
The Effects of Fertilization on the Growth and Physiological Characteristics of Ginkgo biloba L.


Abstract
:1. Introduction
2. Materials and Methods
2.1. The Study Site
2.2. Materials and Experimental Design
2.3. Tree Growth Indictors’ Measurement
2.4. Determination of Photosynthetic Indexes
2.5. Determination of Nutrient Concentration in Leaves
2.6. Statistical Analysis
3. Results
3.1. Effects of Fertilizer on Growth Indicators
3.2. Effects of Fertilizer on Photosynthesis Indicators
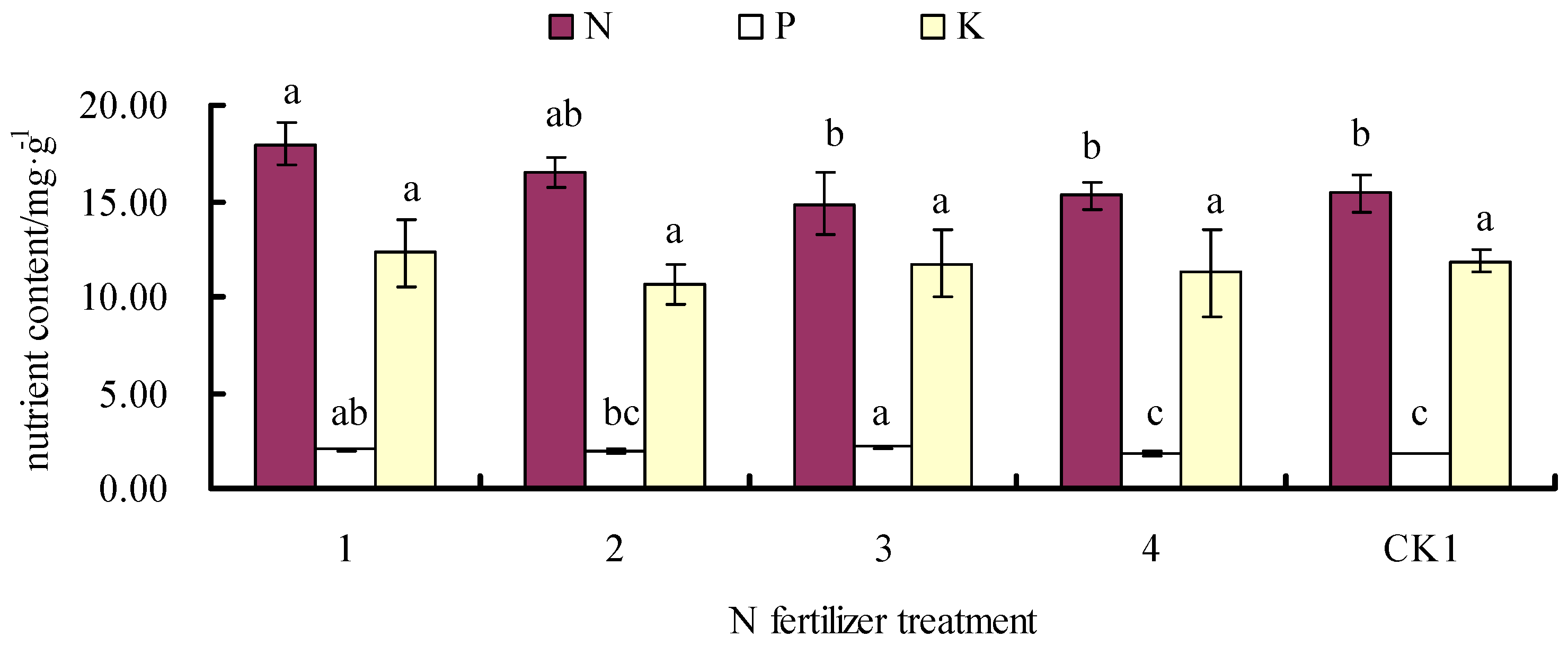
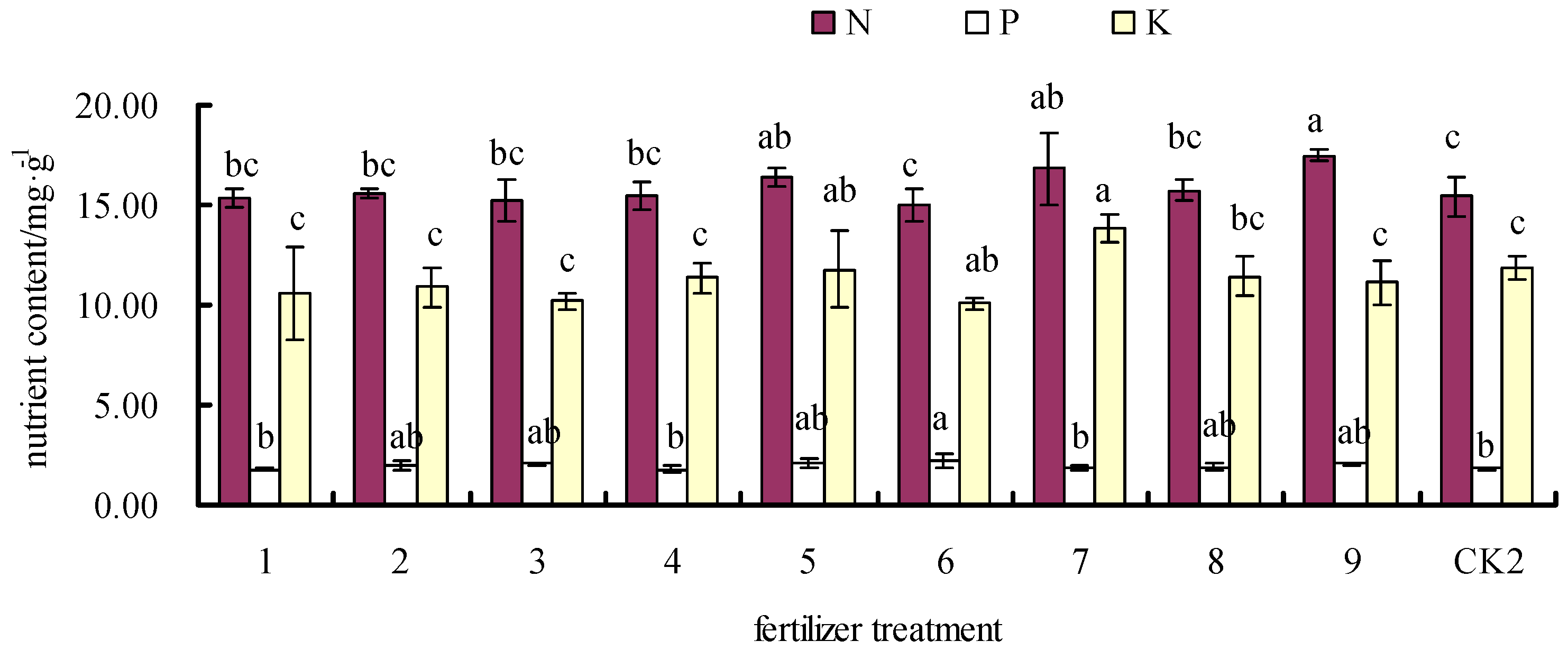
3.3. Effects of Fertilizer on Leaf N, P, and K Contents
4. Discussion
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Pathak, H.; Aggarwal, P.K.; Roetter, R.; Kalra, N.; Bandyopadhaya, S.K.; Prasad, S.; Keulch, H.V. Modelling the quantitative evaluation of soil nutrient supply, nutrient use efficiency, and fertilizer requirements of wheat in India. Nutr. Cycl. Agroecosyst. 2003, 65, 105–113. [Google Scholar] [CrossRef]
- Fox, T.R.; Allen, H.L.; Albaugh, T.J.; Rubilar, R.; Carlson, C.A. Tree Nutrition and Forest Fertilization of Pine Plantations in the Southern United States. South. J. Appl. For. 2006, 31, 5–11. [Google Scholar]
- Sophie, W.; Leigh, A.K.K.; Grayston, S.J. Effects of long-term fertilization of forest soils on potential nitrification and on the abundance and community structure of ammonia oxidizers and nitrite oxidizers. FEMS Microbiol. Ecol. 2011, 79, 142–154. [Google Scholar]
- Gough, C.M.; Seiler, J.R.; Maier, C.A. Short-term effects of fertilization on loblolly pine (Pinus taeda L.) physiology. Plant Cell Environ. 2004, 27, 876–886. [Google Scholar] [CrossRef]
- King, N.T.; Seiler, J.R.; Fox, T.R.; Johnsen, K.H. Post-fertilization physiology and growth performance of loblolly pine clones. Tree Physiol. 2008, 28, 703–711. [Google Scholar] [CrossRef] [PubMed]
- Samuelson, L.J.; Butnor, J.; Maier, C.; Stokes, T.A.; Johnsen, K.; Kane, M. Growth and physiology of loblolly pine in response to long-term resource management: Defining growth potential in the southern United States. Can. J. For. Res. 2008, 38, 721–732. [Google Scholar] [CrossRef]
- Luo, J.; Qin, J.J.; He, F.F.; Li, H.; Liu, T.X.; Polle, A.; Peng, C.H.; Luo, Z.B. Net fluxes of ammonium and nitrate in association with H+ fluxes in fine roots of Populus popularis. Planta 2013, 237, 19–31. [Google Scholar] [CrossRef] [PubMed]
- Güsewell, S. N:P ratios in terrestrial plants: Variation and functional significance. New Phytol. 2004, 164, 243–266. [Google Scholar] [CrossRef]
- Warren, C.R. How does P affect photosynthesis and metabolite profiles of Eucalyptus globulus? Tree Physiol. 2011, 31, 727–739. [Google Scholar] [CrossRef] [PubMed]
- Coskun, D.; Britto, D.T.; Kronzucker, H.J. The physiology of channel-mediated K+, acquisition in roots of higher plants. Physiol. Plant. 2014, 151, 305–312. [Google Scholar] [CrossRef] [PubMed]
- Battie-Laclau, P.; Laclau, J.P.; Beri, C.; Mietton, L.; Muniz, M.R.A.; Arenque, B.C.; Piccolo, M.C.; Jordan-Meille, L.; Bouillet, J.P.; Nouvellon, Y. Photosynthetic and anatomical responses of Eucalyptus grandis leaves to potassium and sodium supply in a field experiment. Plant Cell Environ. 2014, 37, 70–81. [Google Scholar] [CrossRef] [PubMed]
- Christina, M.; Maire, G.L.; Battie-Laclau, P.; Nouvellon, Y.; Bouillet, J.P.; Jourdan, C.; Goncalves, J.L.M.; Laclau, J.P. Measured and modeled interactive effects of potassium deficiency and water deficit on gross primary productivity and light-use efficiency in Eucalyptus grandis plantations. Glob. Chang. Biol. 2015, 21, 2022–2039. [Google Scholar] [CrossRef] [PubMed]
- Mäser, P.; Gierth, M.; Schroeder, J.I. Molecular mechanisms of potassium and sodium uptake in plants. Plant Soil 2002, 247, 43–54. [Google Scholar] [CrossRef]
- Cakmak, I. The role of potassium in alleviating detrimental effects of abiotic stresses in plants. J. Plant Nutr. Soil Sci. 2005, 168, 521–530. [Google Scholar] [CrossRef]
- Li, X.Y.; Mu, C.S.; Lin, J.X.; Wang, Y.; Li, X.J. Effect of alkaline potassium and sodium salts on growth, photosynthesis, ions absorption and solutes synthesis of wheat seedlings. Exp. Agric. 2014, 50, 144–157. [Google Scholar] [CrossRef]
- Albaugh, T.J.; Allen, H.L.; Dougherty, P.M.; Kress, L.W.; King, J.S. Leaf area and above- and belowground growth responses of loblolly pine to nutrient and water additions. For. Sci. 1998, 44, 317–328. [Google Scholar]
- Balster, N.J.; Marshall, J.D. Eight-year responses of light interception, effective leaf area index, and stemwood production in fertilized stands of interior Douglas-fir (Pseudotsuga menziesii var. glauca). Can. J. For. Res. 2000, 30, 733–743. [Google Scholar] [CrossRef]
- Samuelson, L.; Stokes, T.; Cooksey, T. Production efficiency of loblolly pine and sweetgum in response to four years of intensive management. Tree Physiol. 2001, 21, 369–376. [Google Scholar] [CrossRef] [PubMed]
- Gotore, T.; Murepa, R.; Gapare, W.J. Effects of Nitrogen, Phosphorus and Potassium on the Early Growth of Pinus patula and Eucalyptus grandis. J. Trop. For. Sci. 2014, 26, 22–31. [Google Scholar]
- Salifu, K.F.; Jacobs, D.F.; Birge, Z.K.D. Nursery nitrogen loading improves field performance of bareroot oak seedlings planted on abandoned mine lands. Restor. Ecol. 2009, 17, 339–349. [Google Scholar] [CrossRef]
- Li, H.; Li, M.; Luo, J.; Cao, X.; Qu, L.; Gai, Y.; Jiang, X.; Liu, T.; Bai, H.; Janz, D. N fertilization has different effects on the growth, carbon and nitrogen physiology, and wood properties of slow-and fast-growing Populus species. J. Exp. Bot. 2012, 63, 6173–6185. [Google Scholar] [CrossRef] [PubMed]
- Luo, J.; Li, H.; Liu, T.; Polle, A.; Peng, C.; Luo, Z. Nitrogen metabolism of two contrasting poplar species during acclimation to limiting nitrogen availability. J. Exp. Bot. 2013, 64, 4207–4224. [Google Scholar] [CrossRef] [PubMed]
- Miller, B.D.; Hawkins, B.J. Ammonium and nitrate uptake, nitrogen productivity and biomass allocation in interior spruce families with contrasting growth rates and mineral nutrient preconditioning. Tree Physiol. 2007, 27, 901–909. [Google Scholar] [CrossRef] [PubMed]
- Bown, H.E.; Watt, M.S.; Clinton, P.W.; Mason, E.G.; Whitehead, D. The influence of N and P supply and genotype on carbon flux and partitioning in potted Pinus radiata plants. Tree Physiol. 2009, 29, 857–868. [Google Scholar] [CrossRef] [PubMed]
- Son, Y. Effects of Nitrogen Fertilization on Foliar Nutrient Dynamics in Ginkgo Seedlings. J. Plant Nutr. 2002, 25, 93–102. [Google Scholar] [CrossRef]
- Son, Y.; Kim, H.W. Above-ground biomass and nutrient distribution in a 15-year-old ginkgo (Ginkgo biloba) plantation in Central Korea. Bioresour. Technol. 1998, 63, 173–177. [Google Scholar] [CrossRef]
- Yuan, J.; Li, Q.; Xiao, G.L.; Zhu, S.H. Discussion on the utilization and development of ginkgo wood. China For. Sci. Technol. 2008, 16, 6–8. [Google Scholar]
- Andersson, S.; Wang, Y.; Ponni, R.; Hanninen, T.; Mononen, M.; Ren, H.; Serimaa, R.; Saranpää, P. Cellulose structure and lignin distribution in normal and compression wood of the Maidenhair tree (Ginkgo biloba L.). J. Integr. Plant Biol. 2015, 57, 388–395. [Google Scholar] [CrossRef] [PubMed]
- Burgert, I.; Fruhmann, K.; Keckes, J.; Fratzl, P.; Stanzl-Tschegg, S. Structure-function relationships of four compression wood types: Micromechanical properties at the tissue and fibre level. Trees 2004, 18, 480–485. [Google Scholar] [CrossRef]
- Gong, Q.L.; Hu, A.H.; Xing, S.Y.; Wang, F. Research on systematic evolution of Ginkgo biloba based on chemical composition of wood. Spectrosc. Spectr. Anal. 2009, 29, 1512–1516. [Google Scholar]
- Moilanen, M.; Hökkä, H. The growth response of scots pine to PK fertilization depends on the nutrient status of the stand on drained peatlands. Suo 2009, 60, 111–119. [Google Scholar]
- Smethurst, P.; Baillie, C.; Cherry, M.; Holz, G. Fertilizer effects on LAI and growth of four Eucalyptus nitens plantations. For. Ecol. Manag. 2003, 176, 531–542. [Google Scholar] [CrossRef]
- Grant, C.; Bittman, S.; Montreal, M.; Plenchette, C.; Morel, C. Soil and fertilizer phosphorus: Effects on plant P supply and mycorrhizal development. Can. J. Plant Sci. 2005, 85, 3–14. [Google Scholar] [CrossRef]
- Bussi, C.; Smith, M.A.L. Effects of nitrogen and potassium fertilization on the growth, yield and pitburn of apricot (cv. Bergeron). J. Hortic. Sci. Biotechnol. 1998, 73, 387–392. [Google Scholar] [CrossRef]
- Boyce, R.L.; Larson, J.R.; Sanford, R.L. Phosphorus and nitrogen limitations to photosynthesis in Rocky Mountain bristlecone pine (Pinus aristata) in Colorado. Tree Physiol. 2006, 26, 1477–1486. [Google Scholar] [CrossRef] [PubMed]
- Turnbull, T.L.; Warren, C.R.; Adams, M.A. Novel mannose-sequestration technique reveals variation in subcellular orthophosphate pools do not explain the effects of phosphorus nutrition on photosynthesis in Eucalyptus globulus seedlings. New Phytol. 2007, 176, 849–861. [Google Scholar] [CrossRef] [PubMed]
- Moilanen, M.; Silfverberg, K.; Hökkä, H.; Issakainen, J. Comparing effects of wood ash and commercial PK fertiliser on the nutrient status and stand growth of Scots pine on drained mires. Balt. For. 2004, 10, 2–10. [Google Scholar]
- Santiago, L.S.; Wright, S.J.; Harms, K.E.; Yavitt, J.B.; Korine, C.; Garcia, M.N.; Turner, B.L. Tropical tree seedling growth responses to nitrogen, phosphorus and potassium addition. J. Ecol. 2012, 100, 309–316. [Google Scholar] [CrossRef]
- Oosterhuis, D.M.; Loka, D.A.; Raper, T.B. Potassium and stress alleviation: Physiological functions and management of cotton. J. Plant Nutr. Soil Sci. 2013, 76, 331–343. [Google Scholar] [CrossRef]
- Römheld, V.; Kirkby, E.A. Research on potassium in agriculture: Needs and prospects. Plant Soil 2010, 335, 155–180. [Google Scholar] [CrossRef]
- Wang, X.G.; Zhao, X.H.; Jiang, C.J.; Li, C.H.; Cong, S.; Wu, D.; Chen, Y.Q.; Yu, H.Q.; Wang, C.Y. Effects of potassium deficiency on photosynthesis and photo-protection mechanisms in soybean (Glycine max (L.) Merr.). J. Integr. Agric. 2015, 14, 856–863. [Google Scholar] [CrossRef]
- Warren, C.R.; Adams, M.A.; Chen, Z.L. Is photosynthesis related to concentrations of nitrogen and Rubisco in leaves of Australian native plants? Funct. Plant Biol. 2000, 27, 407–416. [Google Scholar] [CrossRef]
- Pokharel, P.; Chang, S.X. Exponential fertilization promotes seedling growth by increasing nitrogen retranslocation in trembling aspen planted for oil sands reclamation. For. Ecol. Manag. 2016, 372, 35–43. [Google Scholar] [CrossRef]
- Warren, C.R.; Adams, M.A. Phosphorus affects growth and partitioning of nitrogen to Rubisco in Pinus pinaster. Tree Physiol. 2002, 22, 11–19. [Google Scholar] [CrossRef] [PubMed]
- Kumagai, E.; Araki, T.; Kubota, F. Effects of nitrogen supply restriction on gas exchange and photosystem 2 function in flag leaves of a traditional low-yield cultivar and a recently improved high-yield cultivar of rice (Oryza sativa L.). Photosynthetica 2007, 45, 489–495. [Google Scholar] [CrossRef]
- Zafar, M.; Abbasi, M.K.; Khaliq, A. Effect of combining organic materials with inorganic phosphorus sources on growth, yield, energy content and phosphorus uptake in maize at Rawalakot Azad Jammu and Kashmir, Pakistan. Arch. Appl. Sci. Res. 2011, 3, 199–212. [Google Scholar]
- Zorb, C.; Senbayram, M.; Peiter, E. Potassium in agriculture: Status and perspectives. J. Plant Physiol. 2014, 171, 656–669. [Google Scholar] [CrossRef] [PubMed]
- Terry, N.; Ulrich, A. Effects of phosphorus deficiency on the photosynthesis and respiration of leaves of sugar beet. Plant Physiol. 1973, 51, 43–47. [Google Scholar] [CrossRef] [PubMed]
- Warren, C.R.; McGrath, J.F.; Adams, M.A. Differential effects of N, P and K on photosynthesis and partitioning of N in Pinus pinaster needles. Ann. For. Sci. 2005, 62, 1–8. [Google Scholar] [CrossRef]
- Moilanen, M.; Silfverberg, K.; Hokkanen, T.J. Effects of wood-ash on the tree growth, vegetation and substrate quality of a drained mire: A case study. For. Ecol. Manag. 2002, 171, 321–338. [Google Scholar] [CrossRef]
- Silfverberg, K.; Issakainen, J.; Moilanen, M. Growth and nutrition of scots pine on drained and fertilized purple moor grass fens in central Finland. Balt. For. 2011, 17, 91–101. [Google Scholar]
- Whitehead, D.; Beadle, C.L. Physiological regulation of productivity and water use in Eucalyptus: A review. For. Ecol. Manag. 2004, 193, 113–140. [Google Scholar] [CrossRef]
- Dickmann, D.I.; Nguyen, P.V.; Pregitzer, K.S. Effects of irrigation and coppicing on above-ground growth, physiology, and fine-root dynamics of two field-grown hybrid poplar clones. For. Ecol. Manag. 1996, 80, 163–174. [Google Scholar] [CrossRef]
- Thornton, F.C.; Bock, B.R.; Behel, A.D.; Houston, A.; Tyler, D.D. Utilization of waste materials to promote hardwood tree growth. South. J. Appl. For. 2000, 24, 230–237. [Google Scholar]
- Barrongafford, G.A.; Will, R.E.; Burkes, E.C.; Shiver, B.; Teskey, R.O. Nutrient concentrations and contents, and their relation to stem growth, of intensively managed Pinus taeda and Pinus elliottii stands of different planting densities. For. Sci. 2003, 49, 291–300. [Google Scholar]
- Tausz, M.; Trummer, W.; Wonisch, A.; Goessler, W.; Grill, D.; Jiménez, M.S.; Morales, D. A survey of foliar mineral nutrient concentrations of Pinus canariensis at field plots in Tenerife. For. Ecol. Manag. 2004, 189, 49–55. [Google Scholar] [CrossRef]
- Jennifer, A.B.; Boerner, R.E.J. Nitrogen fertilization effects on foliar nutrient dynamics and autumnal resorption in maidenhair tree (Gingko biloba L.). J. Plant Nutr. 1994, 17, 433–443. [Google Scholar]
- Pettigrew, W.T.; Meredith, W.R. Dry matter production, nutrient uptake, and growth of cotton as affected by potassium fertilization. J. Plant Nutr. 1997, 20, 531–548. [Google Scholar] [CrossRef]
- Wright, I.J.; Reich, P.B.; Westoby, M.; Ackerly, D.D.; Baruch, Z.; Bongers, F.; Cavender-Bares, J.; Chapin, T.; Cornelissen, J.H.C.; Diemer, M. The worldwide leaf economics spectrum. Nature 2004, 428, 821–827. [Google Scholar] [CrossRef] [PubMed]
- Sundström, E.; Magnusson, T.; Hånell, B. Nutrient conditions in drained peatlands along a north-south climatic gradient in Sweden. For. Ecol. Manag. 2000, 126, 149–161. [Google Scholar] [CrossRef]
- Pietiläinen, P.; Kaunisto, S. The effect of peat nitrogen concentration and fertilization on the foliar nitrogen concentration of Scots pine (Pinus sylvestris L.) in three temperature sum regions. Suo 2003, 54, 1–13. [Google Scholar]
- Wang, D.G.; Yin, G.T.; Yang, J.C.; Li, R.S.; Zou, W.T.; Jia, R.F. Effects of phosphorus fertilization on growth and foliar nutrient (N, P, K) of Phoebe bournei seedlings. J. Nanjing For. Univ. (Nat. Sci. Ed.) 2014, 38, 40–44. [Google Scholar]
- Crous, J.W.; Morris, A.R.; Scholes, M.C. Growth and foliar nutrient response to recent applications of phosphorus (P) and potassium (K) and to residual P and K fertiliser applied to the previous rotation of Pinus patula, at Usutu, Swaziland. For. Ecol. Manag. 2008, 256, 712–721. [Google Scholar] [CrossRef]
- Nakashgir, G.H. Influence of potassium on nitrogen utilization by maize under dryland conditions as affected by water storage. Adv. Plant Sci. 1992, 5, 134–142. [Google Scholar]
- Nardini, A.; Grego, F.; Trifilò, P.; Salleo, S. Changes of xylem sap ionic content and stem hydraulics in response to irradiance in Laurus nobilis. Tree Physiol. 2010, 30, 136–140. [Google Scholar] [CrossRef] [PubMed]
- Sellin, A. Experimental evidence supporting the concept of light-mediated modulation of stem hydraulic conductance. Tree Physiol. 2010, 30, 1528–1535. [Google Scholar] [CrossRef] [PubMed]
- Pervez, H.; Ashraf, M.; Makhdum, M.I. Influence of potassium nutrition on gas exchange characteristics and water relations in cotton (Gossypium hirsutum L.). Photosynthetica 2004, 42, 251–255. [Google Scholar] [CrossRef]





{kind=link}
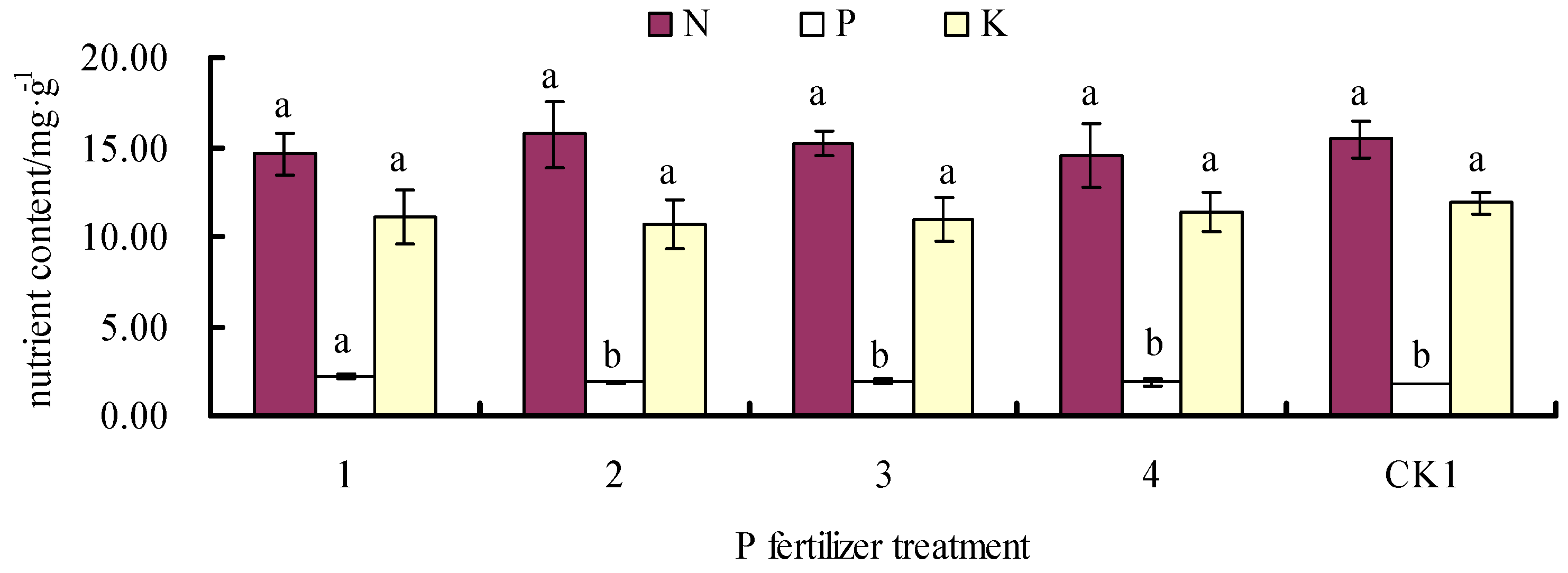{kind=link}
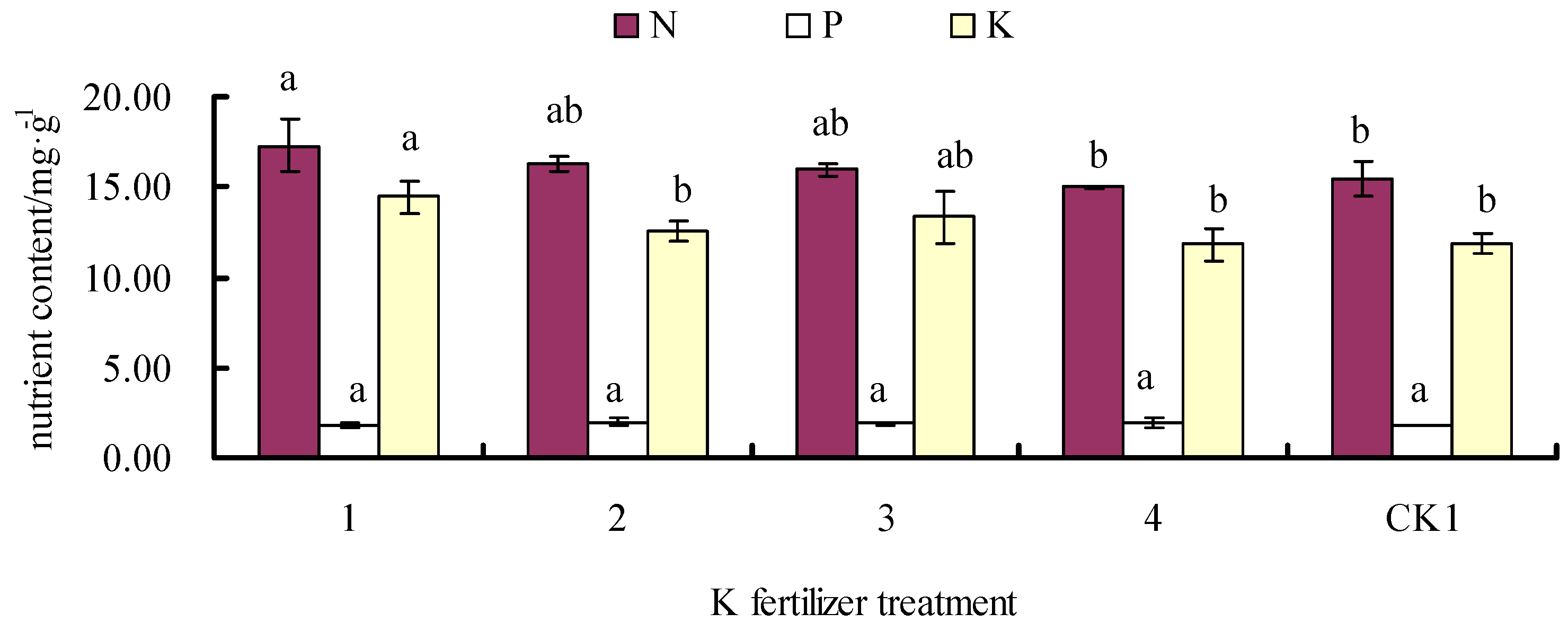{kind=link}
{kind=link}
| Treatment No. | Fertilization Level (g·tree−1) | ||
|---|---|---|---|
| Urea | Superphosphate | Potassium Sulfate | |
| 1 | 100 | 200 | 15 |
| 2 | 100 | 400 | 40 |
| 3 | 100 | 600 | 90 |
| 4 | 200 | 200 | 40 |
| 5 | 200 | 400 | 90 |
| 6 | 200 | 600 | 15 |
| 7 | 400 | 200 | 90 |
| 8 | 400 | 400 | 15 |
| 9 | 400 | 600 | 40 |
| CK2 | 0 | 0 | 0 |
| F | DBH | H | NSL | V |
|---|---|---|---|---|
| N1 600 | 15.34 ± 0.396 a | 9.85 ± 0.383 a | 26.59 ± 1.204 a | 0.0146 ± 0.000929 a |
| N2 400 | 15.11 ± 0.123 a | 9.27 ± 0.492 b | 25.56 ± 1.284 ab | 0.0135 ± 0.000755 ab |
| N3 200 | 14.96 ± 0.301 a | 9.17 ± 0.131 b | 24.64 ± 2.840 ab | 0.0133 ± 0.000448 b |
| N4 100 | 14.92 ± 0.407 a | 9.14 ± 0.068 b | 21.93 ± 2.635 bc | 0.0132 ± 0.000369 b |
| CK1 0 | 14.35 ± 0.095 b | 8.92 ± 0.158 b | 20.23 ± 2.594 c | 0.0124 ± 0.000256 b |
| P1 800 | 14.84 ± 0.063 a | 9.45 ± 0.257 a | 25.98 ± 1.33 a | 0.0136 ± 0.000398 a |
| P2 600 | 14.42 ± 0.255 ab | 9.13 ± 0.221 ab | 24.02 ± 1.86 ab | 0.0127 ± 0.000408 b |
| P3 400 | 14.23 ± 0.333 b | 9.05 ± 0.182 ab | 24.37 ± 0.688 ab | 0.0124 ± 0.000514 b |
| P4 200 | 14.21 ± 0.292 b | 8.83 ± 0.270 b | 22.69 ± 0.419 bc | 0.0121 ± 0.000464 b |
| CK1 0 | 14.35 ± 0.095 b | 8.92 ± 0.158 b | 20.23 ± 2.594 c | 0.0124 ± 0.000256 b |
| K1 200 | 14.91 ± 0.499 a | 9.01 ± 0.288 a | 24.01 ± 0.844 a | 0.0124 ± 0.000965 a |
| K2 90 | 14.65 ± 0.428 a | 8.93 ± 0.195 a | 23.71 ± 1.29 a | 0.0117 ± 0.000445 a |
| K3 40 | 14.66 ± 0.553 a | 8.88 ± 0.326 a | 23.35 ± 2.14 a | 0.0113 ± 0.000699 a |
| K4 15 | 14.38 ± 0.422 a | 8.63 ± 0.459 a | 24.49 ± 0.967 a | 0.0114 ± 0.000993 a |
| CK1 0 | 14.35 ± 0.095 a | 8.92 ± 0.158 a | 20.23 ± 2.59 a | 0.0124 ± 0.000256 a |
| F | DBH | H | NSL | V |
|---|---|---|---|---|
| 1 | 13.66 ± 0.908 bcd | 7.45 ± 0.022 bc | 24.15 ± 0.832 bcd | 0.00983 ± 0.000675 cd |
| 2 | 13.78 ± 0.825 abcd | 7.49 ± 0.065 bc | 26.606 ± 3.250 abc | 0.00997 ± 0.000679 cd |
| 3 | 13.2 ± 0.172 cd | 7.42 ± 0.168 bc | 26.00 ± 2.32 abc | 0.00947 ± 0.000327 de |
| 4 | 13.71 ± 0.233 bcd | 7.65 ± 0.123 ab | 25.90 ± 1.921 abc | 0.01013 ± 0.000207 cd |
| 5 | 13.88 ± 0.173 abc | 7.73 ± 0.293 ab | 25.26 ± 1.705 abc | 0.01036 ± 0.000261 bc |
| 6 | 13.94 ± 0.370 abc | 7.55 ± 0.046 b | 22.83 ± 0.656 cd | 0.01017 ± 0.000304 cd |
| 7 | 14.61 ± 0.347 a | 8.01 ± 0.239 a | 26.77 ± 4.070 abc | 0.01130 ± 0.000302 a |
| 8 | 14.50 ± 0.423 ab | 7.81 ± 0.173 ab | 28.89 ± 2.014 a | 0.01094 ± 0.000240 ab |
| 9 | 14.47 ± 0.208 ab | 7.81 ± 0.205 ab | 27.49 ± 1.980 ab | 0.01092 ± 0.000304 ab |
| CK2 | 12.92 ± 0.231 d | 7.09 ± 0.0346 c | 20.23 ± 2.594 d | 0.00885 ± 0.000161 e |
| F | Ci | Tr | Gs | Pn |
|---|---|---|---|---|
| N1 600 | 278.56 ± 8.57 a | 2.71 ± 0.252 a | 167.64 ± 3.32 a | 12.30 ± 0.472 a |
| N2 400 | 277.33 ± 4.98 a | 2.68 ± 0.150 a | 165.78 ± 2.55 a | 11.74 ± 0.597 ab |
| N3 200 | 269.78 ± 5.10 a | 2.43 ± 0.173 a | 157.78 ± 4.43 a | 10.25 ± 1.42 b |
| N4 100 | 267.79 ± 16.65 a | 2.51 ± 0.284 a | 156.67 ± 7.86 a | 10.57 ± 0.635 b |
| CK1 0 | 267.33 ± 21.73 a | 2.53 ± 0.208 a | 127.33 ± 15.5 b | 8.23 ± 0.833 c |
| P1 800 | 289.89 ± 3.29 a | 3.03 ± 0.153 a | 156.56 ± 5.34 a | 10.50 ± 1.19 a |
| P2 600 | 275.11 ± 8.26 a | 2.80 ± 0.152 a | 139.78 ± 4.17 b | 10.22 ± 1.06 ab |
| P3 400 | 266.89 ± 5.23 a | 2.62 ± 0. 135 a | 129.89 ± 8.06 b | 8.77 ± 1.72 abc |
| P4 200 | 275.33 ± 19.50 a | 2.67 ± 0.333 a | 133.88 ± 6.02 b | 7.14 ± 0.635 c |
| CK1 0 | 267.33 ± 21.73 a | 2.53 ± 0.208 a | 127.33 ± 15.50 b | 8.23 ± 0.833 bc |
| K1 200 | 256.89 ± 1.92 a | 2.60 ± 0.101 a | 148.22 ± 11.5 a | 10.39 ± 0.27 a |
| K2 90 | 251.00 ± 4.58 a | 2.54 ± 0.081 a | 133.11 ± 10.98 a | 10.44 ± 0.44 a |
| K3 40 | 265.78 ± 7.93 a | 2.46 ± 0.102 a | 123.67 ± 15.94 a | 8.37 ± 0.62 b |
| K4 15 | 266.79 ± 9.25 a | 2.27 ± 0.095 a | 121.56 ± 16.60 a | 7.61 ± 0.37 b |
| CK1 0 | 267.33 ± 21.73 a | 2.53 ± 0.208 a | 127.33 ± 15.50 a | 8.23 ± 0.83 b |
| F | Ci | Tr | Gs | Pn |
|---|---|---|---|---|
| 1 | 239.00 ± 10.40 b | 2.56 ± 0.34 a | 122.33 ± 13.39 c | 9.00 ± 0.91 cd |
| 2 | 246.78 ± 27.03 b | 2.41 ± 0.20 a | 127.33 ± 13.00 bc | 9.22 ± 0.74 bcd |
| 3 | 243.00 ± 25.32 b | 2.41 ± 0.47 a | 136.11 ± 7.31 abc | 9.36 ± 1.02 bcd |
| 4 | 254.00 ± 8.09 b | 2.61 ± 0.24 a | 133.56 ± 9.52 bc | 9.48 ± 0.17 bcd |
| 5 | 247.33 ± 5.20 b | 2.59 ± 0.34 a | 141.34 ± 3.51 abc | 9.71 ± 1.51 abcd |
| 6 | 272.11 ± 17.88 ab | 2.74 ± 0.15 a | 140.11 ± 4.40 abc | 10.11 ± 0.23 abc |
| 7 | 271.78 ± 16.68 ab | 2.69 ± 0.25 a | 142.67 ± 3.51 abc | 10.68 ± 1.22 abc |
| 8 | 272.55 ± 20.51 ab | 2.69 ± 0.08 a | 146.89 ± 12.98 ab | 10.71 ± 0.71 ab |
| 9 | 290.33 ± 16.18 a | 2.79 ± 0.16 a | 154.33 ± 12.20 a | 11.27 ± 0.38 a |
| CK2 | 265.33 ± 18.73 ab | 2.50 ± 0.11 a | 129.13 ± 12.50 bc | 8.53 ± 0.58 d |
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Guo, J.; Wu, Y.; Wang, B.; Lu, Y.; Cao, F.; Wang, G. The Effects of Fertilization on the Growth and Physiological Characteristics of Ginkgo biloba L. Forests 2016, 7, 293. https://doi.org/10.3390/f7120293
Guo J, Wu Y, Wang B, Lu Y, Cao F, Wang G. The Effects of Fertilization on the Growth and Physiological Characteristics of Ginkgo biloba L. Forests. 2016; 7(12):293. https://doi.org/10.3390/f7120293
Chicago/Turabian StyleGuo, Jing, Yaqiong Wu, Bo Wang, Yan Lu, Fuliang Cao, and Guibin Wang. 2016. "The Effects of Fertilization on the Growth and Physiological Characteristics of Ginkgo biloba L." Forests 7, no. 12: 293. https://doi.org/10.3390/f7120293