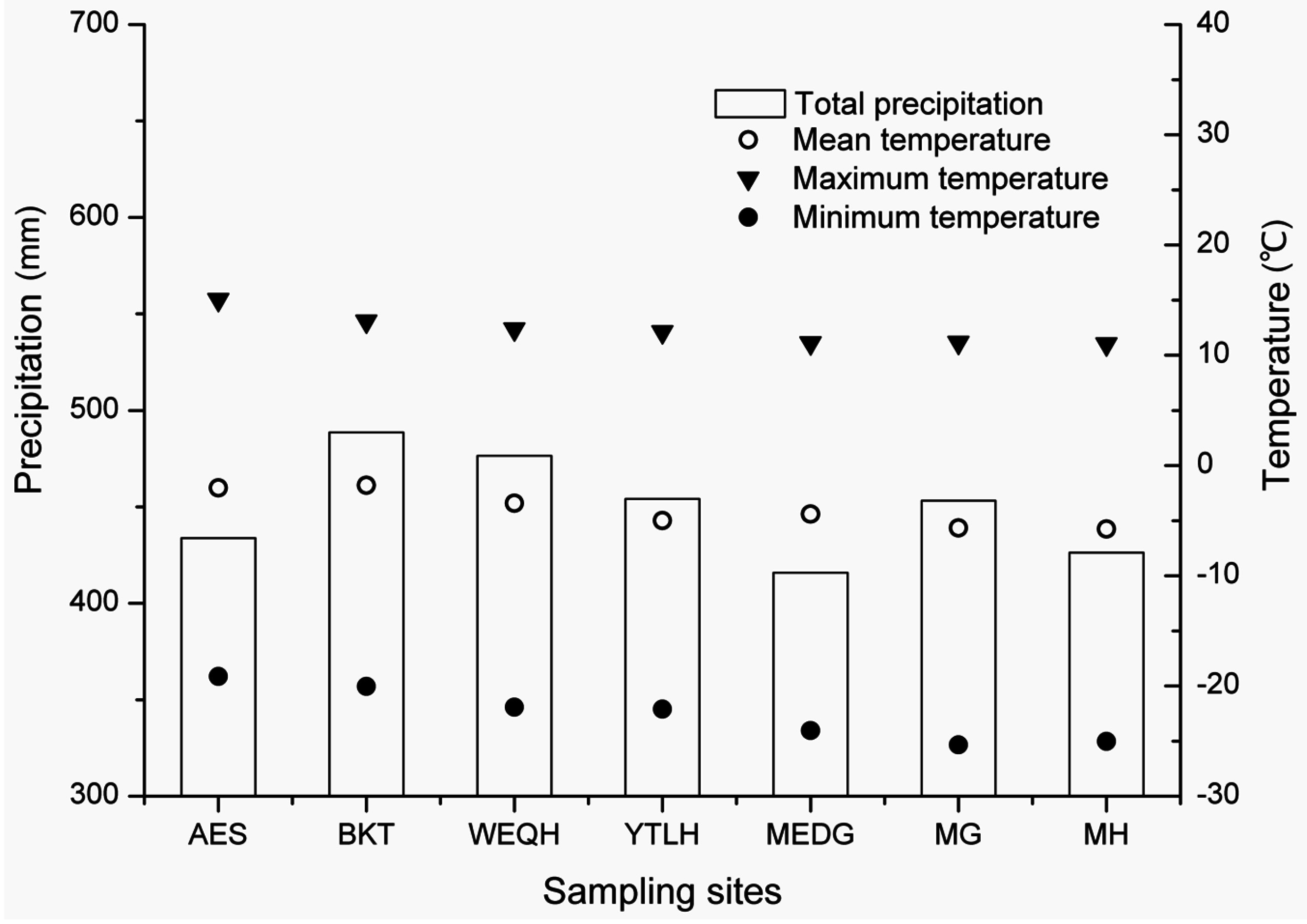
2.3. Climate Data
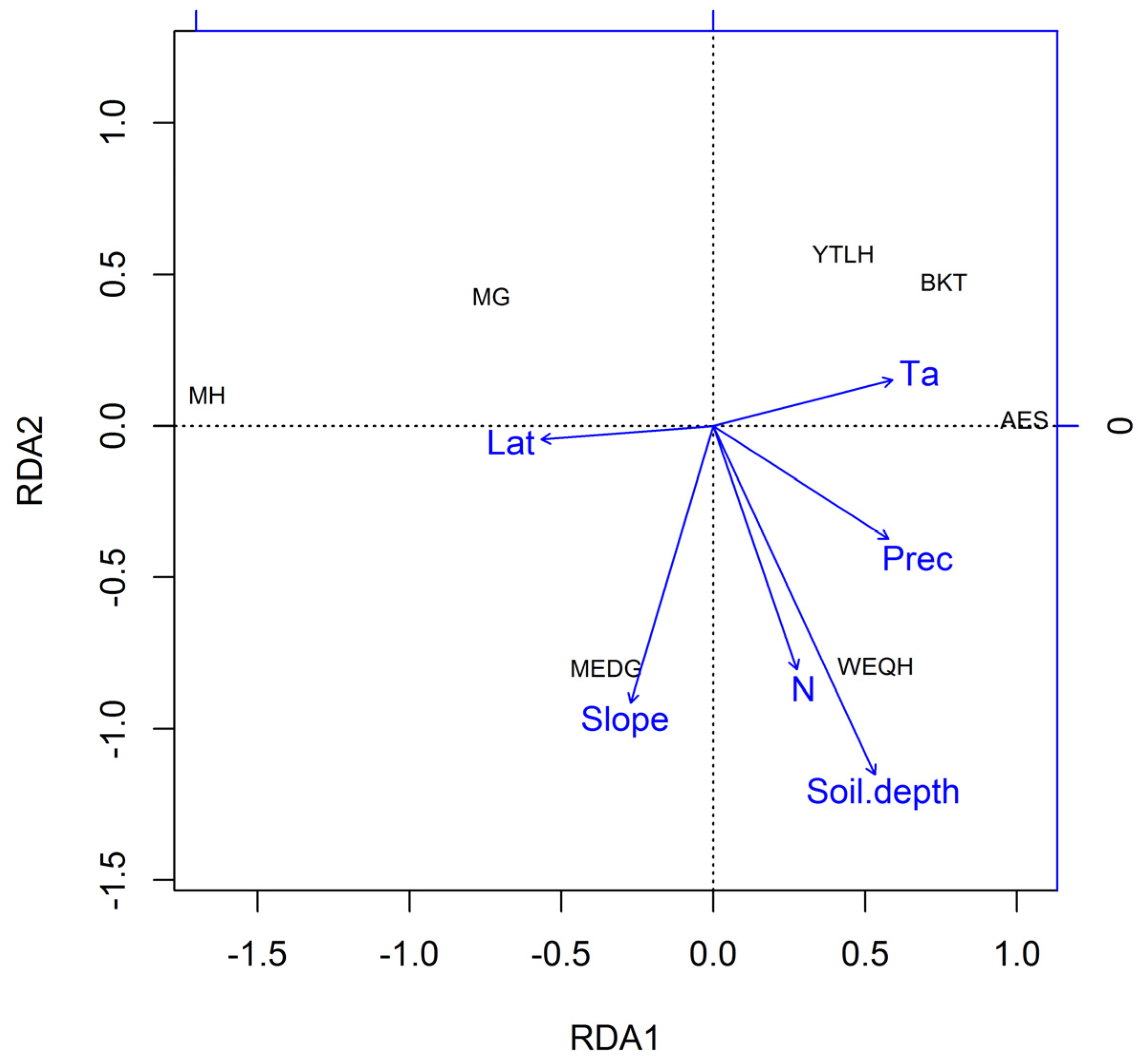
2.4. Climate–Growth Relationship Analysis
2.5. Definition of Regional Patterns
3. Results
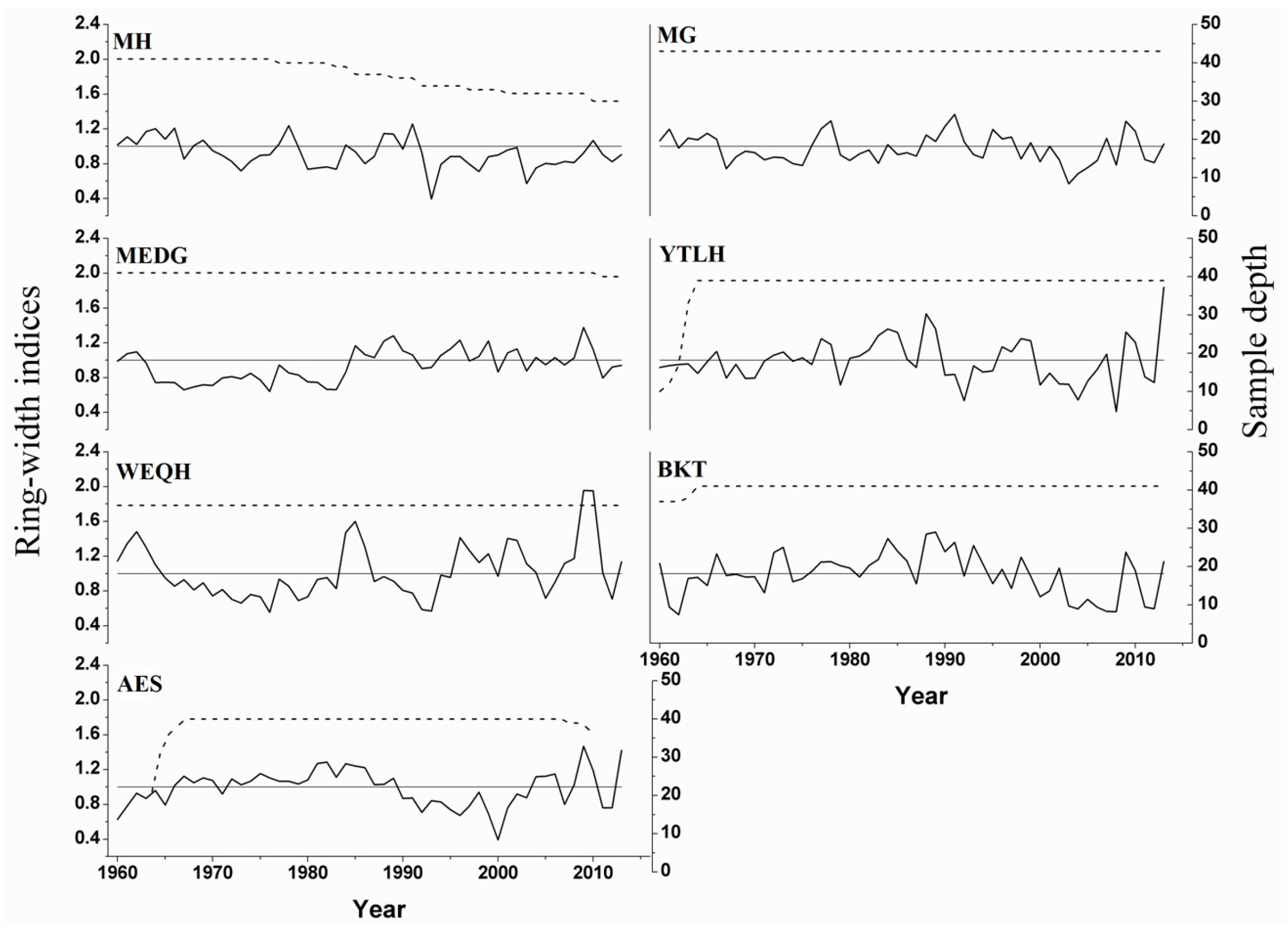
3.1. Chronology Statistics
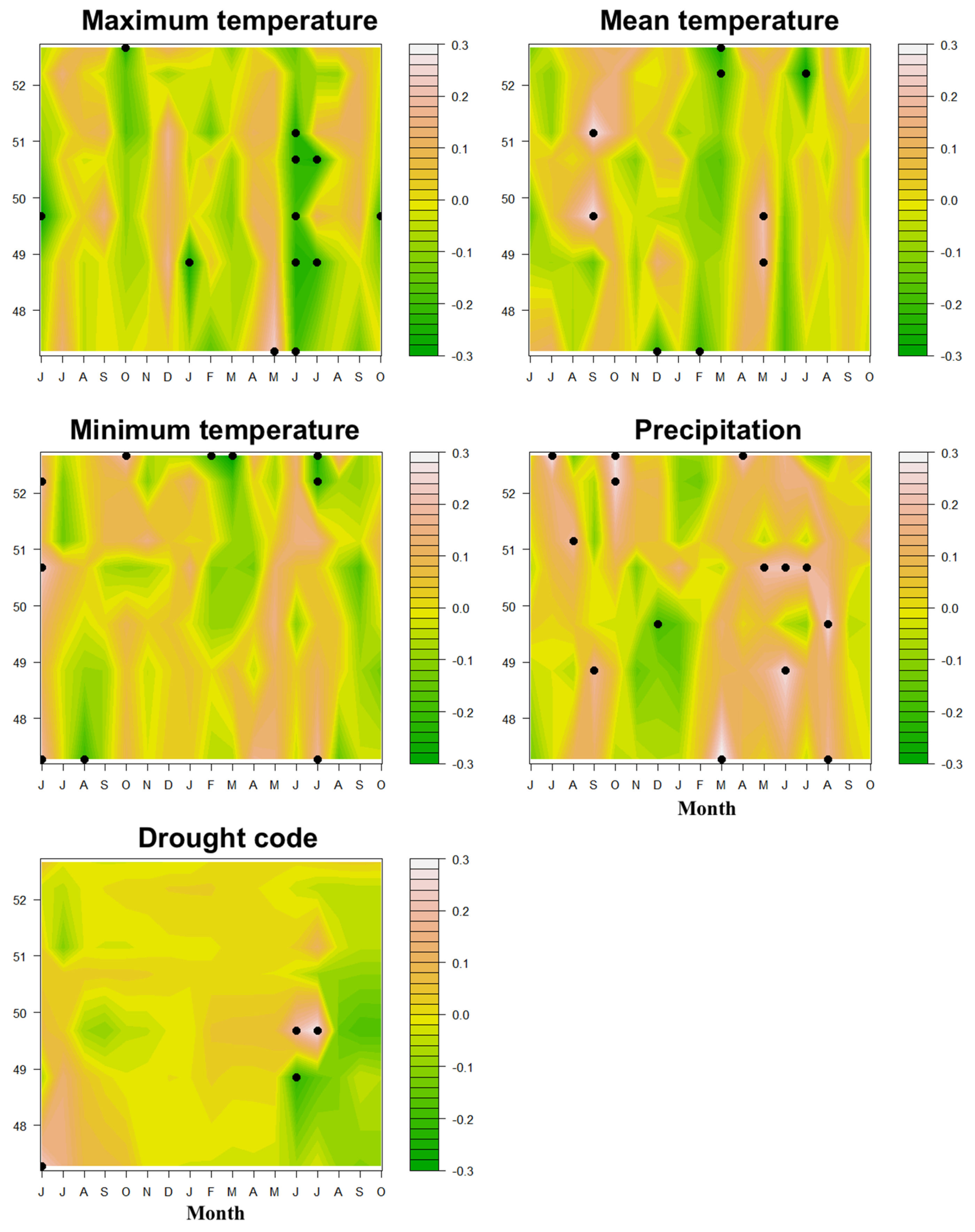
3.2. Radial Growth–Climate Associations
3.3. Spatial Patterns of Climate–Growth Responses
4. Discussion
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Fritts, H.C. Tree Rings and Climate; Academic Press: New York, NY, USA, 1976. [Google Scholar]
- Zhang, X.; Chen, Z. A new method to remove the tree growth trend based on ensemble empirical mode decomposition. Trees 2015, 1–9. [Google Scholar] [CrossRef]
- Liang, E.Y.; Shao, X.M.; Eckstein, D.; Liu, X. Spatial variability of tree growth along a latitudinal transect in the Qilian Mountains, northeastern Tibetan Plateau (EI). Can. J. For. Res. 2010, 40, 200–211. [Google Scholar] [CrossRef]
- Yu, D.; Wang, Q.; Wang, Y.; Zhou, W.; Ding, H.; Fang, X.; Dai, L. Climatic effects on radial growth of major tree species on Changbai Mountain. Ann. For. Sci. 2011, 68, 921–933. [Google Scholar] [CrossRef]
- Thuiller, W.; Lavorel, S.; Arujo, M.B.; Sykes, M.T.; Prentice, I.C. Climate change threats to plant diversity in Europe. Proc. Natl. Acad. Sci. 2005, 102, 8245–8250. [Google Scholar] [CrossRef] [PubMed]
- Tardif, J.C.; Conciatori, F.; Nantel, P.; Gagnon, D. Radial growth and climate responses of white oak (Quercus alba) and northern red oak (Quercus rubra) at the northern distribution limit of white oak in Quebec, Canada. J. Biogeogr. 2006, 33, 1657–1669. [Google Scholar] [CrossRef]
- Metsaranta, J.M.; Lieffers, V.J.; Wein, R.W. Dendrochronological reconstruction of jack pine snag and downed log dynamics in Saskatchewan and Manitoba, Canada. For. Ecol. Manag. 2008, 255, 1262–1270. [Google Scholar] [CrossRef]
- Huang, J.; Tardif, J.C.; Bergeron, Y.; Denneler, B.; Berninger, F.; Girardin, M.P. Radial growth response of four dominant boreal tree species to climate along a latitudinal gradient in the eastern Canadian boreal forest. Glob. Chang. Biol. 2010, 16, 711–731. [Google Scholar] [CrossRef]
- Wang, X.M.; Zhao, X.H.; Gao, L.S. Climatic response of Betula ermanii along an altitudinal gradient in the northern slope of Changbai Mountain, China. Dendrobiology 2013, 70, 99–107. [Google Scholar] [CrossRef]
- Peng, X.; Xiao, S.; Cheng, G.; Xiao, H.; Tian, Q.; Zhang, Q. Human activity impacts on the stem radial growth of Populus euphratica, riparian forests in China’s Ejina oasis, using tree-ring analysis. Trees 2015, 1–14. [Google Scholar] [CrossRef]
- Griesbauer, H.P.; Green, D.S. Geographic and temporal patterns in white spruce climate–growth relationships in Yukon, Canada. For. Ecol. Manag. 2012, 267, 215–227. [Google Scholar] [CrossRef]
- Li, Z.; Liu, G.; Fu, B.; Zhang, Q.; Ma, K.; Pederson, N. The growth-ring variations of alpine shrub Rhododendron przewalskii reflect regional climate signals in the alpine environment of Miyaluo town in western Sichuan province, China. Acta Ecol. Sinica 2013, 33, 23–31. [Google Scholar] [CrossRef]
- Zyryanova, O.A.; Yaborov, V.T.; Tchikhacheva, T.L.; Koike, T.; Makoto, K.; Matsuura, Y.; Zyryanov, V.I. The structure and biodiversity after fire disturbance in Larix gmelinii (Rupr.). Eurasian J. For. Res. 2007, 10, 19–29. [Google Scholar]
- Farjon, A. Pinaceae; Koeltz Scientific Books: Konigstein, Germany, 1990. [Google Scholar]
- Fu, L.K.; Li, N.; Mill, R.R. Larix. In Flora of China; Wu, Z.Y., Raven, P.H., Eds.; Science Press: Beijing, China; Missouri Botanical Garden Press: St. Louis, MI, USA, 1999; pp. 33–37. [Google Scholar]
- Cheng, X.X.; Yan, X.D. Effects of global climate change on forest succession in Daxing’anling of Northeast China. Chin. J. Ecol. 2007, 26, 1277–1284. (In Chinese) [Google Scholar]
- Leng, W.F.; He, H.S.; Bu, R.C.; Hu, Y.M. Sensitivity analysis of the impacts of climate change on potential distribution of three larch (Larix) species in northeastern China. J. Plant Ecol. 2007, 31, 825–833. (In Chinese) [Google Scholar]
- Cheng, X.X.; Yan, X.D. Effects of climate change on typical forest in the northeast of China. Acta Ecol. Sinica. 2008, 28, 534–543. (In Chinese) [Google Scholar]
- Qian, W.; Qin, A. Spatial-temporal characteristics of temperature variation in China. Meteorol. Atmos. Phys. 2006, 93, 1–16. [Google Scholar] [CrossRef]
- Wang, L.; Shao, X.; Huang, L.; Liang, E. Tree-ring characteristics of Larix gmelinii and Pinus sylvestris var. Mongolica and their response to climate in Mohe, China. Chin. J. Plant Ecol. 2005, 29, 380–385. (In Chinese) [Google Scholar] [CrossRef]
- Wang, X.; Zhang, Y.; Mcrae, D.J. Spatial and age-dependent tree-ring growth responses of Larix gmelinii to climate in northeastern China. Trees 2009, 23, 875–885. [Google Scholar] [CrossRef]
- Zhang, X.; Bai, X.; Chang, Y.; Chen, Z. Increased sensitivity of Dahurian larch radial growth to summer temperature with the rapid warming in Northeast China. Trees 2016, 1–8. [Google Scholar] [CrossRef]
- Hutchinson, M.F. ANUSPLIN Version 4.3. Center for Resource and Environmental Studies, Australian National University: Canberra, Australia, 2004. Available online: http://fennerschool. anu. edu. au/ publications/ software/anusplin.php (accessed on 12 December 2008).
- Holmes, R.L. Computer-assisted quality control in tree-ring dating and measurement. Tree-Ring Bull. 1983, 44, 69–75. [Google Scholar]
- Cook, E.R.; Briffa, K.R. Data analysis. In Methods of Dendrochronology; Cook, E.R., Kairiukstis, L.A., Eds.; Kluwer: Boston, MA, USA, 1990; pp. 97–162. [Google Scholar]
- Cook, E.R. A time series approach to tree-ring standardization. Ph.D. Thesis, University of Arizona, Tucson, AZ, USA, 1985. [Google Scholar]
- Wigley, T.M.L.; Briffa, K.R.; Jones, P.D. On the average value of correlated time series, with applications in dendroclimatology and hydrometeorology. J. Clim. Appl. Meteorol. 1984, 23, 201–213. [Google Scholar] [CrossRef]
- Chinese Meteorological Data Sharing Service System. Available online: http://www.cma.gov.cn (accessed on 16 September 2015).
- Global Historical Climatology Network. Available online: https://www.ncdc.noaa.gov (accessed on 20 September 2015).
- Huang, C.; Zheng, X.; Tait, A.; Dai, Y.; Yang, C.; Chen, Z.; Wang, Z. On using smoothing spline and residual correction to fuse rain gauge observations and remote sensing data. J. Hydrol. 2013, 508, 410–417. [Google Scholar] [CrossRef]
- Li, T.; Zheng, X.; Dai, Y.; Yang, C.; Chen, Z.; Zhang, S.; Liao, R. Mapping near-surface air temperature, pressure, relative humidity and wind speed over Mainland China with high spatiotemporal resolution. Adv. Atmos. Sci. 2014, 31, 1127–1135. [Google Scholar] [CrossRef]
- Biondi, F.; Waikul, K. DENDROCLIM 2002: AC ++ program for statistical calibration of climate signals in tree-ring chronologies. Comput. Geosci. 2004, 30, 303–311. [Google Scholar] [CrossRef]
- Legendre, P.; Legendre, L. Numerical Ecology; Elsevier Scientific: New York, NY, USA, 1998. [Google Scholar]
- Braak, C.J.F.T.; Prentice, I.C. A theory of gradient analysis. Adv. Ecol. Res. 1988, 18, 271–317. [Google Scholar] [CrossRef]
- TerBraak, C.J.F.; Smilauer, P. CANOCO Reference Manual and Cano Draw for User’s Guide Windows: Software for Canonical Community Ordination, Version 4.5. Microcomputer Power: Ithaca, NY, USA, 2002. [Google Scholar]
- Mäkinen, H.; Nöjd, P.; Kahle, H.P.; Neumann, U.; Tveite, B.; Mielikäinen, K.; Spiecker, H. Radial growth variation of Norway spruce (Picea abies (L.) Karst.) across latitudinal and altitudinal gradients in central and northern Europe. For. Ecol. Manag. 2002, 171, 243–259. [Google Scholar] [CrossRef]
- Lloyd, A.H.; Bunn, A.G.; Berner, L. A latitudinal gradient in tree growth response to climate warming in the Siberian taiga. Glob. Chang. Biol. 2010, 17, 1935–1945. [Google Scholar] [CrossRef]
- Goodine, G.K.; Lavigne, M.B.; Krasowski, M.J. Spring time resumption of photosynthesis in balsam fir (Abiesbalsamea). Tree Physiol. 2008, 28, 1069–1076. [Google Scholar] [CrossRef] [PubMed]
- LaMarche, V.C., Jr. Palecoclimatic inferences from long tree-ring records. Science 1974, 183, 1043–1048. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Cui, M.X.; Ma, Y.J.; Wu, T.; Chen, Z.J.; Ding, W.H. Larix gmelinii tree-ring width chronology and its responses to climate change in Kuduer, Great Xing’an mountains. Chin. J. Appl. Ecol. 2010, 21, 2501–2507. [Google Scholar]
- Fritts, H.C.; Smith, D.G.; Cardis, J.W.; Budelsky, C.A. Tree-ring Characteristics along a vegetation gradient in northern Arizona. Ecology 1965, 46, 393–401. [Google Scholar] [CrossRef]
- Su, H.; Axmacher, J.C.; Yang, B.; Sang, W. Differential radial growth response of three coexisting dominant tree species to local and large-scale climate variability in a subtropical evergreen broad-leaved forest of China. Ecol. Res. 2015, 30, 745–754. [Google Scholar] [CrossRef]
- Graumlich, L.J. Response of tree growth to climatic variation in the mixed conifer and deciduous forest of the upper Great Lakes region. Can. J. For. Res. 1993, 23, 133–143. [Google Scholar] [CrossRef]
- Suni, T.; Berninger, F.; Vesala, T. Air temperature triggers the recovery of evergreen boreal forest photosynthesis in spring. Glob. Chang. Biol. 2003, 9, 1410–1426. [Google Scholar]
- Savva, Y.; Bergeron, Y.; Denneler, B.; Koubaa, A.; Tremblay, F. Effect of interannual climate variations on radial growth of jack pine provenances in Petawawa, Ontario. Can. J. For. Res. 2008, 38, 619–630. [Google Scholar] [CrossRef]
- Butt, N.; Bebber, D.P.; Riutta, T.; Crockatt, M.; Morecroft, M.D.; Malhi, Y. Relationships between tree growth and weather extremes: Spatial and interspecific comparisons in a temperate broadleaf forest. For. Ecol. Manag. 2014, 334, 209–216. [Google Scholar] [CrossRef]
- George, S.S.; Ault, T.R. The imprint of climate within Northern Hemisphere trees. Quat. Sci. Rev. 2014, 89, 1–4. [Google Scholar] [CrossRef]
- Vaganov, E.A.; Hughes, M.K.; Kirdyanov, A.V.; Schweingruber, F.H.; Silkin, P.P. Influence of snowfall and melt timing on tree growth in subarctic Eurasia. Nature 1999, 400, 149–151. [Google Scholar]
- Li, B.; Zhou, X.; Zhao, Z.; Wang, J.; Wang, S.; Shang, K.; Li, P. Change features of different types and grades of precipitation event in northeast China in recent 50 years. Plateau Meteorol. 2013, 32, 1414–1424. (In Chinese) [Google Scholar]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Site Code | MH | MG | MEDG | YTLH | WEQH | BKT | AES |
|---|---|---|---|---|---|---|---|
| Long. (E) | 121.57 | 122.3 | 120.28 | 121.7 | 122.31 | 122.1 | 120.3 |
| Lat. (N) | 52.66 | 52.21 | 51.15 | 50.68 | 49.68 | 48.85 | 47.27 |
| Elev. (a.s.l.) | 908 | 931 | 971 | 908 | 988 | 996 | 1018 |
| Slope | 10 | 10 | 0 | 12 | 20 | 0 | 0 |
| Aspect | South | South | South | South | South | South | South |
| Soil depth (cm) | 10 | 50 | 10 | 20 | 100 | 10 | 100 |
| Soil N content (g/kg) | 0.12 | 0.25 | 0.19 | 0.29 | 0.34 | 0.18 | 0.42 |
| Annual mean temperature (°C) | −5.8 | −5.6 | −4.4 | −4.9 | −3.4 | −1.8 | −2 |
| Annual total precipitation (mm) | 426 | 453 | 415 | 454 | 476 | 488 | 433 |
| Site code | MH | MG | MEDG | YTLH | WEQH | BKT | AES |
|---|---|---|---|---|---|---|---|
| SD | 0.178 | 0.275 | 0.321 | 0.229 | 0.278 | 0.145 | 0.143 |
| MS | 0.228 | 0.221 | 0.237 | 0.308 | 0.253 | 0.286 | 0.249 |
| AC1 | 0.528 | 0.590 | 0.580 | 0.680 | 0.610 | 0.460 | 0.790 |
| RBAR | 0.665 | 0.593 | 0.504 | 0.660 | 0.504 | 0.582 | 0.541 |
| EPS | 0.965 | 0.966 | 0.958 | 0.983 | 0.952 | 0.969 | 0.958 |
| PC1 | 0.369 | 0.410 | 0.390 | 0.628 | 0.280 | 0.460 | 0.730 |
| SNR | 27.355 | 28.646 | 22.785 | 56.848 | 20.012 | 30.782 | 23.089 |
| MRW (mm) | 0.77 ± 0.2 | 1.13 ± 0.4 | 0.67 ± 0.2 | 2.03 ± 0.5 | 0.68 ± 0.4 | 2.09 ± 0.5 | 3.4 ± 0.4 |
| No. of radii (trees) | 41 (22) | 43 (28) | 41 (26) | 40 (27) | 37 (25) | 40 (32) | 39 (25) |
| Period | 1919–2014 | 1902–2013 | 1763–2013 | 1941–2013 | 1863–2013 | 1939–2013 | 1955–2013 |
| AES | BKT | WEQH | YTLH | MEDG | MG | MH | |
|---|---|---|---|---|---|---|---|
| AES | 1 | 0.46 | 0.44 | 0.43 | 0.26 | 0.06 | 0.01 |
| BKT | 1 | 0.31 | 0.63 | 0.36 | 0.36 | 0.23 | |
| WEQH | 1 | 0.52 | 0.68 | 0.44 | 0.26 | ||
| YTLH | 1 | 0.39 | 0.46 | 0.21 | |||
| MEDG | 1 | 0.39 | 0.29 | ||||
| MG | 1 | 0.63 | |||||
| MH | 1 |
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jiang, Y.; Zhang, J.; Han, S.; Chen, Z.; Setälä, H.; Yu, J.; Zheng, X.; Guo, Y.; Gu, Y. Radial Growth Response of Larix gmelinii to Climate along a Latitudinal Gradient in the Greater Khingan Mountains, Northeastern China. Forests 2016, 7, 295. https://doi.org/10.3390/f7120295
Jiang Y, Zhang J, Han S, Chen Z, Setälä H, Yu J, Zheng X, Guo Y, Gu Y. Radial Growth Response of Larix gmelinii to Climate along a Latitudinal Gradient in the Greater Khingan Mountains, Northeastern China. Forests. 2016; 7(12):295. https://doi.org/10.3390/f7120295
Chicago/Turabian StyleJiang, Yangao, Junhui Zhang, Shijie Han, Zhenju Chen, Heikki Setälä, Jinghua Yu, Xingbo Zheng, Yingtao Guo, and Yue Gu. 2016. "Radial Growth Response of Larix gmelinii to Climate along a Latitudinal Gradient in the Greater Khingan Mountains, Northeastern China" Forests 7, no. 12: 295. https://doi.org/10.3390/f7120295